
3种天牛幼虫蛀食振动小波包特征及其识别*
2021-01-26 00:48:34刘圣煌杨江天崔建新
林业科学 2020年12期
刘圣煌 杨江天 崔建新
(1.北京交通大学机械与电子控制工程学院 北京 100044; 2.河南科技学院害虫天敌繁育研究中心 新乡453003)
天牛是鞘翅目(Coleoptera)叶甲总科(Chrysomeloidea)天牛科(Cerambycidae)昆虫的总称,全世界已知超过2万种,我国有4 000多种(王直诚, 2014),绝大多数为各类树木的蛀干害虫。天牛幼虫在树木内部蛀食树干,通常会引起树木生长受阻,产量减低,树势削弱,寿命缩短。受害严重时,可能导致树木迅速枯萎与死亡。此外,被蛀食的树木容易引起其他害虫及病菌的侵入,易受大风吹折。林木受蛀害后,品相必然会降低,甚至失去观赏性和商品价值。例如栗山天牛(Massicusraddei)在我国东北地区为害栎树每年造成数十亿元经济损失(唐艳龙等, 2011); 云斑天牛(Batocerahorsfieldi)严重为害白蜡(Fraxinuschinensis)、杨树(Populus)、柳(Salix)、榆(Ulmus)、桑(Morus)、核桃(Juglansregia)等林木,对我国城市园林绿化景观造成巨大危害(严敖金等, 1997; 路纪芳等, 2016); 灭字脊虎天牛(Xylotrechusquadrips)为害经济作物咖啡(Coffea),在印度每年造成大约2亿美元的损失(付兴飞等, 2020)。同时天牛幼虫会随着木材的流动在世界范围内迁移,造成生物入侵,对生态环境的稳定造成威胁。天牛不同种间的生物学习性差异很大,如栗山天牛在辽宁3年1代,云斑天牛在华北地区2年1代,灭字脊虎天牛在云南1年1代或2代(肖刚柔, 1991; 唐艳龙等, 2011; 杨文波等, 2017)。天牛幼虫存活在树干内部,传统的农药毒杀防治效果不佳而且污染环境,因而采用寄生蜂进行生物防治是常用的控制方法。利用肿腿蜂防治天牛幼虫是我国最为重要的生防措施之一,在防治栗山天牛时,释放白蜡吉丁肿腿蜂(Sclerodermuspupariae)防治的最佳时期是每年的8—9月和次年的5—6月,栗山天牛的幼虫龄期分别为1龄和2龄,寄生率和致死率较高,防治效果最好(唐艳龙等, 2014)。而利用川硬皮肿腿蜂(Sclerodermussichuanensis)防治云斑天牛的最佳时间是6月中旬至7月中旬,此期云斑天牛的幼虫龄期适宜(杨桦等, 2011; 梅增霞等, 2014; 肖刚柔, 1991)。由于利用寄生蜂只能针对某一种天牛或某些类天牛的特定发育阶段寄生。因此,在不破坏树干的前提下,确定天牛幼虫的种类,了解其生物学习性,可以准确地指导适时防治,是开展天牛类隐蔽蛀干害虫科学防治的关键技术。
目前常用的蛀木害虫检测方法仍为目视、剖检、敲击等人工检测手段(潘杰等, 2013)。此种方法费时、费力、效率低。同时由于检测方法没有工程化,准确性受检测人员的专业技术水平的限制。目前常用的工程化检测方法有X光检测法和声音检测法。X光检测法存在以下问题: 1) 设备笨重且价格昂贵,不适合野外使用; 2) 难以确定天牛的种类; 3) X射线对人体有一定的伤害。利用害虫发出的声音识别害虫种类已经有成功的报道,卜宇飞等(2016)利用Adobe Audition 3.0和MATLAB完成了对7种害虫声音信号的采集分析处理,有效提取了害虫声音特征。Mankin等(2008)在消音箱内采集害虫发出的声音信号,进行频谱分析,成功识别出树干内天牛幼虫种类。但害虫发出的声音信号信噪比低,且易受到环境噪音干扰,实际应用受到很大限制。本文提出了通过采集分析幼虫蛀食树干产生的振动信号,检测天牛种类方法,克服了声音检测方法的缺点,同时具有价格低廉、检测时对树木伤害小的突出优点,有很好的应用前景。
1 材料与方法
1.1 材料
试验树木及天牛幼虫种类见表1。试验地点为河南省新乡市人民公园,测试时间为2017年7月18日,采样时气温约为33 ℃,当天气温均值28 ℃。

表1 试验树木及天牛幼虫种类Tab.1 Species of trees and longicorn larvae
1.2 方法
振动传感器采用江苏联能电子有限公司生产的YD-189型压电式加速度计,灵敏度5.015 V·g-1,频率范围0.2~5 000 Hz,最大横向灵敏度<5%,量程±5 g。数据采集器为基于NImyDAQ开发的便携式测振仪,适合野外环境下使用。
信号采集选择无风(至少没有强风)晴朗天气条件下进行,这是因为大风会造成林木树干晃动,对害虫蛀木振动信号产生干扰。天牛幼虫蛀木振动信号采集办法如下:
1)为了保证试验的准确性,应提前进行调查,选择只有一种天牛幼虫的树木进行试验。挑选粪孔附近的树干表面,削去树皮并刮出一个φ20 mm的平面,用502胶将传感器黏接在树干上。
2)用便携式检测仪采集天牛蛀木振动信号。设定采样频率10 000 Hz,每段采样时间120 s。程控放大增益为小树(周长小于80 cm)200倍,大树(周长大于80 cm)400倍。这样选择程控增益放大倍数的目的是尽量将微弱的蛀木振动信号放大到数据采集器满量程以下,半量程以上。每种树采集20棵的样本。
采集数据以后,解剖树干,如果树干内有1只以上天牛幼虫或其他蛀干害虫,将这组数据剔除,即保证分析的数据都只有1只天牛幼虫。对各种天牛幼虫初步鉴定后,分别罩笼饲养,直至饲养出成虫完全确定种类为止。天牛成虫的监测常年进行,种类监测结果与室内罩笼饲养结果相互确认。
2 结果与分析
2.1 数据筛选
卜宇飞等(2017)用红外摄像机观测天牛幼虫在蛀道内的活动,同时采集幼虫发出声音信号,找到了天牛幼虫活动的声音信号特征(表2)。“振是声之源”——振动挤压周围的空气,产生声音。天牛幼虫活动声音信号和振动信号有同样的特征(刘圣煌, 2019)。根据这一原理进行振动信号筛选,天牛幼虫在蛀道内活动引发的树干振动信号由一系列冲击振荡组成,如图1所示。从表2中可以看出,外界干扰与天牛幼虫活动引起的振动在波形和相对幅值上存在比较大的差别,可以据此进行筛选,找出天牛幼虫蛀食振动。
图2为光肩星天牛幼虫的蛀食树干振动信号,图3为由于风引发的树干振动,图4未定性的树干振动。可以看出,图2、3、4的波形明显不同。通过此种方法可以筛选出由天牛幼虫活动所产生的振动信号。
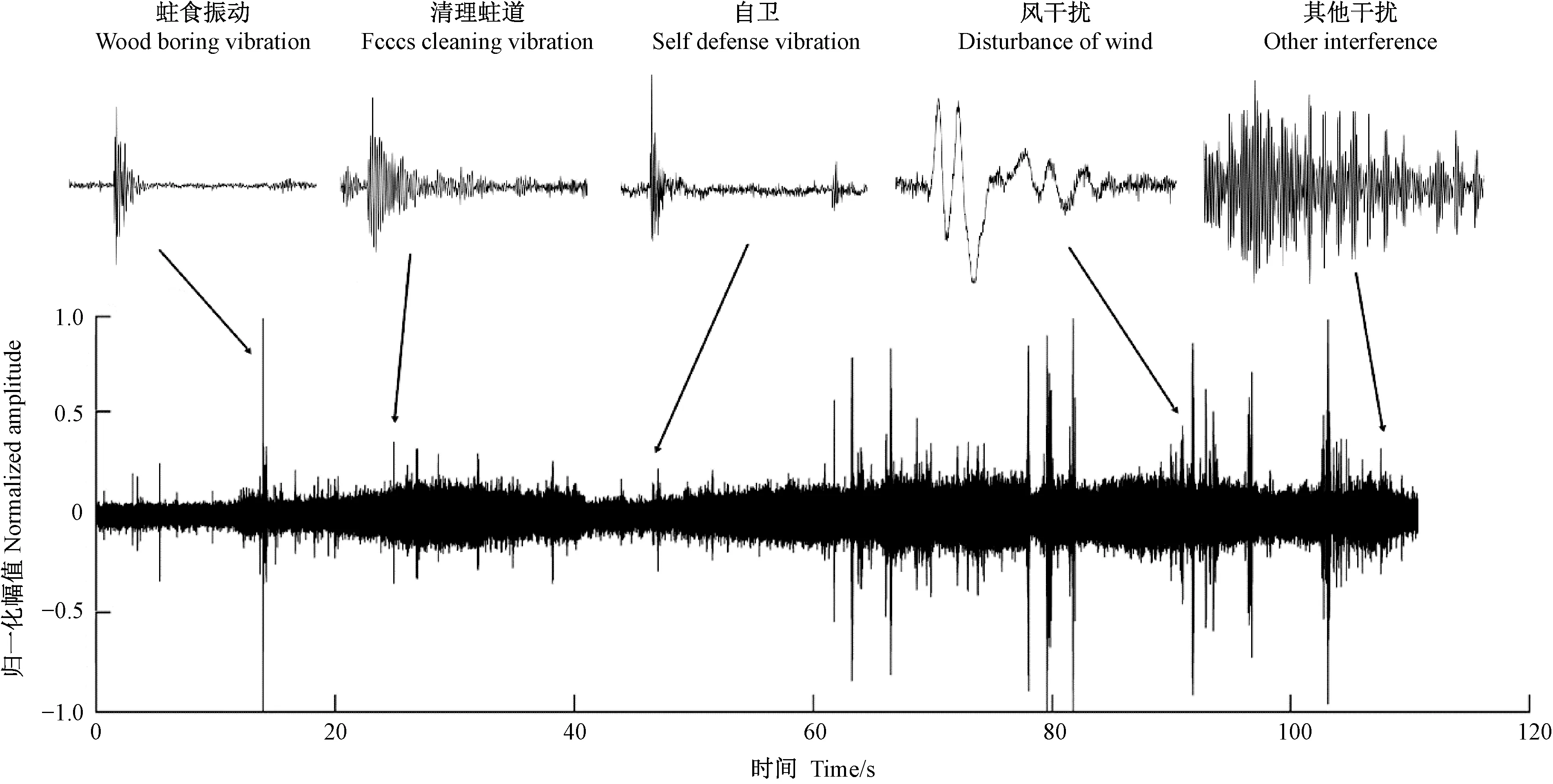
图1 天牛幼虫活动引起的树干振动信号Fig.1 The vibration signal of tree trunk caused by longicorn larvae

表2 天牛幼虫行为占比及特征①Tab.2 Behavioral proportions and characteristics of longicorn larvae

图2 光肩星天牛蛀食二球悬铃木树干振动信号波形Fig.2 The vibration signal of Anoplophora glabripennis larva boring tree trunk of Platanus acerifolia
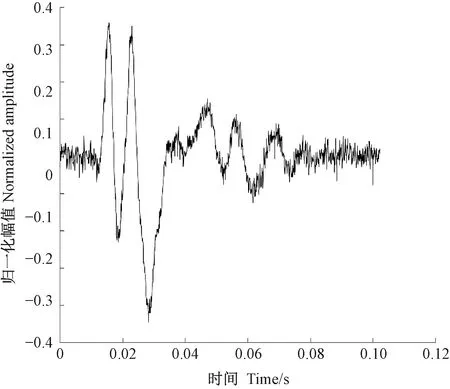
图3 风吹引发的树干振动信号波形Fig.3 The vibration signal caused by wind blows
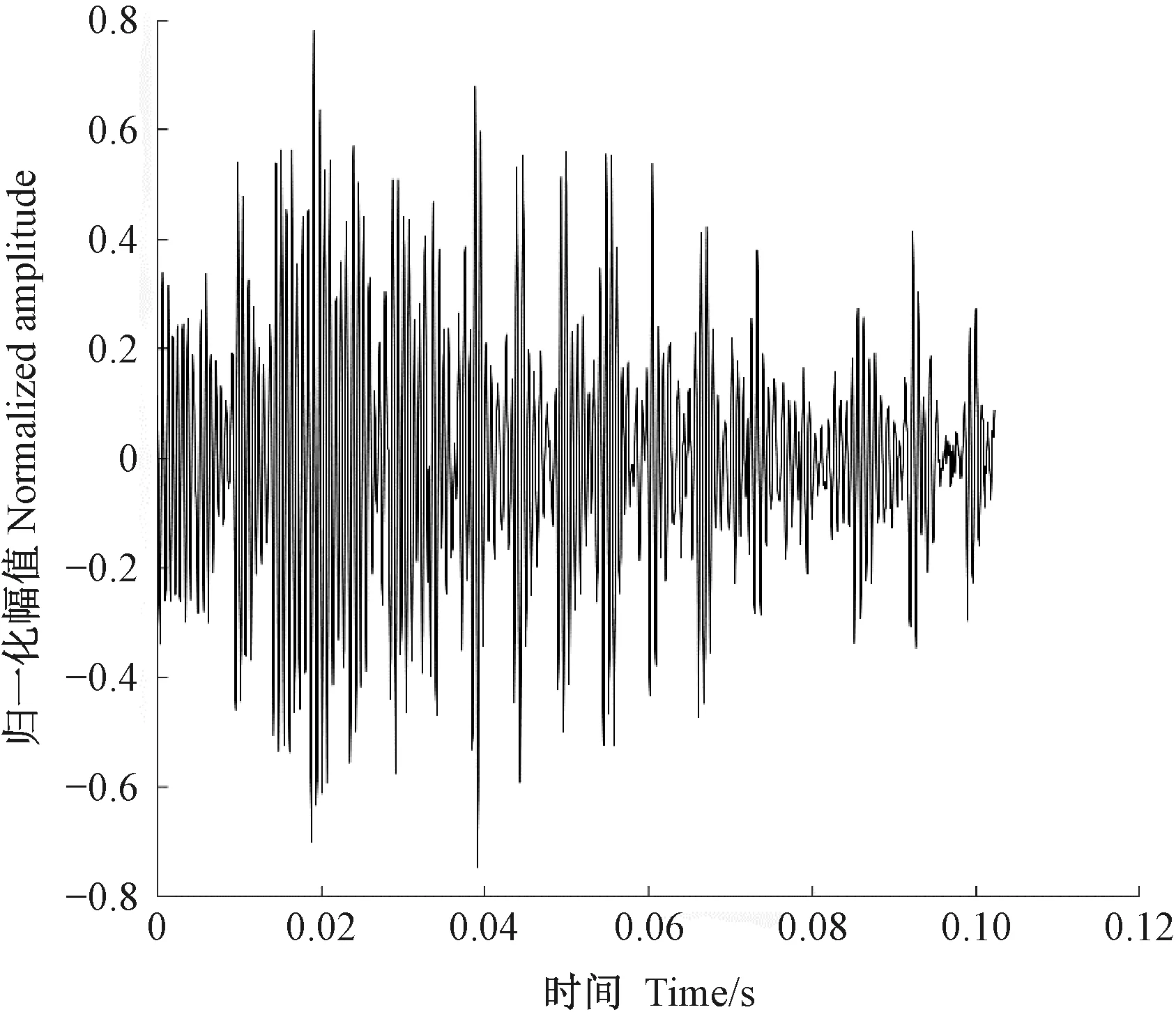
图4 未定性树干振动波形Fig.4 The undetermined trunk vibration waveform
天牛幼虫在蛀道内除了蛀食行为外,还存在清理蛀道、自卫、爬行等其他的行为,这些行为都会引起树干的振动。分析蛀食振动信号需要排除天牛幼虫蛀食以外活动引起振动的干扰。相关研究(卜宇飞等, 2017)表明,天牛幼虫在蛀道内不同活动引起树干振动的相对幅值和波形存在差异。蛀食振动引起的冲击最大,上升沿一个非常陡的脉冲,然后迅速衰减,持续时间短,最大的相对振幅可达0.8,明显大于其他3类行为引起的振动; 天牛幼虫在蛀道内爬行引起的振动持续时间较长且不规律,振幅有波动,相对振幅一般不超过0.2,在4类行为中最小; 天牛幼虫清理蛀道引起的树干振动存在一个较大的冲击,相对振幅值可达0.5~0.6,仅次于蛀食振动; 天牛幼虫自卫引起的振动波形间隔时间极短,一般小于50.0 ms,连续出现,振幅大于爬行引起的振动,相对振幅可达0.4。根据上述特点,筛选天牛幼虫不同行为引起的树干振动。图5为天牛幼虫蛀食树干振动信号波形,图6为幼虫清理蛀道树干振动信号波形,图7为天牛幼虫自卫引发的树干振动波形。由于爬行引发的振动最大相对振幅值为0.2,未采集到振动波形。

图5 云斑天牛幼虫蛀食白蜡树树干振动信号波形Fig.5 The vibration signal of Batocera horsfieldi larva boring tree trunk of Fraxinus chinensis
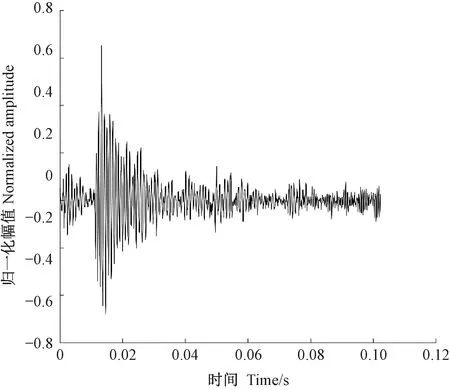
图6 云斑天牛幼虫清理白蜡树树干蛀道振动信号波形Fig.6 The vibration signal of Batocera horsfieldi larva cleaning feces in tree trunk of Fraxinus chinensis

图7 云斑天牛幼虫在白蜡树树干内自卫活动的振动信号波形 Fig.7 The vibration signal of self-defense of Batocera horsfieldi larva in tree trunk of Fraxinus chinensis

图8 云斑天牛蛀食白蜡树树干振动信号变分模态分解Fig.8 The variational mode decomposition of the vibration signal of Batocera horsfieldi larva boring tree trunk of Fraxinus chinensis
2.2 振动信号降噪
使用变分模态分解(variational mode decomposition,VMD) (Dragomiretskiyetal., 2014) 对采集到的害虫蛀木振动信号进行降噪处理。其原理是先将原始信号分解成一系列本征模态函数(intrinsic mode function,IMF),检测各本征模态函数找出噪声分量,滤除噪声分量后重构原信号。使用中心判别法实验确定变分模态分解最佳模态个数为8,惩罚因子为2 000。本文采用波形与时间熵判别法识别噪声和有用信号。
截取天牛幼虫蛀食振动信号前1 000个采样点的数据进行分析,采样频率为10 000 Hz,因此实际振动信号的长度约为0.1 s(100 ms)。害虫蛀食振动信号通常在短时间(30 ms)内迅速衰减,但是噪声信号幅值不会很快衰减。根据这一性质,观察变分模态分解得到的各本征模态函数的时域波形,如果幅值变化大就是有用信号,反之就是噪声。
图8为云斑天牛(为害白蜡)蛀干振动信号做变分模态分解得到8个本征模态函数。容易看出,IMF1—IMF6 波形迅速衰减,为有用信号; 而 IMF7、IMF8波形衰减很小,是噪声。同时计算各分量的时间熵分别为2.25、2.02、1.79、1.77、1.69、1.76、2.96和3.00。熵是信号混乱程度的度量,熵值越大,说明信号越混乱无规律。据此也可以判断IMF7和IMF8 分量为噪声。
2.3 时域特征与频域特征
选择天牛幼虫蛀木振动信号的波形和持续时间作为时域特征,信号能量随频率的分布作为频域特征识别天牛种类。对VMD降噪后的振动信号进行3层小波包分解,小波基函数选用滤波器截止性能最好的dmey小波。对各节点小波包系数做傅里叶变换,计算各小波节点的频域能量,以各节点频域能量在总能量中的占比作为频域特征参数。图9为云斑天牛(为害白蜡树)的蛀食树干振动信号小波包分解,左侧为各结点小波包系数,右侧对应的功率谱。从图中可以看出,前面4个节点信号幅值比后面4个节点信号的幅值大很多,信号能量主要集中在前4个节点,对应频带 0~2 500 Hz。因此本文以前4个节点的能量作为特征,判断天牛幼虫种类。小波包分解各个节点所对应的频带见表3。

图9 云斑天牛蛀食白蜡树树干振动信号小波包分解Fig.9 The wavelet packet decomposition of the vibration signal of Batocera horsfieldi larva boring tree trunk of Fraxinus chinensis

表3 蛀木振动信号小波包分解各结点对应频带Tab.3 Frequency range of wavelet packet nodes
2.4 3种天牛蛀食振动时域特征与频域特征
天牛幼虫在蛀道内活动引起的树干振动信号由一系列频率、幅值差别较大的冲击信号组成,成分复杂。分析这些振动信号是检测天牛种类的基础。在天牛幼虫活动行为中,蛀食树干所占比例最大,引起的振动相对幅值也最大,因此可通过分析天牛幼虫蛀干振动信号识别天牛种类。天牛幼虫蛀干振动信号的上升沿是一个陡的脉冲,与天牛幼虫蛀食树木相对应。随后能量被树干结构阻尼消耗,脉冲以近似指数包络线衰减。蛀木振动信号特征与天牛种类、龄期、个体大小,以及寄主树木材质、水分、粗细、密度等诸多因素有关。光肩星天牛、云斑天牛与桃红颈天牛蛀食二球悬铃木、白蜡树、紫叶李和西府海棠树干的振动信号具有不同的时域与频域特征。图10—13分别为云斑天牛蛀食白蜡树干、光肩星天牛蛀食二球悬铃木树干、桃红颈天牛蛀食紫叶李树干、桃红颈天牛蛀食西府海棠树干的振动信号波形。从图中可以看出,天牛幼虫蛀食树干振动的4种信号都是衰减震荡,但是各自波形特征、频域特征、能量分布不同。这些差别是识别不同种天牛的依据。表4—6分别为4种信号时域特征的对比、频域特征对比、小波包分解各节点能量对比。

图10 云斑天牛幼虫蛀食白蜡树树干的振动信号Fig.10 The vibration signal of Batocera horsfieldi larva boring tree trunk of Fraxinus chinensis
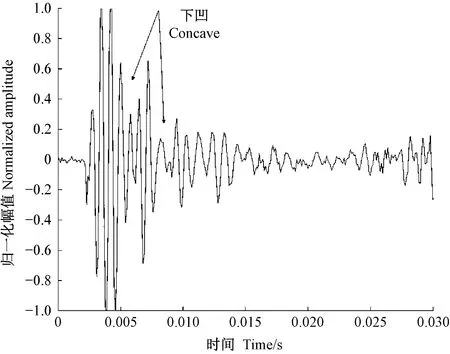
图11 光肩星天牛幼虫蛀食二球悬铃木树干的振动信号Fig.11 The vibration signal of Anoplophora glabripennis larva boring tree trunk of Platanus acerifolia

图12 桃红颈天牛蛀食紫叶李树干的振动信号Fig.12 The vibration signal of Aromia bungii larva boring tree trunk of Prunus cerasifera

图13 桃红颈天牛蛀食西府海棠树干的振动信号Fig.13 The vibration signal of Aromia bungii larva boring tree trunk of Malus micromalus
从表4可以看出,不同种类的天牛幼虫在不同的寄主中的蛀干振动波形和持续时间存在明显差异; 从表5可知,不同种类的天牛幼虫在不同树木内的蛀干振动信号频域特征也不相同; 表6中,不同种类天牛幼虫蛀木振动信号小波包分解各节点能量分布有明显不同。时域特征与频域特征相结合,可准确判断天牛幼虫的种类。
3 讨论
天牛幼虫蛀食树干所引起的振动信号与木材成分、树干含水量、树干被蛀情况等多种因素都有关。本次试验选取河南省新乡市人民公园的树木作为研究对象,同种树木的树龄相同,树木生长环境基本一致。在此种情况下,先使用变分模态分解对采集得到的振动信号进行降噪处理,然后对消噪后的信号做小波包3层分解,成功提取到了天牛幼虫蛀干振动信号的时域特征和频域特征,可用于天牛种类的判别。

表4 3种天牛幼虫蛀食树干振动信号时域特征①Tab.4 Time-domain characteristics of the vibration signal of three longicorn larvae boring tree trunks

表5 3种天牛幼虫蛀食树干振动频域特征Tab.5 Frequency-domain characteristics of the vibration signal of three longicorn larvae boring tree trunks
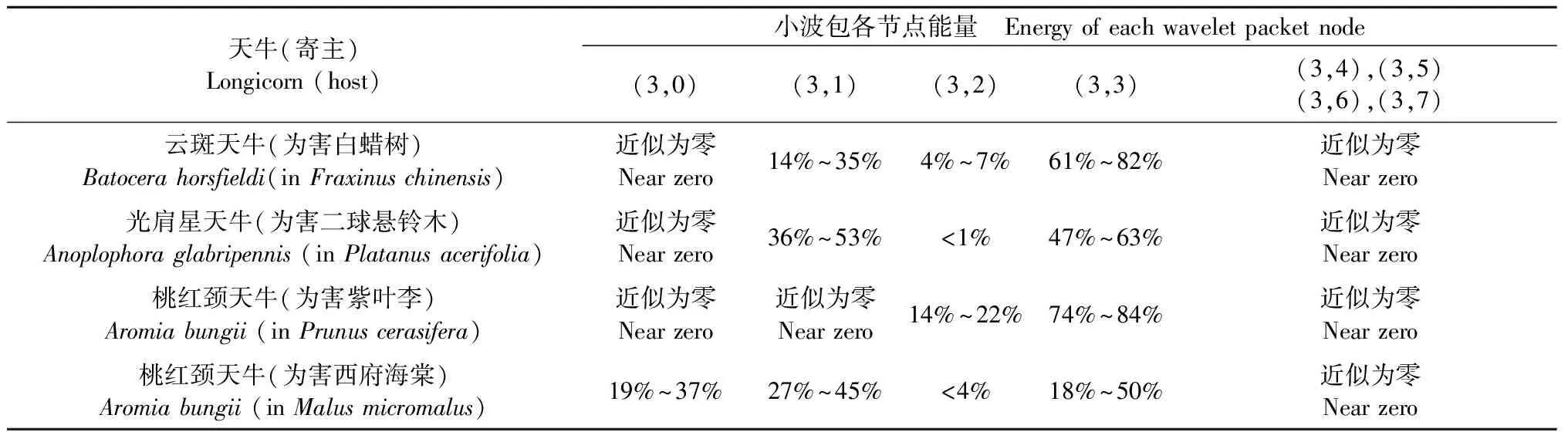
表6 3种天牛幼虫蛀食树干振动信号小波包分解各节点能量Tab. 6 Energy of each wavelet packet nodes of the vibration signal of three longicorn larvae boring tree trunks
天牛幼虫在蛀道中的行为复杂,包括蛀食树干、清理蛀道、自卫、爬行等。其中蛀食树干为咬断木质纤维的过程(Mankinetal., 2008),振动信号能量显著高于其他行为,易于提取分析(卜宇飞等, 2017); 其余行为需要改进检测仪器,开展针对性的行为观测,提高观测精度,才能进一步开展有效的分析研究。
天牛幼虫的龄期与蛀干为害的强度有密切关联,天牛幼虫龄期对幼虫上颚的长度和宽度有显著影响(张海滨等, 2011)。本研究开展的时期在7月中旬,试验中桃红颈天牛和云斑天牛为3~4龄,光肩星天牛为低龄幼虫,野外环境下不同的寄主和营养条件对幼虫龄期的准确判断造成很大困难,相关天牛幼虫的龄期差异对蛀食振动信号的影响也需要后续研究持续关注。
4 结论
本文提出了基于害虫蛀食树干振动信号分析检测识别天牛幼虫种类的方法。该方法先用变分模态分解对采集到的振动信号进行降噪处理,再从天牛幼虫活动引起的振动信号中挑选出蛀食树干振动信号进行分析,利用小波包3层分解提取蛀干振动信号时域与频域特征。 云斑天牛、光肩星天牛、桃红颈天牛幼虫蛀食二球悬铃木、白蜡树、紫叶李和西府海棠树干的振动信号时域与频域特征存在明显差别,采用振动信号分析法能有效识别判断天牛幼虫种类。本研究为建立天牛幼虫蛀木振动信号特征库,实现天牛幼虫活体快速林间检测,制定林区天牛类蛀干害虫防控方案提供了科学依据。
猜你喜欢
种子(2023年5期)2023-08-01 06:54:36
——以蛀食类害虫为例
高教学刊(2022年30期)2022-10-30 02:02:48
小哥白尼(野生动物)(2021年1期)2021-07-16 08:02:52
小学生必读(低年级版)(2018年10期)2019-01-04 10:30:56
测控技术(2018年8期)2018-11-25 07:42:08
故事作文·低年级(2018年10期)2018-10-25 20:56:52
电测与仪表(2016年18期)2016-04-11 11:30:44
江西通信科技(2015年3期)2015-12-05 05:52:10
作文与考试·小学低年级版(2015年11期)2015-07-17 01:02:16
振动、测试与诊断(2014年4期)2014-03-01 01:14:06
