
西方蜜蜂的放蜂密度对东方蜜蜂采集行为的影响*
2021-01-22 09:23:22任晓晓孟柏达赵文正龚雪阳陈新兰黄永权周丹银
云南农业大学学报(自然科学) 2021年1期
任晓晓 ,孟柏达,赵文正,龚雪阳,陈新兰,黄永权,周丹银,董 坤
(1.云南农业大学 动物科学技术学院 东方蜜蜂研究所,云南高校蜜蜂资源可持续利用工程中心,云南 昆明 650201;2.贵州省农业科学院 蚕业研究所,贵州 贵阳 550006;3.云南省罗平县蜂业基地项目中心,云南 罗平 655800)
蜜蜂作为一类重要的传粉昆虫,对于保护植物群落的多样性[1]、维持自然界生态系统的平衡极其重要[2]。目前公认的蜜蜂属的种类有9 种,分别为东方蜜蜂(Apis cerana)、西方蜜蜂(Apis mellifera)、大蜜蜂(Apis dorsata)、小蜜蜂(Apis florea)、黑大蜜蜂(Apis laboriosa)、黑小蜜蜂(Apis andreniformis)、沙巴蜂(Apis koschevnikovi)、绿努蜂(Apis nulunsis)和苏拉威西蜂(Apis nigrocincta)[3]。其中,东方蜜蜂(Apis cerana)与西方蜜蜂(Apis mellifera)因为易于饲养和繁殖得以快速发展,目前已成为成为世界上饲养量最大的2 个蜂种。
中国作为养蜂大国,东、西方蜜蜂的饲养量均位居世界之首。其中,东方蜜蜂是中国分布较广的土著蜂种,因具有采蜜期长、善于利用零星蜜粉源、适应性和抗逆性较强等特点[4-6],对中国的生态系统、农业授粉以及养蜂业均有着不可替代的作用[7-8]。但是,随着西方蜜蜂的引入和快速发展,中国东方蜜蜂的生存受到了一定程度的影响,其原因主要是两者在生存空间和食物资源利用等方面存在生态位重叠[9-10]。此外,东方蜜蜂因存在易躁、易分蜂和易飞逃难以维持强群的特征,且在产蜜量和产卵能力方面远不及西方蜜蜂,因此大多数养蜂者更愿意饲养西方蜜蜂。随着人工饲养西方蜜蜂的种群数量大幅度提升,加之西方蜜蜂大多采用转地放蜂的饲养模式,给东方蜜蜂造成了巨大的生存压力,从而导致其种群数量和分布区域呈下降趋势[11]。因此,为了在发展蜂产业的同时保护中国本土的传粉蜂种,应采取一定的措施来避免东、西方蜜蜂之间的相互竞争,且这一建议在1988 年就已经由中国学者提出[12]。但目前为止,国内外有关该方面的研究还比较少,一些国家(如德国和美国)研究了西方蜜蜂对本地主要传粉蜂种产生的影响[13-14],而国内关于该方面的研究主要集中在西方蜜蜂对东方蜜蜂种间交尾的干扰和遗传多样性的影响等方面[15-17],关于东、西方蜜蜂种间采集行为和食物资源竞争的研究鲜见报道。
云南省罗平县是全国著名的“蜜蜂春繁基地”,每年油菜花期都有大量的西方蜜蜂蜂场从全国各地涌向罗平放蜂,这促使我们开始关注大量西方蜜蜂“空降”罗平后对本地东方蜜蜂的影响。因此,本研究在云南省罗平县的油菜花期,通过调查和观测研究西方蜜蜂放蜂密度,比较西方蜜蜂不同放蜂密度下东方蜜蜂的采集行为(访花频率、单花采集停留时间)及访问蜜源植物种类的变化。试图从食物资源竞争的角度,阐明西方蜜蜂对东方蜜蜂种群影响的生态机制,为合理保护中国本土蜂种,维持植物群落结构及其稳定性提供科学依据。
1 材料与方法
1.1 研究区概况
云南省罗平县地处E103°57 ′~104°43 ′、N24°31′~25°25′之间,位于滇、桂、黔三省交界处,平均海拔1 500 m,年平均降雨量1 743.9 mm,降雨量充沛,属于南亚热带气候和高原季风气候。当地年种植油菜面积达4.6 万hm2,通常在每年的2—3 月达到盛花期,每年大约有5 万余群西方蜜蜂到此地进行春繁。
1.2 试验方法
1.2.1 试验时间和地点
2016 年2 月17—21 日油菜盛花期内,在九龙镇和大水井乡的主要干道上对人工饲养的西方蜜蜂进行走访调查,统计西方蜜蜂的蜂群数量、平均群势和放蜂密度,并根据统计结果在九龙镇和大水井乡的主干道上选择走马田、鲁特、马把、外纳、下阿来和下庄科(以下简称ZMT、LT、MB、WN、XAL 和XZK) 共6 个试验点,在这6 个试验点分别放置供试东方蜜蜂蜂群,以东方蜜蜂放置点周围3 km 的范围为各试验点的试验区域,每个试验区域内均包含2 种生境(农田和山林),其西方蜜蜂放蜂密度见表1。2016 年3 月1—11 日在西方蜜蜂放蜂期对每个试验点农田样方和山林中东方蜜蜂的采集行为进行观测,每个主干道的3 个试验点每次调查需用2 d 时间,所有试验点完成1 次调查需4 d 时间,西方蜜蜂放蜂期内对所有试验点重复观察了2 次。2016 年3 月18—21 日在西方蜜蜂蜂场大规模撤离之后,对所有试验点农田样方和山林中东方蜜蜂的采集行为进行1 次观测。2016 年3 月1—21 日在观测东方蜜蜂采集行为的同时调查试验点周围显花植物种类并采集花粉样本,同时采集东方蜜蜂携带的花粉团。
由表1 可见:在九龙镇方向上,鲁特村周围3 km 范围内的西方蜜蜂蜂群数量最多、放蜂密度最高,其次是走马田村,西方蜜蜂放蜂密度最低的是马把村;在大水井方向上,西方蜜蜂蜂群数量最多、放蜂密度最高的是下阿来村,然后依次是外纳村和下庄科村。在九龙镇和大水井乡的2 条路线上西方蜜蜂的放蜂密度均形成了高、中、低的放蜂密度梯度。
1.2.2 西方蜜蜂放蜂密度对东方蜜蜂采集行为的影响
(1)农田样方中东方蜜蜂的采集行为观测
在距离东方蜜蜂放置点50 m 和80 m 处,各选择2 个2 m×2 m 的样方,观测样方内传粉蜜蜂的采集行为。观察时用摄像机(索尼HDR-XR 5201)记录传粉蜜蜂的访花行为,周围尽量避免人为干扰。晴天观察,观察时间为10:00—18:00,每个样方每隔1 h 观察1 次,每次观察持续时间为20 min。各试验点重复观察3 d。用计数器统计蜜蜂种类和数量,通过整理录像确定东方蜜蜂的访花频率、单花采集停留时间。通过对比,分析不同西方蜜蜂放蜂密度下,农田样方中东方蜜蜂采集行为的变化情况。
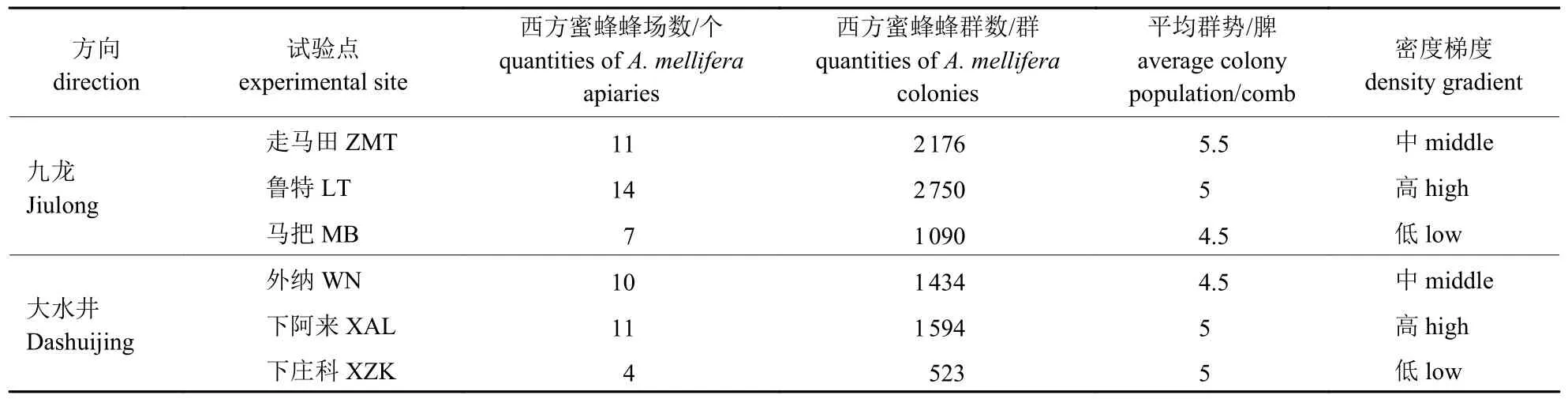
表1 试验点周围西方蜜蜂的放蜂密度Tab.1 The colony densities of A.mellifera around different test sites
(2)山林中东方蜜蜂的采集行为观测
因在山林中受到地形和植被限制不便选择固定样方,所以在东方蜜蜂放置点周围50~80 m 的范围内采取随机走动的观察方式观察山林中东方蜜蜂的采集行为。10:00—18:00 沿山间小道观察并统计视野中东方蜜蜂的数量,记录东方蜜蜂的访花频率和单花停留时间。每隔 1 h 观察1 次,每次观察持续时间20 min,各试验点重复观察3 d。
1.2.3 西方蜜蜂放蜂密度对东方蜜蜂利用蜜粉源植物的影响
(1)试验点周围显花植物调查及对照花粉采集
在6 个试验点周围3 km 范围内的山林和农田中调查显花植物种类并采集油菜及其他野生蜜粉源植物的花朵(采集时要选择还未散粉的花朵,以免有异种花粉污染),置于FAA 固定液(90 mL 70%乙醇+5 mL 甲醇+5 mL 冰乙酸)中保存,带回实验室4 ℃保存制作花粉永久玻片[18]。显花植物调查每隔3 d 进行1 次,采集植物花粉时记录采集时间、地点并编号,同时拍摄野生显花植物样本的照片。
(2)永久玻片制作
本研究采用涂片法制作花粉永久玻片,首先切取适量花粉染剂(琼脂粉5 g+去离子水200 mL+丙三醇200 mL+藏红T 饱和液数滴,微波加热后冷却备用)置于干净的载玻片上,放在酒精灯上烘烤至熔化,稍稍冷却后备用。将花朵从FAA固定液中取出,使用双蒸水冲洗干净,切下花药放置于花粉染剂中并用镊子将花药夹碎,使花粉散落出来,轻轻盖上盖玻片,避免产生气泡。花粉玻片制作完毕后,标注编号、植物种类、采集时间及地点,放置于高倍显微镜下调好视野后拍照[19-20]。
(3)东方蜜蜂花粉团采集
对供试东方蜜蜂蜂群进行花粉团采集,采集时在每群东方蜜蜂巢门口各自安装1 个脱粉器,脱粉器下放置1 个干净的浅盘。每天18:00 收集浅盘上的花粉团,标明采集时间及地点,带回实验室4 ℃保存备用。每隔2 d 安装1 次脱粉器,每群东方蜜蜂各收集3 次。
(4)东方蜜蜂花粉团的花粉种类及数量比例分析
对每种颜色的花粉团随机选择10 粒(不够10 粒的都取),分别置于离心管中,加入1 mL 30 ℃的双蒸水溶解,之后3 000 r/min 离心10 min,倒弃上清;向离心管中加入1 mL 冰乙酸,摇匀后静置2 h,然后再3 000 r/min 离心10 min,倒弃上清;向离心管中加入新配置的乙酰解的混合液[V(乙酸酐)∶V(浓硫酸)=9∶1] 1 mL,85 ℃水浴8 min,取出后冷却备用[21];取1 个干净的载玻片,滴上适量样液,小心盖上盖玻片,使用带CCD 照相机的“Zeiss”生物显微镜在400 倍镜下观测并拍照,每张玻片观察10 个视野;请植物分类工作者根据植物照片及花粉玻片照对显花植物种类进行鉴定,根据鉴定结果和制备好的对照花粉玻片鉴别花粉种类,记录每个视野中各种花粉的数量,得出每种花粉所占百分比;取1 个血球计数板,在中央计数室盖上盖玻片,取10 μL样液置于盖玻片的边缘,使样液慢慢渗入,静置5 min 后在显微镜下观察计数,每个样本计数3 次,统计每个花粉团所含花粉的总数量,分析西方蜜蜂不同放蜂密度对东方蜜蜂利用蜜粉源植物的影响。
1.3 数据处理
运用SAS 软件及Microsoft Office Excel 2007进行数据分析和作图。
2 结果与分析
2.1 西方蜜蜂不同放蜂密度对东方蜜蜂采集行为的影响
2.1.1 西方蜜蜂大规模撤走前、后对东方蜜蜂在2 种生境中分布比例的影响
由图1 所示:油菜盛花期(3 月1—11 日)西方蜜蜂蜂场还没有大规模撤离时,山林中东方蜜蜂的比例均大于农田中东方蜜蜂的比例;油菜末花期(3 月18—21 日) 西方蜜蜂蜂场大部分撤离后,在田间样方中观测到的东方蜜蜂所占的比例相比于油菜盛花期有明显增加,山林中观测到的东方蜜蜂所占比例有明显的降低。

图1 东方蜜蜂在2 种生境中所占数量比例随时间(西方蜜蜂蜂场撤离)的变化图Fig.1 The quantitative proportion of A.cerana varied over time in two ecological types
2.1.2 西方蜜蜂不同放蜂密度对农田样方中东方蜜蜂的访花频率和单花采集停留时间的影响
由图2 所示:Pearson 相关系数r=0.971 1,所对应的P=0.001 2<0.05,说明农田样方中东方蜜蜂的平均访花频率与其周围3 km 范围内的西方蜜蜂数量呈显著的正相关关系;Pearson 相关系数r=−0.959 3,所对应的P=0.002 5<0.05,说明农田样方中东方蜜蜂的单花采集停留时间与其周围3 km 范围内的西方蜜蜂数量呈显著的负相关关系。即周围3 km 范围内的西方蜜蜂放蜂密度越大,对应的东方蜜蜂在农田中的平均访花频率越大,单花采集停留时间越短。
2.1.3 西方蜜蜂不同放蜂密度对山林中东方蜜蜂访花频率和单花采集停留时间的影响
由图3 所示:在九龙方向上不同西方蜜蜂放蜂密度下山林中东方蜜蜂的访花频率和单花采集停留时间均存在显著差异(P<0.05),且东方蜜蜂的访花频率大小顺序为:鲁特>走马田>马把,东方蜜蜂的单花采集停留时间大小顺序为:马把>走马田>鲁特;在大水井方向上下阿来和外纳试验点山林中东方蜜蜂的访花频率和单花采集停留时间差异不显著(P>0.05),但是下阿来和外纳试验点山林中东方蜜蜂的访花频率和单花采集停留时间分别与下庄科试验点相比差异均显著(P<0.05)。
2.2 西方蜜蜂不同放蜂密度对东方蜜蜂采集粉源植物情况的影响
2.2.1 西方蜜蜂不同放蜂密度对东方蜜蜂采集花粉总量的影响
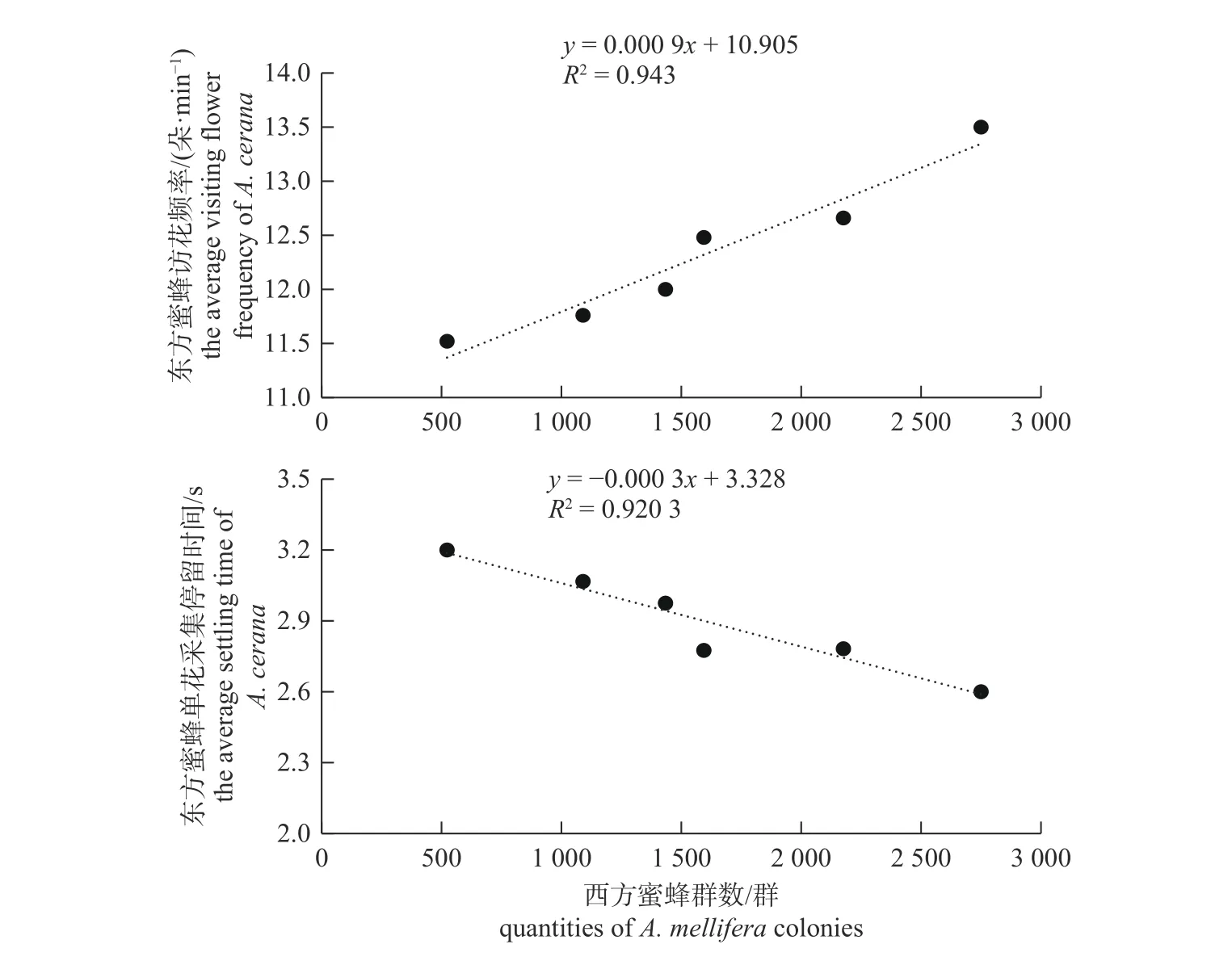
图2 农田样方中东方蜜蜂平均访花频率和单花采集停留时间与西方蜜蜂数量的相关性分析图Fig.2 The correlation analysis between the average visiting frequency and the average settling time of A.cerana in farmland quadrats with the quantity of A.mellifera
利用脱粉器对6 个试验点供试东方蜜蜂的花粉团进行了采集,经观察,东方蜜蜂脱下的花粉团有黄色、橘色、白色、粉色和褐色5 种。对各群东方蜜蜂的花粉团进行数量分析时,为避免东方蜜蜂的群势不同对试验结果产生影响,将花粉团总数量和不同颜色花粉团数量除以每群的脾数后再进行分析。由图4 所示:东方蜜蜂花粉团总数量与其周围3 km 范围内的西方蜜蜂数量呈负相关关系(r=−0.952 3,P=0.003 4<0.05),即周围3 km 范围内西方蜜蜂的放蜂密度越大,东方蜜蜂采集到的花粉团数量越少。这说明西方蜜蜂放蜂给东方蜜蜂带来了较大的竞争压力,西方蜜蜂不仅在农田中访问油菜,也会对山林中的野生蜜粉源进行采集,且西方蜜蜂的放蜂密度越大,对东方蜜蜂的竞争压力越大,东方蜜蜂得到的食物报酬越少。
2.2.2 西方蜜蜂不同放蜂密度下东方蜜蜂采集的花粉种类
油菜花期,在各试验点供试东方蜜蜂蜂群周围3 km 范围内共采集到了59 种显花植物,隶属于22 目29 科57 属。每个试验点东方蜜蜂采集植物花粉统计结果如表2 所示。由表2 可知:在九龙方向上,西方蜜蜂放蜂密度最高的鲁特试验点,东方蜜蜂采集油菜花粉的比例最低(8%),但采集植物花粉的种类最多(10 种),在西方蜜蜂放蜂密度最低的马把试验点东方蜜蜂采集油菜花粉的比例最高(21%),但采集植物花粉的种类最少(4 种);在大水井方向上,也得到了相似结果,即随着西方蜜蜂放蜂密度的增加,东方蜜蜂采集油菜花粉的比例降低,但采集的粉源植物种类增加。
2.2.3 西方蜜蜂不同放蜂密度对东方蜜蜂采集零星蜜粉源植物花粉比例的影响
由图5 所示:Pearson 相关系数r=0.852 39,所对应的P=0.031 1<0.05,说明东方蜜蜂采食零星粉源植物花粉的比例与其周围3 km范围内的西方蜜蜂数量存在显著的正相关关系,即西方蜜蜂的放蜂密度增大时,对应的东方蜜蜂采食零星粉源植物花粉的比例就越大(这里将油菜以外的粉源植物称为零星蜜粉源)。
3 讨论
本研究首先关于相同试验点西方蜜蜂放蜂密度随着时间推移发生的变化对东方蜜蜂在农田和山林中的数量比例产生的影响进行了研究。结果显示:在油菜盛花期农田中东方蜜蜂的数量比例普遍低于山林区域,但是在油菜末花期,随着西方蜜蜂蜂场的大量转移,农田中访花的西方蜜蜂数量骤减,东方蜜蜂在农田中采集所面对的竞争压力也随之减小,因此之前被排挤到山林中采集野生蜜粉源的部分东方蜜蜂又回到农田中采集油菜蜜粉源,最终出现农田中东方蜜蜂的数量比例提高、山林中东方蜜蜂的数量比例降低的现象。这表明西方蜜蜂数量多,放蜂密度大时,东、西方蜜蜂之间确实存在激烈的食物竞争,而且西方蜜蜂由于数量多、个体大,并善于利用农田中油菜这一大宗蜜源,在竞争过程中具有明显的优势,迫使在农田中采集的东方蜜蜂不得不到山林中采集野生蜜粉源。

图3 西方蜜蜂不同放蜂密度下山林中东方蜜蜂的访花频率和单花采集停留时间Fig.3 The average visiting flower frequency and the average settling time of A.cerana in the mountain forest under different colony densities of A.mellifera
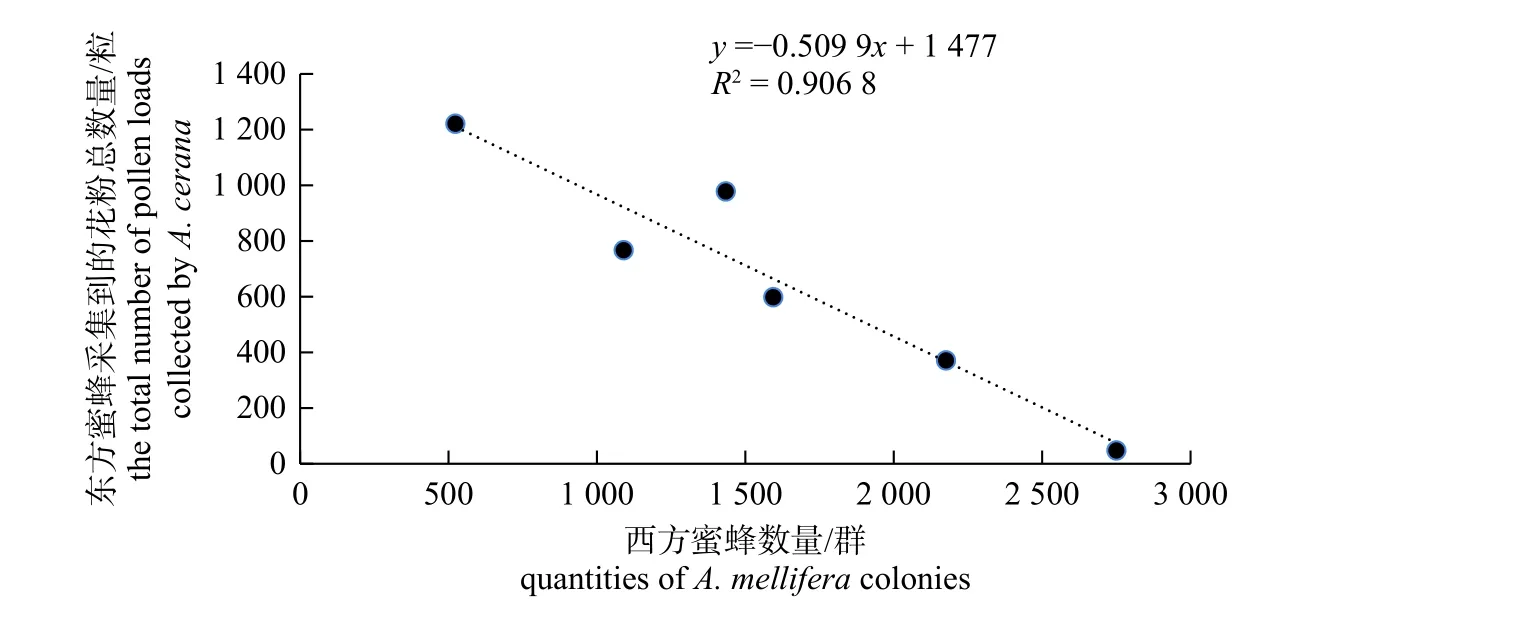
图4 东方蜜蜂花粉团总数量与西方蜜蜂放蜂数量相关性分析图Fig.4 Correlation analysis between the the total number of pollen loads collected by A.cerana with the quantity of A.mellifera
西方蜜蜂放蜂密度在空间上也会存在差异,这导致东方蜜蜂在不同区域采集时受到西方蜜蜂干扰的程度也不同,其采集行为(采集行为指标通常为访花频率和单花采集停留时间)会发生不同程度的变化[22]。本研究中,在农田生境下,油菜盛花期随着西方蜜蜂蜂群数量的增加,东方蜜蜂的访花频率随之提高,单花采集时间随之缩短。其原因是西方蜜蜂的密度越大,对花蜜的采食量越大,从而导致同一区域内东方蜜蜂在每一朵花上能够采集到的花蜜量减少,东方蜜蜂的单花采集时间缩短,访花频率增加。而在山林生境中,九龙方向上的3 个试验点东方蜜蜂的访花频率和单花采集停留时间均存在显著差异,这表明西方蜜蜂对东方蜜蜂采集行为产生影响的情况已经扩散到了山林区域,而且所产生影响的程度会随西方蜜蜂放蜂密度的不同而发生变化;在大水井方向上,由于外纳和下阿来试验点附近西方蜜蜂蜂群数量比较接近,二者山林中东方蜜蜂的访花频率和单花采集停留时间没有出现显著差异,但二者与低密度的下庄科试验点相比,东方蜜蜂采集行为指标均发生了显著变化。

表2 各试验点东方蜜蜂采集的植物花粉的种类及比例Tab.2 Species and proportion of plant pollen collected by A.cerana in experiment sites %
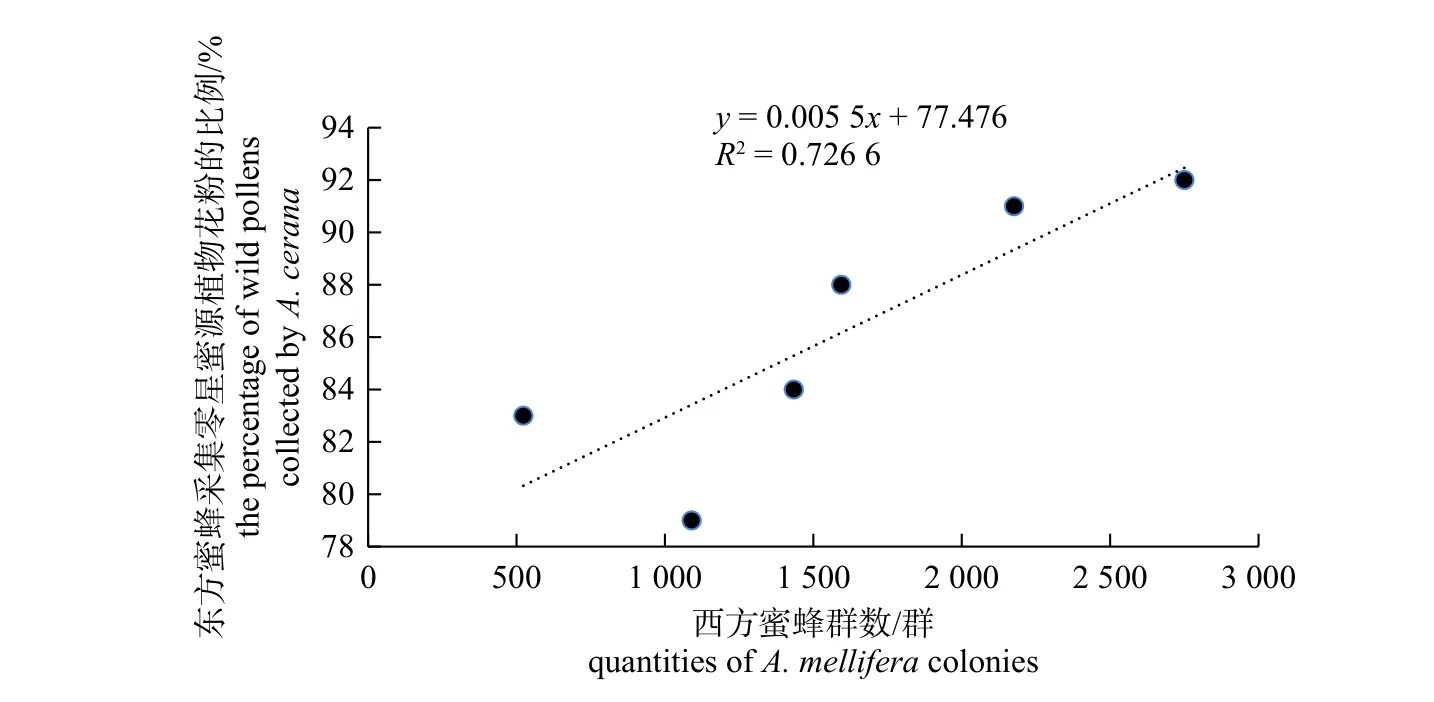
图5 东方蜜蜂采食零星源植物花粉的比例与西方蜜蜂放蜂密度的相关性分析Fig.5 Correlation analysis between the percentage of wild pollens collected by A.cerana with the colony densities of A.mellifera
西方蜜蜂对处于平缓地势的大宗蜜源具有较强的采集能力,而东方蜜蜂能够发现并善于利用分散的零星蜜源[23]。本研究中东方蜜蜂在不同西方蜜蜂放蜂密度的影响下对大宗蜜粉源和零星蜜粉源的利用程度有所不同,在西方蜜蜂放蜂密度大的区域内东方蜜蜂对油菜大宗蜜粉源的利用率降低,对零星蜜源的利用率提高,且各群东方蜜蜂采集到的花粉团总数量与其周围3 km 范围内的西方蜜蜂数量呈负相关关系,各群东方蜜蜂采食野生粉源花粉的比例与其周围3 km 范围内西方蜜蜂蜂群数量呈正相关关系。这表明在西方蜜蜂的竞争压力下,东方蜜蜂在食物资源利用方面的生态位会发生偏离并逐渐朝着以零星蜜粉源为主要食物来源的方向发展。
根据本试验的研究结果,我们推测东方蜜蜂并非不喜欢大宗蜜源,而是迫于西方蜜蜂的竞争压力,转而采集回报较少的野生蜜粉源。在国外也有类似的研究结果,如FAYEZ 等[14]对意大利蜜蜂(西方蜜蜂亚种)与德国本地野生传粉蜂类的种间竞争进行的研究,结果显示因两蜂种之间绝大部分生态位重叠,食物竞争激烈,意大利蜜蜂迫使本地野生传粉蜂类改变其原有的采集偏好;美国的一项研究也证明:西方蜜蜂能够凭借其种群数量优势迫使美国本地传粉蜂类(Halictus ligatus、H.confuses和Augochlorella striata)访问回报较少、质量较差的蜜粉源植物。本试验中,东方蜜蜂的采集行为和对食物资源的利用受到了西方蜜蜂的影响,但是,在这一情况下东方蜜蜂种群的种群繁殖力是否会受到影响还有待进一步研究。
4 结论
西方蜜蜂放蜂密度对东方蜜蜂的采集行为和粉源植物利用有显著影响,西方蜜蜂放蜂密度越大,种间食物竞争越激烈,东方蜜蜂的采集区域由大宗蜜源转向零星蜜源的趋势越明显,最终获得的食物报酬也越少。本研究结果为今后进一步制定西方蜜蜂大规模放蜂区域内东方蜜蜂的保护措施提供了基础资料。
猜你喜欢
河南农业(2024年3期)2024-03-05 02:20:52
中国蜂业(2023年2期)2023-04-04 01:11:38
经济林研究(2022年2期)2022-07-20 09:40:24
作文周刊·小学一年级版(2022年4期)2022-03-07 01:13:49
中国蜂业(2021年5期)2021-05-22 02:59:26
中国蜂业(2021年10期)2021-03-02 01:17:02
爱你(2019年17期)2019-11-14 11:56:19
爱你·健康读本(2019年5期)2019-06-06 04:00:06
现代农业(2018年7期)2018-08-01 12:27:44
中国蜂业(2017年8期)2017-09-05 12:42:34
