
横断山脉中部川滇冷杉和丽江云杉径向生长对气象因子的响应*
2021-01-26 00:48陈佳佳刘国华王永平李俊清刘琪璟
林业科学 2020年12期
于 健 陈佳佳 周 光 刘国华 王永平 李俊清 刘琪璟
(1. 江苏农林职业技术学院风景园林学院 句容 212400; 2. 北京林业大学林学院 北京100083;3. 江西省林业科学院 南昌 330032)
气候变暖是全球变化的重要驱动力,它可影响植物的生长、繁殖和分布进而控制CO2的吸收和排放,影响全球碳循环模式(Toledoetal., 2011)。过去几十年中,大多数森林在物种组成和分布以及生产力等方面均发生了与气候变暖有关的显著变化(刘国华等, 2001; Trumboreetal., 2015)。理解气候变化和森林植被之间的关系,对评估气候变化对森林生态系统影响尤为重要(范广洲等, 2008)。
树木径向生长是最易识别和观察的特征之一,受气候变化影响很大。树木年轮能提供树木径向生长模式、物种行为、树龄和树木生命周期的环境响应以及对未来气候变化和森林动态的可能反应等丰富信息(Fritts, 1976)。探讨气候变化对树木径向生长的影响一直是树木年代学研究中的一个重要课题。近年来,世界各地开展了有关树木年轮与气象因子关系及古气候重建的大量研究(Liangetal., 2009; 于健等, 2016)。相比而言,同一生境下不同树种的径向生长对气象因子的响应比较仍备受关注(Pedersonetal., 2004; 赵志江等, 2019),同一地区不同树种的气候变化响应可能不同甚至相反(Lyuetal., 2017)。开展上述研究,可提供不同物种间生长趋势和特性的比较,以便预测未来树木组成和物种共存的改变,提高对区域森林生态系统动态变化的理解。

1 研究区概况
研究区地处横断山脉中部,位于青藏高原东南部,通常为四川、云南两省西部和西藏自治区东部南北向山脉的总称。因“横断”东西间交通得名。横断山脉是世界上年轻山群之一,平均海拔4 000 m。横断山脉也是开展全球生物多样性研究的热点地区,植物区系丰富。此外,4条亚洲主要河流(金沙江、澜沧江、怒江和独龙江)流经该山脉。该地区气候主要受高空西风环流、印度洋和太平洋季风环流的影响,冬干夏雨,干湿季非常明显。以香格里拉气象站为例,该区年均气温 6.0 ℃,1月最冷(-2.9 ℃),7月最热(13.7 ℃),年均降水量624.7 mm,73%的降水量集中在6—9月。土壤类型以各种属于棕壤系列的土壤为主(李锡文等, 1993)。
2 研究方法
2.1 样芯采集和年表建立
2009年6月,在横断山脉中部小中甸吉沙村神山原始林内(99°48′40″ E,27°26′07″ N,海拔3 340 m),选择人为干扰较少的区域,采集树木年轮样芯。林分郁闭度约为0.8。该地区主要乔木树种为川滇冷杉和丽江云杉,其他乔木树种主要包括花楸树(Sorbuspohuashanensis)、稠李(Padusavium)、三角槭(Acerbuergerianum)和红桦(Betulaalbosinensis)等。林内灌木主要有忍冬(Lonicerajaponica)、日本小檗(Berberisthunbergii)、刺蔷薇(Rosaacicularis)和杜鹃(Rhododendronsimsii)等。随机选取位于林冠上层、胸径较大且生长健康的川滇冷杉和丽江云杉,利用树木生长锥(内径5.15 mm)在树干胸高位置钻取完整的树木年轮样芯(尽可能通过髓芯)。为了减少对树木生长造成伤害,每株树仅钻取一根树轮样芯。川滇冷杉和丽江云杉分别钻取了45和65根样芯。将所取样芯装入自制纸筒内,对样芯编号,并记录每株树的树种、胸径及生长情况等信息。
野外采集的样芯被带回实验室,经过粘贴和打磨等常规预处理。利用LINTABTm6.0年轮宽度测量仪量测年轮宽度,精度为0.001 mm。运用COFECHA程序(Holmes, 1983)对交叉定年和测量结果进行检验,剔除与主序列相关性差以及难以交叉定年的样芯(川滇冷杉和丽江云杉分别剔除了3和4根),将保留的样芯用于年表建立。年表建立采用ARSTAN程序完成。采用以2/3年龄为步长的样条函数法去除树木本身的遗传因子和干扰竞争产生的生长趋势(即去趋势),并对去趋势的序列以双重平均法合成川滇冷杉和丽江云杉的标准年表(图1),用于建立标准年表的样芯数量分别为42和61根。
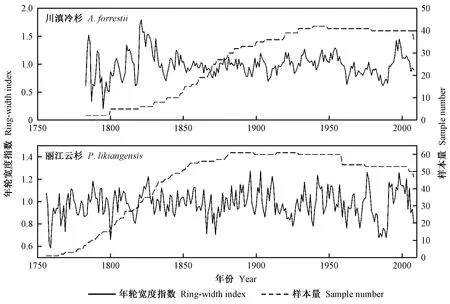
图1 横断山脉川滇冷杉和丽江云杉标准年表序列及样本量Fig.1 Standard chronologies and number of the core sampled of A. forrestii and P. likiangensis in Hengduan Mountains
2.2 气象数据与统计分析
选取距采样点最近、直线距离46.3 km的香格里拉气象站(99°42′ E, 27°50′ N, 海拔3 276.7 m)的1958—2015年的气象数据(取于国家气象科学数据中心http:∥data.cma.cn/)。气象要素包含月均气温(Tm)、月均最高气温(Tmax)、月均最低气温(Tmin)和月均降水量(Pm)。由图2可知,该地区气候变化的特点是雨热同期。由图3可以看出,过去58年间,平均气温不断升高,降水量微弱增加,二者的倾向率分别为0.037 ℃·a-1和0.056 mm·a-1。
分别对川滇冷杉和丽江云杉标准年表与逐月气象因子进行响应函数分析,明确径向生长与气象因子间的关系。考虑到上年气象条件可能对当年树木生长产生影响,故选用上年5月到当年9月(共计17个月)的气象数据与标准年表进行分析。此外,为进一步确认这种关系,还计算了标准年表与逐月气象因子的响应函数一阶差。同时,还采用滑动响应分析,研究径向生长与气象因子的动态变化关系。滑动分析选择的滑动区间为35年,从1958年开始每35年计算1次标准年表与气象因子的响应系数,逐渐依次向后滑动。
2.3 数据处理
采用DendroClim2002程序(Biondietal., 2004)对标准年表与气象因子进行响应函数分析,该软件采用拔靴法1 000次重采样计算响应系数,并检验其在0.05水平下的显著性。为了评估标准年表的大范围空间代表性,利用标准年表与CRU TS3.1(0.5°×0.5°)格点数据进行空间相关分析(http:∥www.climexp.knmi/nl)。使用SigmaPlot 10.0 软件作图。
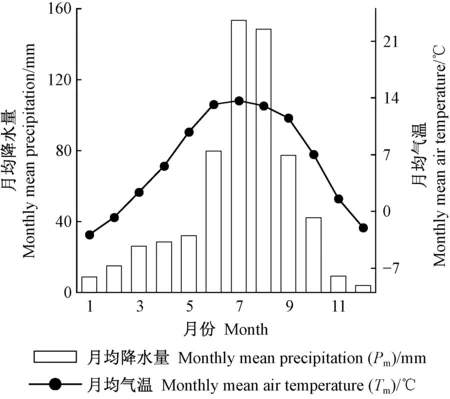
图2 1958—2015年研究区月均气温和月均降水量Fig.2 Monthly mean air temperature and precipitation in the study area from 1958 to 2015
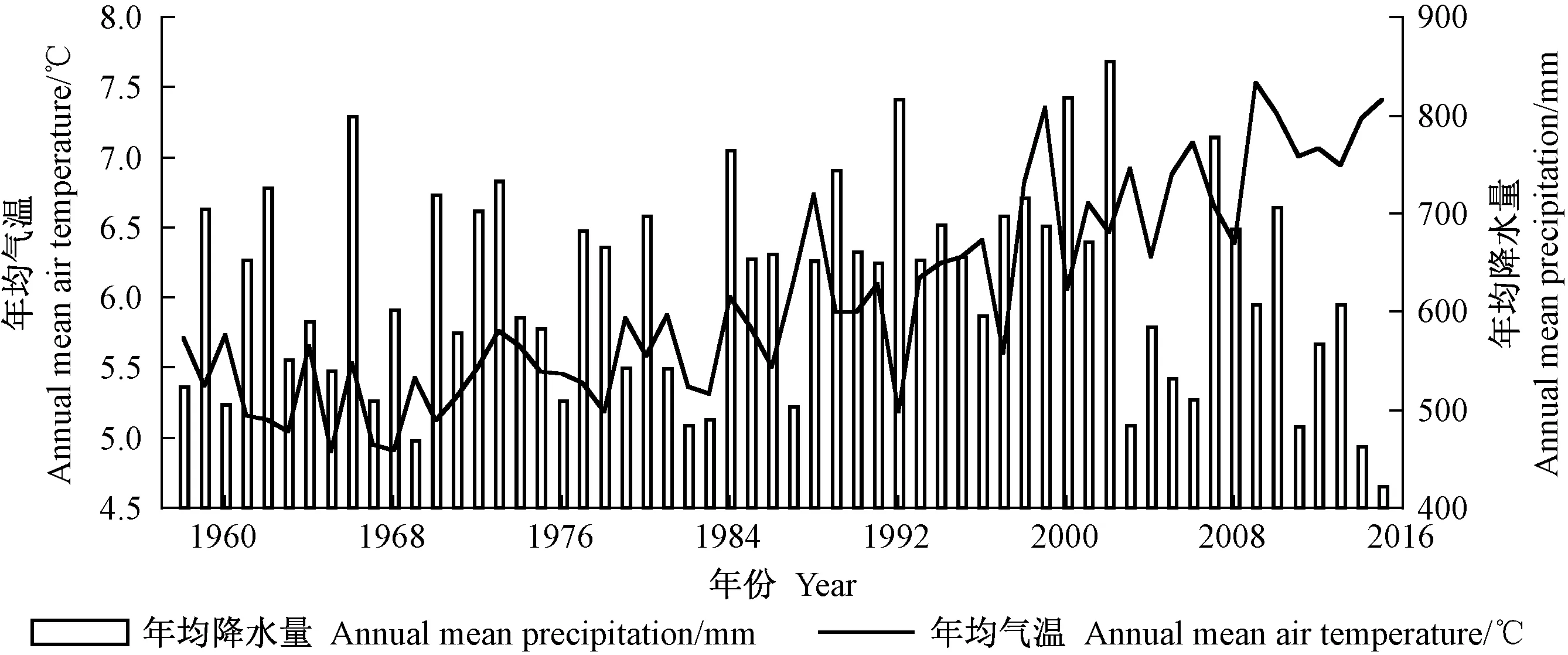
图3 1958—2015年研究区年均气温和年均降水量Fig.3 Annual mean air temperature and precipitation in the study area from 1958 to 2015
3 结果与分析
3.1 年表的统计特征
川滇冷杉标准年表时间跨度为226年(1783—2008年),公共区间为120年(1888—2007年),丽江云杉年表时间跨度为253年(1756—2008年)和148年(1856—2003年)。由表1可知,川滇冷杉年表的平均敏感度、标准差、一阶自相关和第一主成分方差解释量均高于丽江云杉年表。丽江云杉的信噪比也较高(15.19)。上述2个树种的样本总体代表性分别为0.93和0.94,均超过了样本总体代表性(EPS)可接受的临界阈值0.85(Wigleyetal., 1984)。通常而言,高质量的树轮宽度年表具有较高的标准差、平均敏感度、信噪比和样本总体代表性(吴祥定, 1990a)。综上表明,川滇冷杉和丽江云杉标准年表对气候变化响应均具有一定的敏感性,并且川滇冷杉年表在多数统计特征值上均优于丽江云杉年表,表明川滇冷杉对气候变化的响应较丽江云杉更敏感,更适合用于树轮气候学研究。
3.2 年表与逐月气象因子的相关关系
响应函数分析表明(图4),限制川滇冷杉和丽江云杉径向生长的气象因子存在一定差异。具体表现为,川滇冷杉径向生长主要与上年11月和当年8月平均气温显著正相关(P<0.05),与上年11月和当年4月最高气温显著正相关(P<0.05),与上年5月最低气温显著正相关(P<0.05),与当年3月降水量显著正相关(P<0.05)。然而,丽江云杉径向生长与上年11月平均气温显著正相关(P<0.05),与上年7月最低气温显著负相关(P<0.05),与当年3月降水量显著正相关(P<0.05),与各月最高气温均未表现出显著相关关系(P>0.05)。标准年表与逐月气象因子的响应函数一阶差表明(图5),川滇冷杉径向生长与上年11月平均气温和最高气温显著正相关(P<0.05),丽江云杉径向生长与上年11月平均气温显著正相关(P<0.05),这也进一步证明响应函数分析中得出的上年11月气温是限制横断山脉中部地区树木径向生长的主要气象因子这一结论。
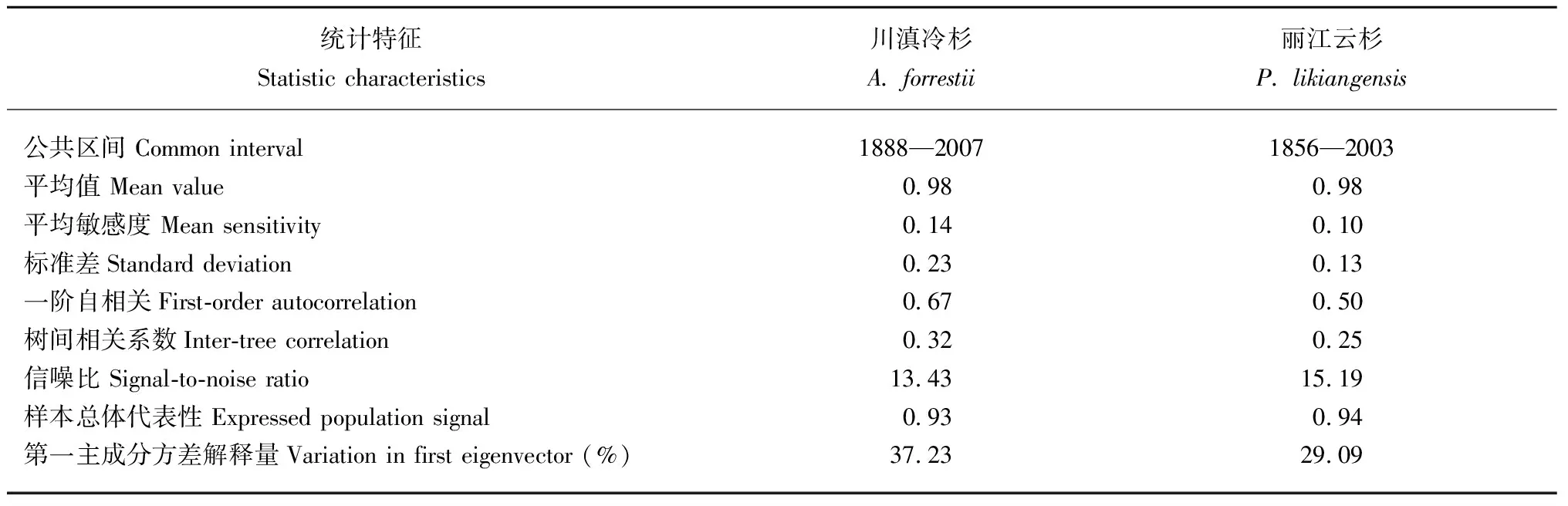
表1 川滇冷杉和丽江云杉标准年表的统计特征及共同区间分析Tab. 1 Statistical characteristics of A. forrestii and P. likiangensis and common intervals analysis
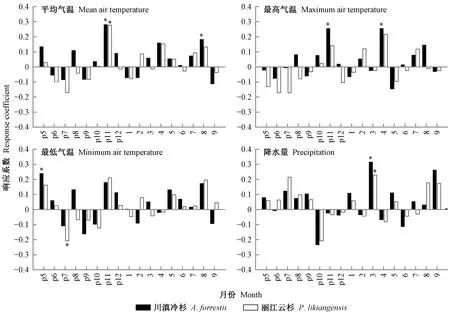
图4 横断山脉川滇冷杉和丽江云杉年表与气象因子的响应函数分析Fig.4 Response function analysis of standard chronology for A. forrestii and P. likiangensis in relation to climatic factors in Hengduan Mountainsp: 上年Previous year; *: P<0.05. 下同The same below
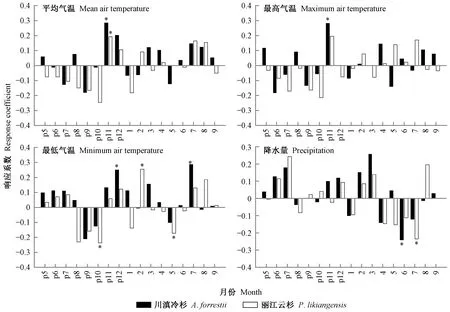
图5 横断山脉川滇冷杉和丽江云杉年表与气象因子的响应函数一阶差分析Fig.5 First-order difference response function analysis of standard chronology for A. forrestii and P. likiangensis in relation to climatic factors in Hengduan Mountains
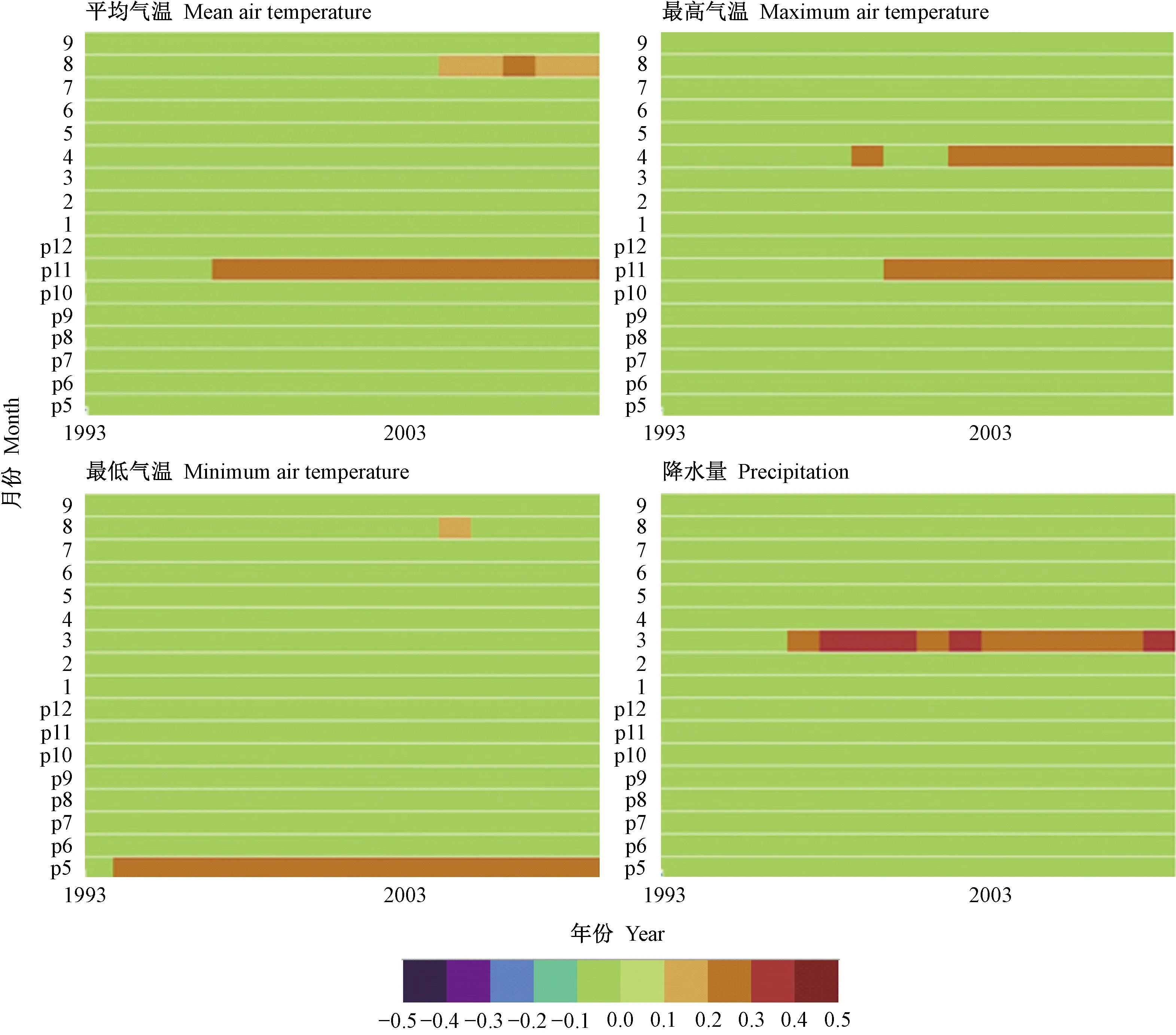
图6 川滇冷杉与气象因子的滑动响应关系分析Fig.6 Moving response analysis of A. forrestii with climate factors
3.3 年表与逐月气象因子的动态关系
为评估长期气候变化对川滇冷杉和丽江云杉径向生长的影响,通过窗口为35年的滑动响应函数研究2个树种径向生长随逐月气象因子变化的动态关系。由图6看出,上年11月平均气温、最高气温及上年5月最低气温对川滇冷杉径向生长的影响具有一定的稳定性,尤其是上年5月最低气温在整个滑动时间序列呈显著正相关并保持较高稳定性。川滇冷杉与当年3月降水量也存在显著正相关,但相关关系在一定时间间隔内存在波动。由图7可知,上年11月平均气温对丽江云杉径向生长的促进作用具有一定稳定性,在1966—2001和1970—2005年滑动区间内响应关系增强。然而,丽江云杉与最低气温的响应关系仅在个别月份个别时间段达到显著水平。
3.4 大尺度区域气候信号
由图4、5可知,上年11月气温是限制横断山脉中部地区川滇冷杉和丽江云杉径向生长的主要气象因子。为进一步评估上年11月气温对川滇冷杉径向生长影响的空间代表性,利用川滇冷杉标准年表分别与CRU TS 4.03(0.5°×0.5°)11月平均气温和最低气温格点气象数据进行空间相关分析。1959—2008年,横断山脉川滇冷杉标准年表与11月平均气温和最低气温显著正相关,能较大范围地反映上年11月最低气温变化,且显著相关关系主要围绕青藏高原东南部及附近地区,但采样地点附近地区相关性最高。综上所述,上年冬季(尤其是11月)气温对横断山脉中部地区川滇冷杉径向生长有明显促进作用。
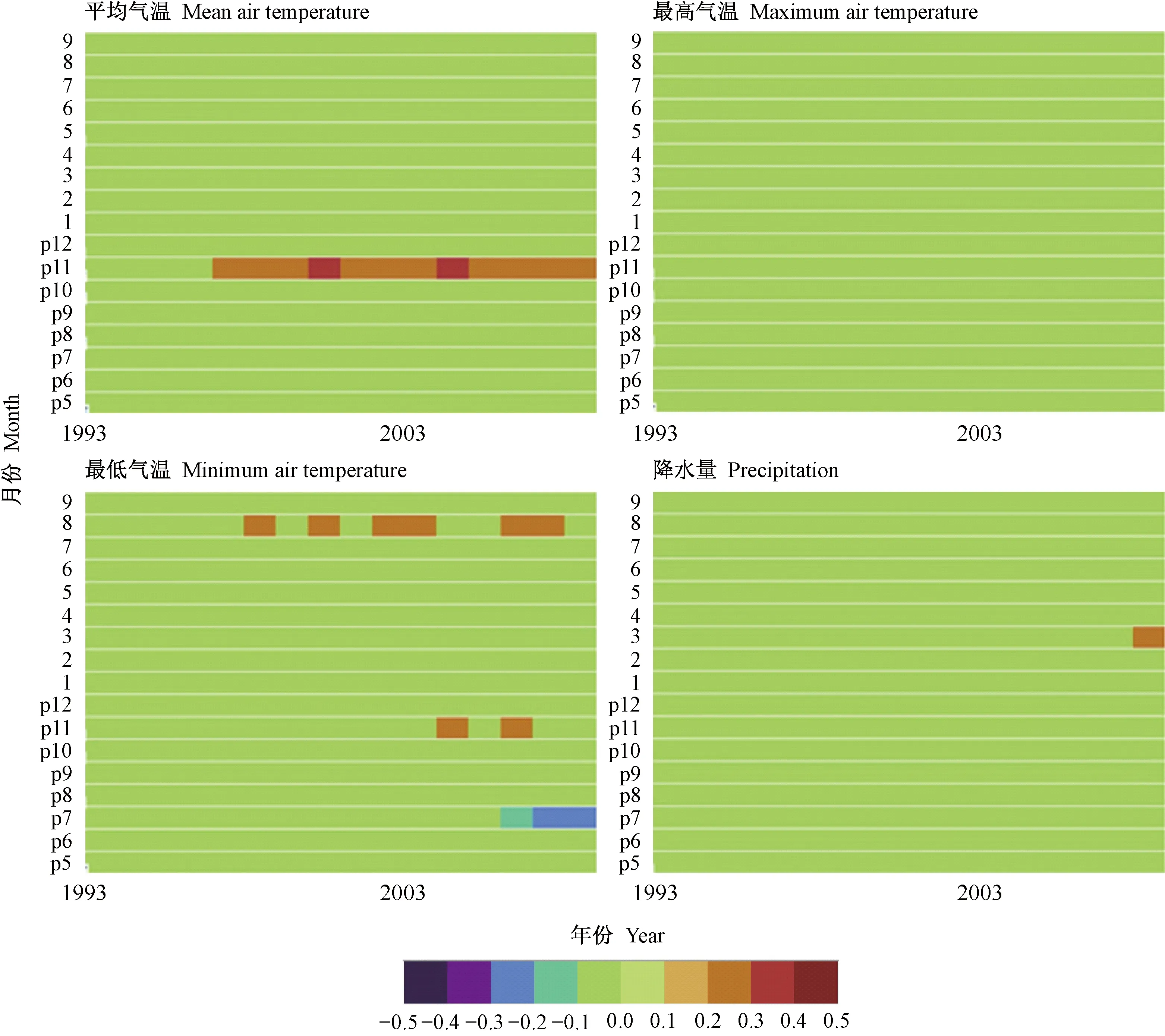
图7 丽江云杉与气象因子的滑动响应关系分析Fig.7 Moving response analysis of P. likiangensis with climate factors
4 讨论
4.1 2个树种径向生长与气象因子的关系
川滇冷杉和丽江云杉径向生长与气象因子的响应关系既存在共性又表现出一定差异性,这与Fan等(2009)的研究结果一致,可能是由于物种的生理学特性不同所致(Gaoetal., 2017),这与高露双等(2011)在长白山、于健等(2017)在小兴安岭和Guo等(2018)在青藏高原东缘等地区的研究结论一致。据报道,鉴于成排木质部母细胞的大小和数量受气候、地形、年龄、活力和竞争等因素影响,径向生长对气候变化的响应会表现出很大的环境或树种差异(Fontietal., 2012)。在本研究地点,川滇冷杉和丽江云杉径向生长主要受气温的限制,而不是降水量,这与前人在横断山脉中部地区的研究结果(Fanetal., 2008; 2009)一致。然而,Bi等(2015)研究发现,玉龙雪山丽江云杉主要受气温和降水量的共同影响,并基于标准年表重建了过去361年春季帕尔默干旱指数(PDSI)变化。尽管上述2个研究海拔高度非常接近(3 240~3 340 m),但为什么会表现不同研究结论?Bunn等(2018)在加利福尼亚沙斯塔山高山树线研究指出,季节平均气温低于7.5 ℃的树木主要受气温驱动,而季节平均气温高于7.5 ℃的树木主要受降雨量限制。树木径向生长与气候变化的关系是树木生理特征与环境条件长期适应的结果(吴祥定, 1990a)。在山区,高海拔树木径向生长通常受气温控制,而低海拔径向生长速率通常受降水量限制(Fanetal., 2009)。高海拔地区树木径向生长受生长季气温限制的相似研究结论在昆仑山(Huangetal., 2019a)、长白山(于健等, 2016)和印度东北部(Yadavaetal., 2015)等地均有报道。横断山脉地区森林上限生长季相对较短(7—8月),8月树木刚好处于生长最旺盛的时期,树木的各项生理活动进入盛期,此时较高的气温利于增加光合作用,提供充足的光合产物供树木生长,进而延长生长季,一定程度地利于树木径向生长,较易形成宽轮。相反,较低的夏季气温可能会导致树木生长提前停止,光合作用减弱,细胞分裂速度减慢,较易形成窄轮(Fritts, 1976)。
树木生长不仅受生长季气象因子的影响,生长季节前的秋季、冬季和春季的气象条件也可能影响树木生长(Fritts, 1976)。本研究发现,冬季气温也对川滇冷杉和丽江云杉的生长起着至关重要的作用,这与青藏高原东北部(Gouetal., 2007)、东部(Zhangetal., 2015)和东南部(Huangetal., 2019b)的研究结果一致。因此,推断高海拔地区树木生长对冬季气温的积极响应似乎是青藏高原山区的普遍现象(Fanetal., 2009)。不仅如此,世界上其他潮湿山区也记录了类似反应模式,如中国的东北部(Zhuetal., 2009)、东南部(Shietal., 2010)、中部(Zengetal., 2019)、西北部(Gaoetal., 2013)、西南部(Fangetal., 2018)以及印度喜玛拉雅西部(Borgaonkaretal., 2009)和美国哈德逊河谷(Pedersonetal., 2004)等。迄今为止,针对冬季低温对随后树木径向生长的影响有如下几种解释: 高海拔地区树木可能因寒冷的冬季气温而引发芽损伤、霜冻干燥和细根死亡,从而限制下一个生长季节的树木生长(Gouetal., 2007)。此外,较低的冬季气温可能还会造成较厚的冻土层,推迟融雪时间和春季开始日期,缩短树木生长季,对树木生长产生不利影响(Fritts, 1976; Fanetal., 2008)。相反,温暖的冬季可使树木合成非结构性碳水化合物和其他有机物质,在生长季节促进木材的早期生长(Gouetal., 2007)。此外,本研究还观察到川滇冷杉和丽江云杉的生长与当年3月降雨量存在显著正相关,这与该地区的前人研究结论(Fanetal., 2008; Chenetal., 2018)相符。低温也许不是树木生长和高山树线动态变化的唯一驱动力,但树木生长对水分表现出了共同敏感性(Lyuetal., 2019)。冬季降水可增加土壤湿度,从而增加树木生长初期的有效湿度,减少生长季前期由于气温过高导致的水分不足,产生宽轮。换句话说,冬季的水分可利用性在树木径向生长中起着关键作用。
4.2 空间大尺度气候变化对2个树种的影响
近年来,在青藏高原东南部高海拔地区开展的很多古气候重建研究均捕捉到了最近史无前例的气候变暖事件(Liangetal., 2009; Zhuetal., 2011; Lietal., 2017)。然而,一项在西藏东南部基于树木年轮密度的研究没有跟踪到这种变暖趋势(Bräuningetal., 2004)。在昌都地区,仪器测定的夏季气温和树木年轮数据都没有显示最近十年的变暖趋势(Briffaetal., 2001; Bräuningetal., 2004)。在青藏高原东部地区,过去250年夏季末气温重建同样没有显示变暖趋势(Xingetal., 2014)。“分离现象”已在大量研究中得到证实,气候变暖降低了北半球高纬度地区近期树木生长对气温的敏感性,并影响树木生存(D’Arrigoetal., 2008)。树木生长的显著下降可能造成或伴随着树木死亡和森林死亡率的显著增加(Williamsetal., 2013)。然而,与广泛报道的北半球高纬度地区出现响应“分离现象”不同(Briffaetal., 1998; D’Arrigoetal., 2008),在本研究区域,高海拔地区树木生长对最近气候变暖的敏感性并没有明显丢失,这与Zhang等(2010)在青藏高原东北部的研究结果一致。类似的研究结论在长白山(Caoetal., 2018)、大别山(Shietal., 2013)、亚洲戈壁沙漠南缘(Chenetal., 2014)和加拿大落基山脉北部(Floweretal., 2011)等地区也被证实,这可能是在上述地区缺乏某些与“分离现象”相联系的因素有关,如干旱胁迫和融雪延迟(D’Arrigoetal., 2008)。因此,未来有必要从世界各地的更多地点收集关于气候变暖对树木生长影响的全面证据。
不仅如此,大量研究发现局地气候影响不能单独解释该地区树木径向生长对气候变化的响应,一些大尺度大气-海洋-陆地系统变化也可能通过影响该地区局地气候间接影响树木生长(Zhangetal., 2014; Shietal., 2017)。Li等(2011)研究发现,厄尔尼诺-南方涛动(ENSO)、太平洋年代际振荡(PDO)和太阳辐射可以影响横断山脉中部地区夏季气温的冷暖变化,进而对该区域树木生长产生影响。Liang等(2016)利用丽江云杉树木年轮最大晚材密度重建了青藏高原东南部多年8月平均最低气温变化,发现该地区夏季气温变化与北大西洋多年代际振荡(AMO)存在明显的同向关系。Shi等(2017)研究指出,AMO也许是青藏高原东南部冬季平均气温变化的关键驱动力。Cheng等(2019)研究表明,自20世纪70年代以来,青藏高原南部树木生长对ENSO的敏感性增加,这可能是由全球气候变暖导致的。此外,上述研究结果不仅在小兴安岭(于健等, 2017)、秦岭(于健等, 2018)、中国东南部(Wangetal., 2018)和丝绸之路东部(Sunetal., 2019)等地区有类似发现,而且在哈萨克斯坦(Zubairovetal., 2019)和加拿大(Pitmanetal., 2013)的一些地区亦有类似研究结果。因此,在未来气候变暖情况下应将大规模大气-海洋-陆地气候反馈纳入区域森林经营管理战略。
5 结论
横断山脉中部地区川滇冷杉和丽江云杉树木径向生长与气象因子的响应关系表现出一定差异性。相比较而言,川滇冷杉径向生长对气象因子的响应较丽江云杉更敏感,更适用于树轮气候学研究。气温是川滇冷杉和丽江云杉径向生长的主要制约因子,且川滇冷杉对气温的响应强于丽江云杉,主要表现为与上年11月和当年8月平均气温显著正相关,与上年11月和当年4月最高气温显著正相关,与上年5月最低气温显著正相关,与当年3月降水量显著正相关。空间相关分析表明,川滇冷杉标准年表能在较大空间范围反映上年11月最低气温变化,且二者在研究地区附近相关性最高。未来全球气候变暖将有利于上述2个树种的树木生长。本研究结果可为进一步重建我国西南地区过去气候变化历史提供一定的基础数据,同时也为全球气候变化背景下川滇冷杉和丽江云杉的分布预测和保护研究提供理论参考。
猜你喜欢
世界建筑(2022年10期)2022-10-25
生物化工(2021年3期)2021-07-10
景德镇陶瓷(2021年1期)2021-03-24
现代农村科技(2020年7期)2020-12-19
艺术品鉴(2019年12期)2020-01-18
花卉(2020年6期)2020-01-07
花火A(2019年1期)2019-03-22
乡村科技(2019年14期)2019-02-21
启迪与智慧·教育版(2018年12期)2018-02-26
现代养生·上半月(2017年7期)2017-07-07
