
湖南低山丘陵区薄壳山核桃的开花物候期观测及品种配置
2021-02-02 06:30魏海林高昌虎王大故冯楠可刘榕榕吕芳德
南京林业大学学报(自然科学版) 2021年1期
蒋 瑶,魏海林,高昌虎,王大故,冯楠可,李 睿,刘榕榕,吕芳德
(1.中南林业科技大学,经济林培育与保护省部共建教育部重点实验室,经济林育种与栽培国家林业局重点实验室,经济林培育与利用湖南省协同创新中心,湖南 长沙 410004;2.湖南省林业科学院,湖南 长沙 410004;3.怀化市鹤城区林业局,湖南 怀化 418000;4.怀化市中方县农业局, 湖南 怀化 418005)
薄壳山核桃[Caryaillinoinensis(Wangenh.)K.Koch]为胡桃科(Juglandaceae)山核桃属(Carya)多年生落叶乔木,其树体高大挺拔、坚果营养丰富且油脂健康,为优良的果材油兼用树种[1-2]。随着人们对薄壳山核桃需求量日益增加,其产业发展前景良好。薄壳山核桃是雌雄异花同株的树种,雌雄成熟时间有差异,雄花散粉期与雌花可授期完全一致的品种不多[3],因此授粉不良造成落果、低产是影响薄壳山核桃产业发展的一个重要问题。由于各地的气候条件差异,引种的薄壳山核桃开花物候期与原产地不一致,完全套用原产地或其他地区的品种配置往往并不合理。当前,充分了解薄壳山核桃各品种的开花物候期,配置适宜的授粉树是国内外研究热点[4-7]。因此,观察当地薄壳山核桃生物学特性变化、正确认识与掌握薄壳山核桃花期是制定适宜栽培技术及措施的重要依据。特别是掌握薄壳山核桃生长开花的物候特征,是合理搭配品种、提高薄壳山核桃产量的理论基础之一。本研究对湖南省怀化市14个薄壳山核桃品种的花发育情况及花期特性进行持续观察,分析并掌握雌雄花开花物候期,以期为解决薄壳山核桃雌雄花期不遇问题、选择配置各品种的授粉树并提高坐果率提供参考。
1 材料与方法
1.1 试验地概况
试验地设在湖南省怀化市中方县桐木镇薄壳山核桃基地(109°85′E,27°32′N),海拔397 m,属湖南低山丘陵区,沿沉水两岸有河溪冲积小平原。地带性土壤为红壤,建园区内主要是红壤、山地黄壤,有机质含量为25~30 mg/g,pH 5.3左右,土层较为深厚肥沃、保水力强。该地区属亚热带季风气候,四季分明,日照充足,雨量充沛,年降雨量1 378 mm,年均气温17 ℃,1月均温5.3 ℃,7月均温27.9 ℃,年日照总时间1 336.9 h,年绝对无霜期为297 d。
1.2 试验材料
选取14个薄壳山核桃品种作为花期观测材料。试验材料均为长势良好且一致、发育正常、树冠开阔、无明显病虫害的7年生品种。供试的14个薄壳山核桃品种包括‘湘林1号’(‘XiangLin No.1’)、‘福克特’(‘Forket’)、‘密西西比’(‘Mississipi’)、‘金奥瓦’(‘Kiowa’)、‘亚林100’(‘YaLin No.100’)、‘威奇塔’(‘Wichita’)、‘佩鲁奎’(‘Peruque’)、‘波尼’(‘Pawnee’)、‘亚林13’(‘YaLin No.13’)、‘普兹内’(‘Pyzner’)、‘斯图尔特’(‘Stuart’)、‘特贾斯’(‘Tejas’)、‘马汉’(‘Mahan’)和‘卡多’(‘Caddo’)。
1.3 雌雄花的发育过程及物候期观测
在供试品种中各选取3个标准株,对雌花和雄花的不同发育阶段进行标记,观察开花过程中雌雄花外部的形态特征,并记录和拍照。
1)雌雄花物候期观察:于2020年4月在怀化市中方县桐木镇薄壳山核桃栽培示范基地进行。从供试品种中各选定2~3个标准株,在每个标准株树冠中下部外围的4个不同方位选择2~3枝生长良好且一致的枝条作为标准枝。对标准株、标准枝进行挂牌、编号,定点观察,参照核桃的物候期观察项目及标准[8]进行,前期3~4 d观测1次,后期1~2 d 1次。观察记录薄壳山核桃花发育情况。雌花初花期定为5%的花显露出子房;盛花始期为25%的雌花柱头开裂且分泌物增多,盛花期为50%的雌花柱头开裂并成为“倒八字”形且分泌物增多,盛花末期为75%的雌花柱头开裂为“倒八字”形且分泌物增多;终花期为80%的雌花柱头萎蔫。雄花萌动期以5%的雄花萌出为准,雄花序伸长期为75%的雄花序伸长至最终长度;盛花始期为25%的雄花开始散粉,盛花期为50%的雄花散粉,盛花末期为75%的雄花散粉;终花期为90%的雄花序脱落。
2)雌雄花单花物候期观测:在各方位的每个标准枝上分别选择2~4个雄花序和雌花序,标记并编号,在花开放期间每天观察1次标准枝上花的开放情况。分别观测雌花单花现蕾期、柱头开裂期、柱头“倒八字”形湿润期、柱头干涸期以及子房膨大期的时间及形态特征;观测雄花单花苞片开裂期、花药变黄期、散粉期、花药变黑以及雄花序脱落期等时期的时间及形态特征[9-10]。
3)花期气候调查:记录并统计薄壳山核桃雌雄花花芽从萌动到花期结束时的天气状况,将平均温度、降水、相对湿度变化及日照时间情况结合中国气象数据网(http://data.cma.cn/)资料,分析积温对开花物候期的指示作用。
1.4 授粉品种配置方法
在各方位的每个标准枝上分别选择3~5个雄花序和雌花序,根据标准枝上雌花柱头的发育状态判断可授期,当雌花二裂柱头打开呈“倒八字”形且湿润时视为可授。雌花可授期从一个方位标准枝上雌花均呈现“倒八字”形柱头且分泌物增多时开始,直至所有方位的标准枝雌花序的柱头表面枯萎,柱头反向卷曲;雄花散粉期从一个方位所有标准枝雄花序散粉开始,直至所有方位的标准枝均没有花粉散出为止。根据不同品种的雌花可授期和雄花散粉期进行品种配置。
1.5 数据处理
对所观测的数据进行汇总,运用Excel程序进行数据统计处理,并制图分析。
2 结果与分析
2.1 薄壳山核桃花发育外部形态特征
2.1.1 雄花形态
薄壳山核桃雄花为柔荑花序,雄花发育时可依次出现雄花序萌发、雄花序伸长、苞片由包裹紧密变成松散、花轴显露、花药逐渐饱满、花药颜色由绿变黄、散粉以及花药变黑的性状。供试薄壳山核桃各品种的雄花序生长发育进度不一致,外部形态也有较明显的区别。
雄花发育中比较明显的变化体现在雄花的长度、粗度、苞片的包裹程度、花药及花轴可见度,以及花药的颜色。以‘福克特’为代表的细长型雄花序,根据变化可分为以下6个阶段:①萌发期,雄花序于小枝基部的叶腋萌出,花序较短(3~5 cm),小花及苞片包裹紧密,直径约0.2 cm,与花轴之间的夹角约5°,此时花药和花轴均不可见(图1A),这一阶段持续4~6 d;②苞片松散始期,小花开始分离,苞片也向外展开,雄花序伸长至5~7 cm,直径约0.4 cm,苞片逐渐打开,与花轴夹角为15°,花药和花轴仍不可见(图1B),此阶段持续3~4 d;③雄花序生长期,雄花序伸长至6~9 cm,直径约0.6 cm,苞片与花轴夹角为45°,花药和花轴不可见(图1C),此阶段持续3~6 d;④花药可见期,雄花序伸长至7~12 cm,直径约0.8 cm,苞片与花轴夹角为75°,花药可见并逐渐饱满,花轴不明显(图1D),此阶段持续3~4 d;⑤花药变黄期,雄花序伸长至8~13 cm,直径约0.9 cm,苞片与花轴夹角为90°,花药饱满呈黄色,即将散粉,花轴明显可见(图1E),此阶段持续3~6 d;⑥花药变黑期,小花从基部开始散粉,并向先端延伸,散粉后1~3 d花药变成黑色,并逐渐脱落(图1F)。具细长型雄花序的品种有‘福克特’、‘金奥瓦’、‘亚林100’、‘威奇塔’、‘亚林13’、‘特贾斯’和‘马汉’。

A-F.‘福克特’雄花发育的萌发期、苞片松散始期、雄花序生长期、花药可见期、花药变黄期及花药变黑期germination period, perianth dehiscence period, elongation period, anther visible period, anther turning yellow period, anther turning black period of ‘Forket’;G-L.‘波尼’雄花发育的萌发期、苞片松散始期、雄花序生长期、花药饱满期、花药变黄期及花药变黑期germination period, perianth dehiscence period, elongation period, anther fullness period, anther turning yellow period, anther turning black period of ‘Pawnee’。图1 薄壳山核桃雄花发育形态Fig.1 Developmental morphology of male flowers of Carya illinoinensis
以‘波尼’为代表的粗短型雄花序发育过程与‘福克特’类似,区别在于粗短型雄花序在萌发期其苞片与花轴的夹角即达10°左右,花序直径约0.3 cm,较粗(图1G);在第2阶段雄花序上的小花已分离开,此时可见绿色花药,苞片与花轴夹角约为40°(图1H);第3至第5阶段与细长型雄花区别在于花轴并不明显,小花之间距离较短,雄花序直径0.9~1.2 cm,长度为6~10 cm,从外形上显得更加粗短(图1I-1K)。具粗短型雄花序的品种有:‘波尼’、‘湘林1号’、‘普兹内’、‘佩鲁奎’、‘斯图尔特’和‘卡多’。‘密西西比’介于细长型和粗短型之间,属于中长型。
2.1.2 雌花形态

A-F.‘福克特’雌花发育的现蕾期、柱头开裂始期、柱头V字型期、柱头倒八字型期、柱头干涸期及柱头变黑期 budding period, stigma cracking period, stigma V-shaped period, stigma inverted-shaped period, stigma wilting period and stigma turning black period of ‘Forket’ ;G.‘特贾斯’雌花female flower of ‘Tejas’; H-M.‘波尼’雄花发育的现蕾期、柱头圆凸状期、柱头平展期、柱头圆锥状期、柱头干涸期及柱头变黑期 budding period, stigma round convex period, stigma flattening period, stigma conical period, stigma wilting period and stigma turning black period of ‘Pawnee’.图2 薄壳山核桃雄花发育形态Fig.2 Developmental morphology of female flowers of Carya illinoinensis
薄壳山核桃不同品种雌花呈现的外部形态相差较大,主要表现为雌花的颜色和柱头开裂情况。
以‘福克特’为代表的品种其雌蕊呈黄绿色,发育过程可分为:①现蕾期,4月下旬新梢展开8~10片复叶时,雌蕊萌发于枝条顶端,此时子房刚露出,呈圆柱状,外观俯看似绿色圆点,二裂柱头全合拢,无受精活力,此阶段3~5 d(图2G);②柱头开裂始期,柱头裂开,向两边略张,基部肉质类蟹钳状,二裂柱头间夹角15°~30°,持续3~4 d(图2H);③V字型期,二裂柱头呈“V字”形,柱头角度加大至4柱头上有少量突触状腺质细胞,持续4~8 d(图2I);④倒八字型期,二裂柱头呈“倒八字”形,柱头间夹角约120°,柱头上有大量突触状腺质细胞,分泌物增多,黏性增加,柱头由绿色变成黄绿色,雌蕊内部胚珠发育完成,珠被、珠心及珠柄分化基本完成[11],此时为最佳授粉期,持续 4~7 d(图2J);⑤柱头干涸期,柱头稍合拢,表面突出物萎蔫干涸,并伴有少量黑斑,表明进入授粉末期,持续4~7 d(图2K)此时授粉效果极差;⑥柱头变黑期,柱头间夹角5°~90°,完全变黑,此雌花不具有接受花粉的能力,子房膨大,此时为雌花终花期(图2L)。从雌花子房显露到子房开始膨大需要18~31 d。观察还发现,‘特贾斯’等品种的柱头开裂角度并不明显,在达到可授期时其柱头呈现类圆锥状,不经历V字型期和倒八字型期,外形与‘波尼’类似(图2M)。雌花柱头呈绿色的品种有:‘福克特’、‘密西西比’、‘威奇塔’、‘佩鲁奎’、‘亚林13’、‘斯图尔特’、‘特贾斯’和‘马汉’。
以‘波尼’为代表的品种其雌蕊呈红棕色,在柱头的颜色及二裂柱头开裂的角度和形态方面与绿色型柱头存在明显区别。此类品种在萌发时呈绿色(图2H),待柱头明显可见时即显露出红棕色,此时柱头分裂情况并不明显,整体呈圆凸状,突起物上有3条明显的缢痕(图2I),并不具备接受花粉的能力;进入相应的V字型期时其柱头基部愈合部分较多,柱头不明显,分裂角度近180°(图2J);在可授期时柱头基部进一步膨大,整体似圆锥状,颜色加深,此时柱头表面突触状腺质细胞明显增加,并且伴有大量分泌物,黏性强,此时授粉效果最好(图2K);3~4 d后柱头颜色变深,呈现出紫红色,表面干涸,进入授粉末期,此时授粉效果较差,为雌花盛花末期(图2L);此后4~6 d柱头完全变黑,此时雌花不具有接受花粉的能力,子房膨大(图2M)。雌花柱头呈红色的品种有:‘波尼’、‘湘林1号’、‘金奥瓦’、‘亚林100’、‘普兹内’和‘卡多’。‘密西西比’雌花柱头为‘淡红色’。
2.2 薄壳山核桃雌雄花的开花物候期
2.2.1 雄花开花过程
薄壳山核桃的雄花为葇荑花序,如雄花开花物候期(表1)所示,14个品种雄花花序的萌动期从3月25日持续到4月18日。其中‘普兹内’的雄花在3月25日萌动,时间最早;萌动较晚的是‘福克特’、‘亚林100号’、‘亚林13号’和‘斯图尔特’,其中‘福克特’在4月23日才达到5%雄花序萌动,14个品种在萌动时间上相差28 d。雄花序从萌动后即进入花序伸长期,经历13~26 d,各品种75%的雄花序均伸至最大长度,雄花序最早伸至最长的品种是‘普兹内’、最晚的是‘福克特’,但‘普兹内’雄花序伸长的持续时间长达26 d。各品种雄花序的散粉期(包括始期、盛期和末期)集中在4月24日至5月21日,持续6~12 d。其中散粉最早的是‘普兹内’,散粉最晚的是‘福克特’;散粉时间持续最长的是‘特贾斯’(13 d),散粉时间持续最短的是‘金奥瓦’、‘佩鲁奎’和‘卡多’,均为6 d。试验材料中雄花序的终花期集中在5月,‘普兹内’落花最早,与其花芽萌发早是相对应的;落花最晚的是‘福克特’,该品种具有持续萌发雄花序的特点,可在同一时期出现萌发和散粉的雄花序,整个雄花序发育的时间相对较长,且该品种雄花序萌发量大、花粉量多,十分适宜作为授粉树。
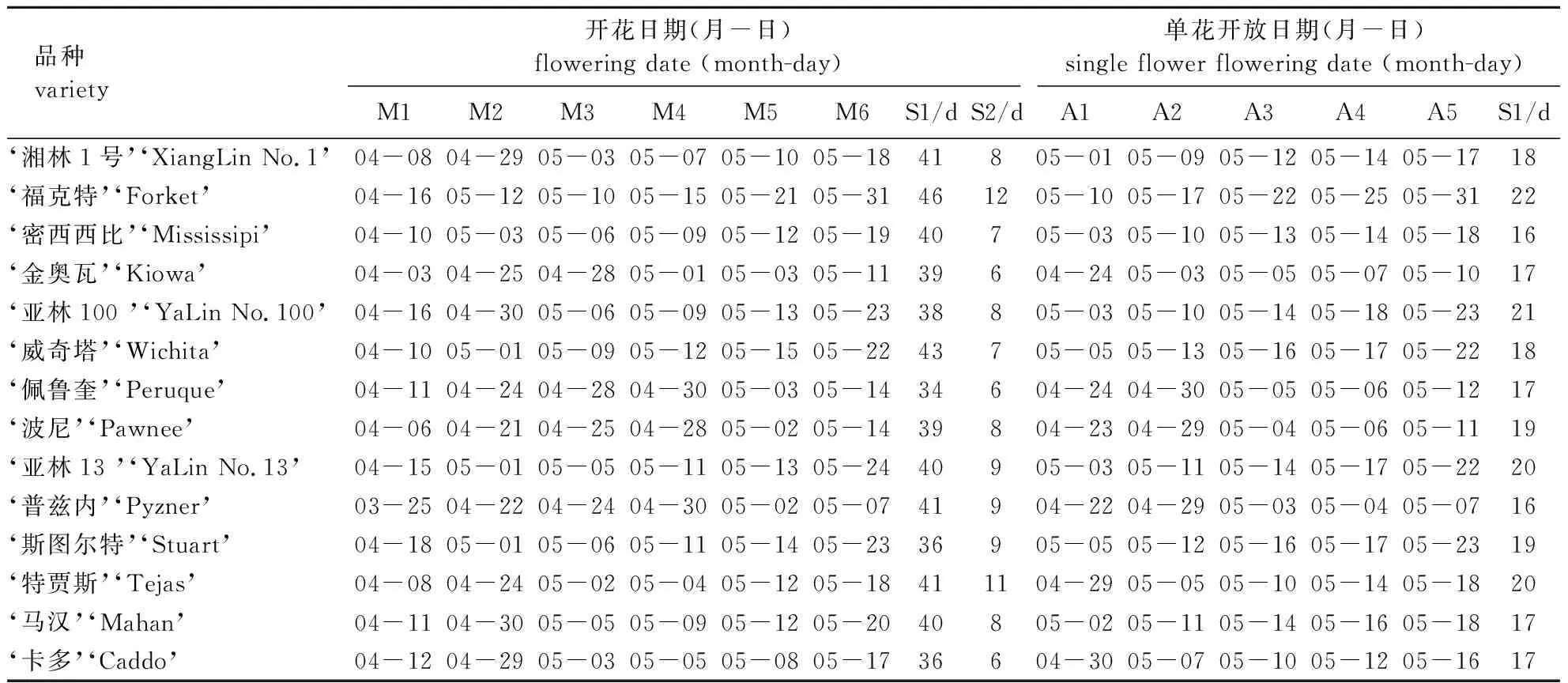
表1 薄壳山核桃不同品种雄花的开花物候期及单花开花过程
薄壳山核桃不同品种雄花的单花开放过程详见表1。由表1可知:薄壳山核桃的14个品种苞片开裂期集中在4月22日至5月10日,各品种之间相差19 d,苞片裂开时间最早的是‘普兹内’,最晚的是‘福克特’。各品种雄花花药由绿变黄的时间从4月29持续至5月17日,最早进入花药变黄期的是‘普兹内’和‘波尼’,最晚的是‘福克特’,其中从苞片开裂进入花药变黄期所需时间最长的是‘福克特’(8 d),最短的是‘威奇塔’(4 d)。雄花药变黄之后各品种均在3~6 d之内进入花药散粉期,其中进入花药散粉期最早的是‘普兹内’,其次是‘波尼’、‘佩鲁奎’和‘金奥瓦’,最晚的是‘福克特’。雄花药散粉之后的1~5 d花药变黑,各品种花药变黑时间从 5月4日持续到5月25日,其中进入花药变黑期最早的是‘普兹内’,最晚的是‘福克特’。最后雄花序陆续脱落,各品种脱落时间从5月7日持续到5月31日,最早的是‘普兹内’,最晚的是‘福克特’。试验材料中雄花单花开放持续时间为16~22 d,其中‘福克特’雄花单花开放持续时间最长,达到22 d,而‘普兹内’和‘密西西比’雄花单花开花时间只持续了16 d。
2.2.2 雌花开花过程
薄壳山核桃的雌花为穗状花序,一般由4~7朵雌花聚成伞状生长在新梢的顶端,每朵雌花由总苞、花被和子房组成。对雌花开花物候观测(表2)可知,所观测品种的雌花均在4月中下旬露出子房,最早的是‘特贾斯’,4月中旬即现蕾,其次是‘斯图尔特’和‘普兹内’,它们的雌花最早达到可授期,调查显示这3个品种也是果实早熟的品种。其余品种均在4月下旬现蕾,最晚的品种是‘威奇塔’,直至4月底才现蕾。不同品种的雌花发育期并不一致,一般的品种现蕾期持续3~6 d,但‘湘林1号’和‘福克特’同一单株上现蕾时间可达10~12 d。因此,在同一株树上可观察到有的雌花刚刚现蕾,有的已进入可授期。这种持续现蕾的特性能使雌花接受不同时期的花粉,有利于提高坐果率。薄壳山核桃各品种在现蕾后8~15 d进入盛花始期,从4月28日持续到5月8日,最早进入盛花始期的是‘特贾斯’,最晚进入的是‘威奇塔’。各品种的雌花盛花期持续时间为6~18 d,其中雌花盛花期持续时间最长的是‘湘林1号’,其次为‘福克特’(12 d),两个品种均有持续萌发雌花序的特性;雌花盛花期持续时间最短的是‘亚林13号’和‘马汉’,其他品种的盛花期持续时间均在8~9 d。盛花期之后雌花柱头干涸萎蔫,不具有可授性。各品种进入终花期的时间集中在5月10—23日,最早进入终花期的是‘特贾斯’,最晚进入的是‘福克特’。两者相差14 d。各品种之间的雌花从现蕾到柱头萎蔫不可授粉的持续时间为20~32 d(表2)。
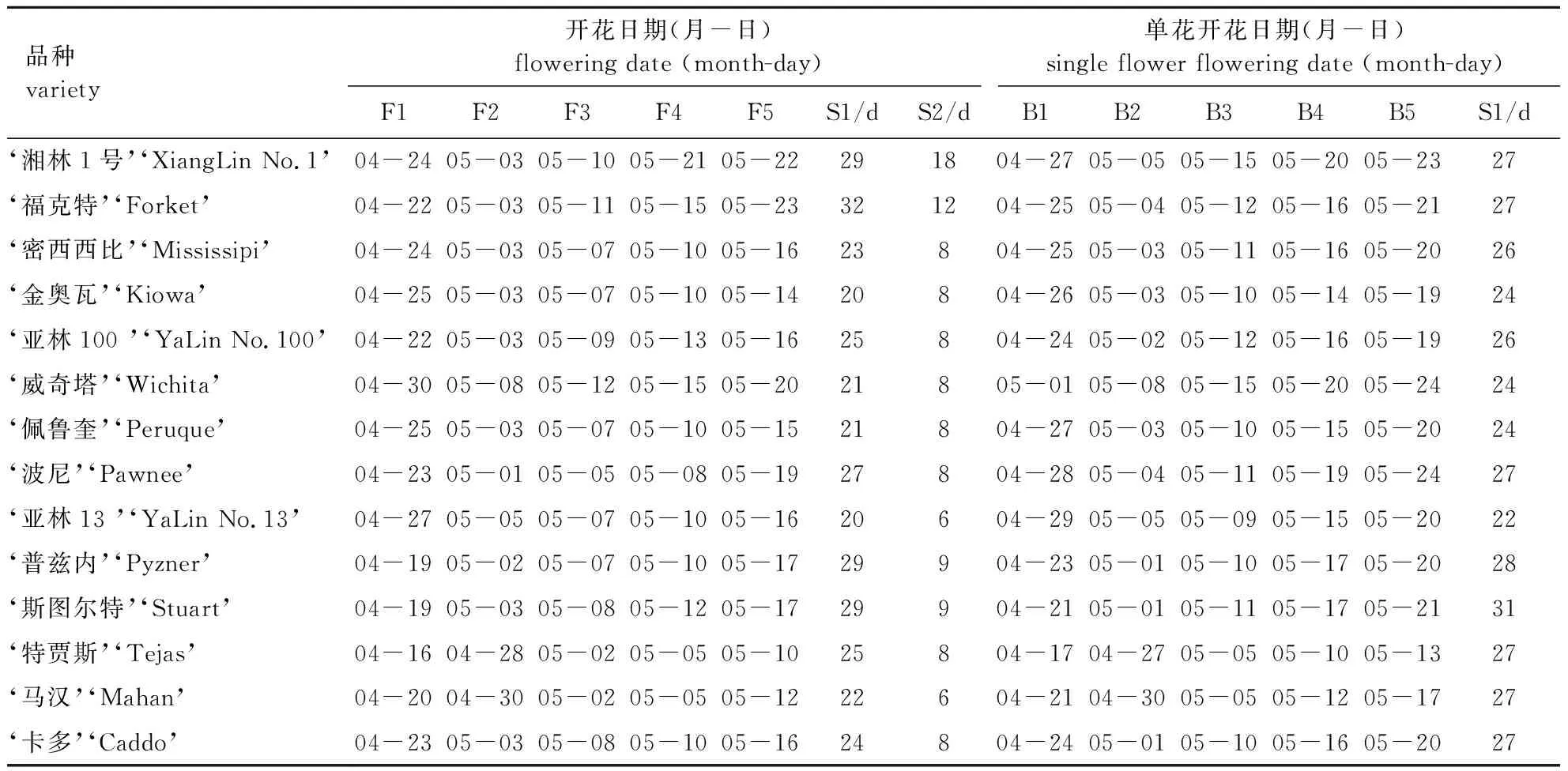
表2 薄壳山核桃不同品种雌花开花物候期及单花开放过程
薄壳山核桃14个品种的雌花单花开放观察结果见表2。由表2可知:试验材料中现蕾期集中在4月17日到5月1日,最早进入现蕾期的是‘特贾斯’,最晚进入的是‘威奇塔’,两者相差 15 d。经过7~12 d雌花柱头裂开,最早开裂的是‘特贾斯’,最晚的是‘威奇塔’。柱头开裂后的5~10 d进入柱头倒八字型时期,此时雌花进入可授阶段,各品种进入可授期的时间在5月5—15日之间,相差11 d,其中最早的是‘特贾斯’和‘马汉’,最晚的是‘威奇塔’和‘湘林1号’。在雌花柱头湿润期之后5~8 d,各品种雌花柱头干涸,最早进入干涸期的是‘特贾斯’,最晚的是‘福克特’。各品种雌花单花开放持续时间为22~31 d,其中最长的是‘斯图尔特’,由于‘斯图尔特’雌花发育较慢,单花开放持续时间长达31 d,虽现蕾较早,但进入可授期的时间与其他品种相差不大;最短的是‘亚林13号’,雌花单花发育快,有‘晚现蕾早结果’的现象。
2.3 薄壳山核桃花期的气候状况
本研究中薄壳山核桃试验地从雌雄花萌发至花期结束时的天气情况统计如下:3月统计时段有阴6 d、多云2 d、雨10 d、晴4 d、雨夹雪1 d,有降水10 d、无降水21 d;4月有阴11 d、多云7 d、雨8 d、晴4 d,有降水8 d、无降水22 d;5月有阴4 d、多云3 d、雨17 d、晴7 d,有降水17 d、无降水14 d。统计试验地3—5月的降水情况可知,降水主要集中在3月底和5月底,其中,降水量3月为142.6 mm,4月为79.7 mm,5月为217.2 mm。日最高降水量为3月27日(67.1mm),4月中旬和5月上旬降水略有波动。总体而言,调查期总降水量较往年偏少,降水以小雨或阵雨为主;在各品种散粉期间(4月24至5月21日)总降水量为88.9 mm,期间4次较明显的降水时间分别为5月4日、5月5日、5月9日及5月15日,降雨持续时间短、雨后天气转晴温度升高,对散粉及受粉影响不大。
花发育阶段的温度对开花物候期有较大的影响[12],能直接调控开花结实的时间和质量,分析花发育期间的积温,能初步得出促进开花的温度条件,也有利于推测不同地区和不同年份的开花物候期[13-15]。本研究薄壳山核桃试验基地从雌雄花萌发至花期结束时的日平均温度变化情况如图3A所示。试验地3—5月的月平均温度、月平均最低温度及月平均最高温度呈整体上升趋势,在3月中下旬出现一次温度上升又急速下降的现象,对花芽的萌发有一定的影响,但低温持续时间不长(3 d),进入4月后气温略有波动,总体稳步上升,没有倒春寒现象。试验地3月的平均气温为14.1 ℃,平均最低气温为11.2 ℃,平均最高气温为17.6 ℃,最高气温出现在3月21日(24.6 ℃),最低气温出现在3月29日(6.6 ℃),≥10 ℃有效积温达到140.2 ℃;3月气温影响花枝伸长速度,间接影响开花物候期和果实发育的起始时间。试验地4月的平均气温为16.5 ℃,平均最低气温为12.4 ℃,平均最高气温为21.2 ℃,最高气温出现在4月30日(24.7 ℃),最低气温出现在4月11日(9.9 ℃)。4月上旬为雄花萌动期,此阶段积温为192.4 ℃;4月中旬至下旬为雄花序伸长期,雌花进入初花期,此时积温为326 ℃;试验地5月的平均气温为22.4 ℃,平均最低气温为19.4 ℃,平均最高气温为26.3 ℃,最高气温出现在5月4日(28.8 ℃),最低气温出现在5月25日(18 ℃)。在授粉期间(4月30日至5月21日)平均气温为23.6 ℃,平均最低气温为19.8 ℃,平均最高气温为26.3 ℃。5月上旬薄壳山核桃进入散粉始期,雌花进入盛花期,此时积温为458.4 ℃;5月中旬薄壳山核桃散粉基本结束,雌花进入终花期,此时积温为579.7 ℃;5月下旬积温为709.3 ℃,薄壳山核桃雄花进入终花期,雌花子房开始膨大。5月期间未出现极端天气,有利于雄花散粉及雌花受粉。
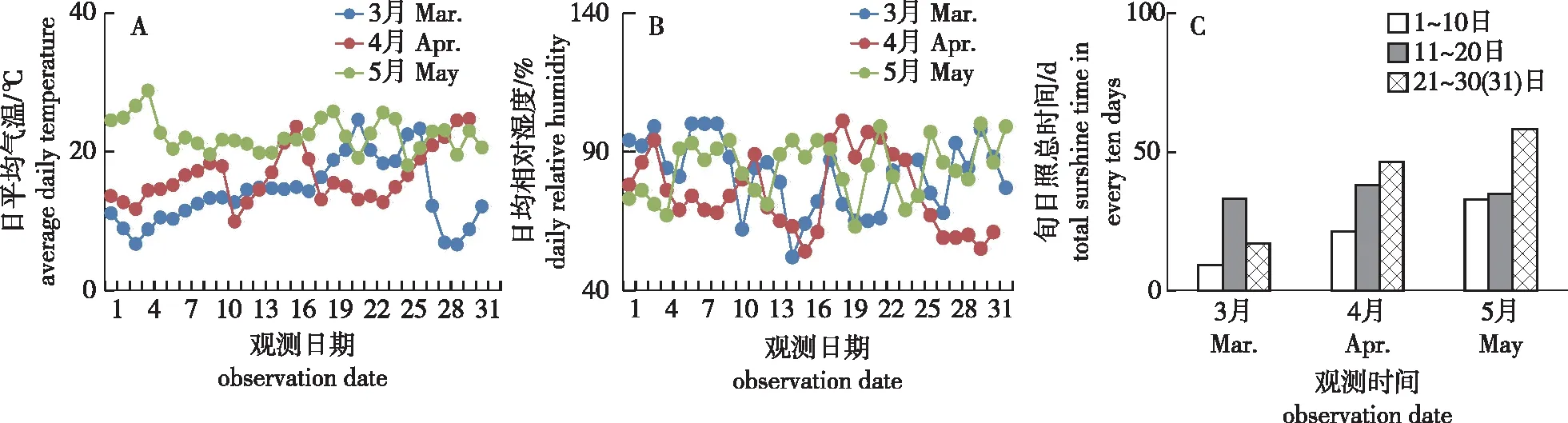
图3 试验基地3—5月日平均气温、相对湿度和旬日照总时间Fig.3 Average daily temperature,daily relative humidity and total sunshine time in every ten days of experiment field from March to May
薄壳山核桃试验基地从雌雄花萌发至花期结束时的日均相对湿度变化情况如图3B所示。试验地3月平均相对湿度为81.6%,4月平均相对湿度为75.2%,5月平均相对湿度为84.2%,3—5月的相对湿度均在52%以上,且随气温、降水量的变化而波动。总体而言,在雄花散粉期间及雌花受粉期间平均相对湿度为78.2%,湿度适宜,既不影响雄花散粉,亦有利于维持雌花可授期柱头湿度。
薄壳山核桃试验基地从雌雄花萌发至花期结束时的日平均温度变化情况如图3C所示。试验基地的月日照总时间在3—5月逐月增加,从图3C可知,3月总日照时间为59.2 h,4月总日照时间为105.8 h,5月总日照时间为126 h;在薄壳山核桃雌雄花开放期间总日照时间达到了291 h。
2.4 薄壳山核桃品种配置
薄壳山核桃是典型的雌雄同株异花植物,雄花着生在1年生枝条的中部或中下部,雌花着生于当年抽生新梢的顶端,雌雄花的萌发及发育的时间不一致,根据雌花可授期和雄花散粉期的差异可分为雌先型、雄先型、雌雄同期型。
由14个薄壳山核桃品种雌雄花开花物候期调查结果(图4)可知:供试材料中‘密西西比’、‘亚林100’、‘威奇塔’、‘斯图尔特’和‘卡多’5个品种的雌、雄花期相遇超过5 d,具有一定的自花授粉能力,可视为雌雄同期型品种;‘福克特’、‘亚林13’、‘特贾斯’和‘马汉’4个品种属于雌先型品种;‘湘林1号’、‘金奥瓦’、‘佩鲁奎’、‘波尼’和‘普兹内’5个品种属于雄先型,雌先型和雄先型品种的雌雄花花期相遇小于4 d或相遇时间远少于雌花可授期持续时间,不能完成自花授粉或授粉效率低,必须配置授粉树。
根据各品种雄花散粉期和雌花可授期进行品种配置,结果如表3所示。表3显示,‘湘林1号’的雌花可授期长达19 d,雄花散粉相遇时间只有8 d,与‘福克特’进行配置可提高授粉率;‘福克特’雌雄花相遇6 d,雌花可授期为13 d,‘特贾斯’是其最佳授粉品种,此外‘湘林1号’搭配‘福克特’或‘威奇塔’也能满足较长时间的授粉需求;‘密西西比’和‘金奥瓦’的可授期一致,‘湘林1号’、‘特贾斯’和‘马汉’能作为授粉树;‘亚林100’具有一定的自花授粉能力,用‘湘林1号’搭配‘亚林100’或‘斯图尔特’等其他品种能提高授粉能力;‘威奇塔’能完成自花授粉,‘福克特’、‘亚林13’、‘斯图尔特’和‘马汉’也能为其授粉;‘特贾斯’和‘马汉’的雌花可授期相对较早,搭配‘金奥瓦’、‘佩鲁奎’、‘波尼’或‘普兹内’都能改善授粉效果;‘斯图尔特’和‘卡多’能完成自花授粉,‘特贾斯’或‘湘林1号’也能作为其授粉树;‘佩鲁奎’、‘波尼’、‘亚林13’、‘普兹内’的最佳授粉树均为‘湘林1号’和‘特贾斯’。
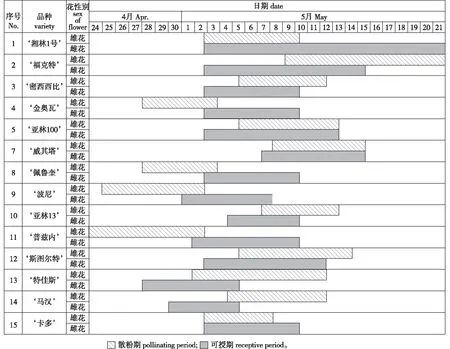
图4 薄壳山核桃雌花可授期与雄花散粉期对应图Fig.4 Female flower receptive periods and male flower pollinating periods of different pecan varieties

表3 14个薄壳山核桃品种的授粉树配置表
3 讨 论
在薄壳山核桃花期发育过程中,花外部形态与开花物候期相对应,不同品种的雌雄花所呈现的外部形态有一些差别。以‘波尼’为代表的品种雄花序较粗,苞片较为膨胀饱满,小花连接紧密;雌花成熟时柱头呈红色圆锥状。以‘福克特’为代表的品种雄花序较为细长,苞片包裹紧密,没有‘波尼’饱满,花序伸长后小花间松散,花轴显露可见;雌花成熟时柱头呈绿色“倒八字”形。花器官外形及物候期特征与外部环境因子和体内基因表达相关[16],这类外形特征既能作为品种特异性状指导品种鉴别,也从另一方面反映出具有相似性状的品种在进化关系上更加接近。
在薄壳山核桃建园品种选择时,以‘波尼’为代表的雄花序粗壮、花苞膨胀饱满连接紧密的雄花量大的品种可作为授粉树的优先选择。此外,选择授粉树时还应尽量筛选出雄花同时存在幼期、中期和盛期且散粉持续时间长的品种,如‘福克特’。‘湘林1号’和‘福克特’现蕾时间可长达13~19 d,在同一株树上有的雌花刚刚现蕾,有些雌花则已进入可授期,这种持续现蕾的特性能使雌花接受不同时期的花粉,对于提高坐果率有很大的帮助,因此在生产上可大量应用‘湘林1号’和‘福克特’为主栽品种,两个品种相互授粉,可最大限度地实现正常授粉条件下的高产。
根据雌花可授期与雄花散粉期的相遇情况,供试的各薄壳山核桃品种雌雄异熟的研究结果与之前的报道一致[17-19],受品种特性、栽种地气候、土壤条件等影响,各品种在不同地区的开花期和花期持续时间有所差别,但雌雄异熟次序不变[20-21]。据文献报道,2012年‘波尼’在南京不同地区分别表现为雄先型和同熟型[9,22];2018年在湖南省邵阳市‘波尼’表现出雌雄同熟[23];云南‘波尼’的可授期与散粉期与南京、湖南以及浙江也存在较大的差异,发生时间提早了12~22 d[24-26]。由于试验材料和试验地点限制,以及观测方法、开花时期温度、降雨、湿度等气候条件的影响,同一品种在不同地域存在差异,同一地区的开花物候期每年也不尽相同[27]。为了更精准地掌握当地的开花物候期,应该连续多年进行花期观测和调查,分析气候因子对花期的影响,并建立模型。
近年来我国的薄壳山核桃种植面积逐年增加,引入及选育的薄壳山核桃品种多达100余种,种植者如何从这些品种中挑选出适宜的品种进行配置是影响果园产量的重要原因。本研究的14个品种中大部分雌雄花花期不遇,大范围人工授粉会消耗大量的人力、物力,操作难度大,这更加凸显出合理配置授粉树重要性。‘密西西比’、‘亚林100’、‘威奇塔’、‘斯图尔特’和‘卡多’为雌雄同熟品种,国内外研究证实自花授粉会导致果仁质量和体积减小、坐果率降低的现象[3],大部分异花授粉的果实表现更佳,因此对雌雄同熟的品种进行品种配置值得推荐。在湖南地区的薄壳山核桃品种授粉配置上可参照本研究提出的授粉组合,结合品种特性、所在地生境条件和气候特征选择其中的一种或几种组合用于建园,达到最优的授粉效果。在可授品种中可通过筛选得出与主栽品种亲和性较高、花粉活力强、花粉量大的授粉树[28],对主栽品种进行授粉杂交实验,根据不同授粉组合的坐果率、单果质量、出仁率、种仁色泽及口感、种仁含油率及营养成分含量等指标进行连续3年以上的综合评定[29-30],可为主栽品种筛选出最佳授粉树。
通过对湖南低山丘陵区薄壳山核桃14个品种花期物候特征的观察,得出以下结论:薄壳山核桃雌花发育期经历雌花现蕾、柱头开裂、柱头湿润、柱头干涸变黑及子房膨大时期。雄花发育期经历雄花序萌发、花序伸长、苞片开裂、花药变黄、散粉、花药变黑及雄花序脱落等时期。雌雄花发育期间≥10 ℃有效积温为458~579.7 ℃时有利于促进开花。‘密西西比’、‘亚林100’、‘威奇塔’、‘斯图尔特’和‘卡多’5个品种雌雄同期,具有一定的自花授粉能力;‘福克特’、‘亚林13’、‘特贾斯’和‘马汉’4个品种属于雌先型品种;‘湘林1号’、‘金奥瓦’、‘佩鲁奎’、‘波尼’和‘普兹内’5个品种属于雄先型。可相互授粉的品种组合有:‘湘林1号’ב福克特’;‘密西西比’ב卡多’;‘金奥瓦’ב特贾斯’、‘金奥瓦’ב斯图尔特’、‘金奥瓦’ב马汉’;‘佩鲁奎’ב特贾斯’、‘佩鲁奎’ב马汉’、‘佩鲁奎’ב斯图尔特’;‘波尼’ב特贾斯’、‘波尼’ב马汉’;‘普兹内’ב特贾斯’、‘普兹内’ב马汉’。‘特贾斯’或‘普兹内’配置‘波尼’和‘威奇塔’能满足绝大部分品种的授粉需求。
猜你喜欢
知识窗(2022年7期)2022-08-05
金沙江文艺(2022年4期)2022-04-26
蚕桑通报(2022年3期)2022-04-10
河北果树(2021年4期)2021-12-02
小天使·三年级语数英综合(2021年4期)2021-06-15
百科探秘·航空航天(2020年2期)2020-03-02
农民致富之友(2018年18期)2018-07-14
小学生作文(中高年级适用)(2018年4期)2018-05-14
大科技(2016年4期)2016-07-13
天然产物研究与开发(2016年6期)2016-06-05
