
杂交水稻表型、杂种优势和配合力的关系
2021-01-21 09:18陈俊孝游艾青
湖北农业科学 2020年24期
陈俊孝,游艾青,刘 凯
(湖北省农业科学院粮食作物研究所∕粮食作物种质创新与遗传改良湖北省重点实验室,武汉 430064)
杂种优势是自然界的一种普遍现象,优良亲本的选育是获得强优势杂种品种的基础,但是实际生产中亲本的优良性状不能很好地传递给后代,没有表型优势。具有优良性状的亲本并不总是将这些性状传递给它们的杂交后代,杂种优势水平高的杂交组合不一定具有高产表型[1]。为了有效评估亲本的生产性能,育种家们提出了配合力的概念,并有效应用于植物杂交育种中亲本和组合的选择,促进了杂交育种的进程。配合力高低是选择杂交种亲本最重要的依据,它直接影响杂交种的综合性能。
配合力分为一般配合力(General combining abil⁃ity,GCA)和特殊配合力(Specific combining ability,SCA)[2,3]。通过双列杂交或不完全双列杂交等交叉设计试验,育种家可以轻松识别出具有高GCA 的亲本和高SCA 的杂交组合。大多数的研究以鉴定优良亲本为主要目的,对配合力遗传基础的分子研究仍有很多不足[4-6]。
本研究构建了一套不完全双列杂交群体,利用数字化考种机并结合人工考察对群体的产量相关性状进行了考察,分析了产量相关性状的表型、亲本的一般配合力、F1的特殊配合力和F1的杂种优势之间的相关性,探讨了GCA 与SCA 对产量相关性状及杂种优势的不同贡献,分析了不同参数在育种中的选择效率并为杂交育种策略提出了见解。
1 材料与方法
1.1 NCII群体的构建与种植
以收集的来自世界范围内的具有多样性的栽培水稻为原材料,筛选生育期适中、株型紧凑、株高适中的126 个株系作为父本材料,分别与H2613S、H6S、C815S 和HD9802S 4 个优质光温敏两系不育系进行不完全双列杂交,构建1 个NCII 群体。剔除获得杂交种不足的亲本和杂交组合材料,最终构建了1 个包含96 个父本与4 个不育系母本的不完全双列杂交群体。利用Illumina HiSeq2000 测序仪对96 个父本进行了低丰度重测序,对4 个母本进行了覆盖基因组12 倍的深度重测序。2013年夏天,将材料以随机顺序排列种植于湖北省鄂州市并安排2次重复。所有的试验材料每个家系种2 排,每排12株,行间距25.5 cm,株间距15.5 cm,正常田间管理,每个家系的中间6 株以单株收种,用作后续的表型性状考察。
1.2 NCII群体表型鉴定
抽穗期的测量:每隔2 d 在田间进行观察,记载小区50% 的单株主茎穗见穗日期减去播种日期换算为抽穗期。株高的测量:待植株成熟后用尺子量取植株最高穗子的颖尖到地面的高度作为株高。产量性状的考察:植株成熟后,将地上部分进行单株收种,种子收获后室温放置1~3 个月,以控制各试验材料间的种子含水量相对一致。所有的完整主穗均被用来检测单株穗数,单株每穗实粒数超过5 粒的穗子被认为是有效穗数。随机选取5 根有效穗考察穗长,量取穗结节部位到最顶端颖壳的长度作为穗长。每个单株的所有谷粒全部脱粒后用于每穗粒数、结实率、粒长、粒宽、长宽比、千粒重和单株产量等产量性状的考察。这些产量性状的考察是使用数字化考种机(Yield Trait Scorer)来完成的[7]。每个家系的平均值作为最终值进行分析。
1.3 NCII群体配合力分析
NCII 设计是两因素的方差设计,父本、母本、父本×母本和环境的效应可以用表1 列出的方差分析来计算。

表1 NCII群体中各参数的计算
根据表1 可以计算出父本的加性遗传方差(δ2m)、母本的加性遗传方差(δ2f)、F1的非加性遗传方差(δ2m)f、F1的遗传方差(δ2G)、环境方差(δ2w)、F1的表型方差(δ2P)、狭义遗传力(Hn)和广义遗传力(Hb)。相关参数的计算公式如下:
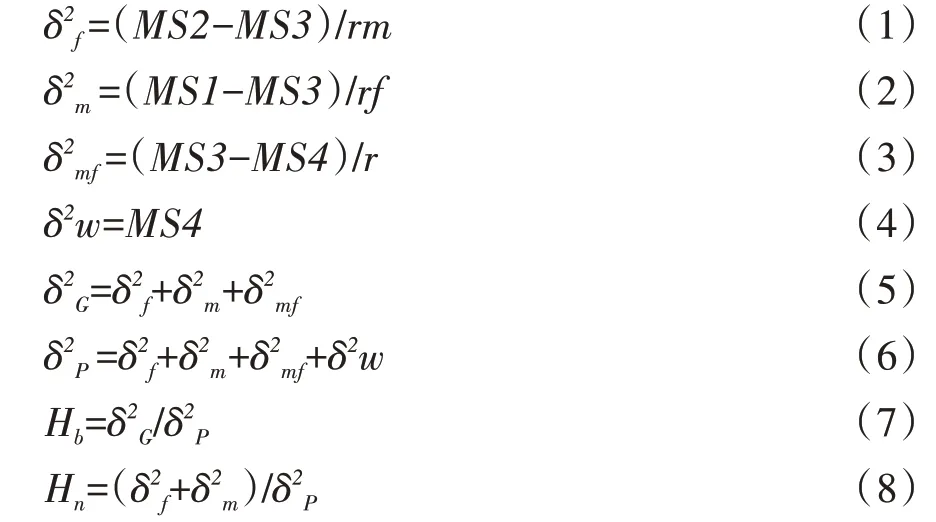
每个性状的表型和配合力之间的关系可以用下面的数学公式表现:

式中,Yij表示每个F1的表型值;Y表示所有F1的表型的平均值;Gi表示父本的一般配合力;Gj表示母本的一般配合力;Sij表示F1的特殊配合力。
一般配合力和特殊配合力的计算采用以下公式:

2 结果与分析
2.1 NCII群体遗传结构与表型相关性分析
利用全基因组的SNPs 信息分别构建了亲本和NCII 群体的进化树,群体结构能够很明显地划分为5 个组别。每个后代家族通过拥有共同的父本或母本血统而有50% 相同的遗传背景。利用fast⁃STRUCTURE 软件对NCII 群体的基因型数据进行了群体结构分析。主成分分析结果(图1a-d)表明,前6 个主成分可以将群体明显地划分为5 个亚群,并能解释约76.6%的遗传变异。
表2、表3 为NCII 群体之间各产量相关性状的表型变异和相关系数。单株产量和株高、穗长、粒宽、单株有效穗数、每穗粒数、结实率和千粒重等性状呈显著或极显著正相关,与粒长、长宽比和抽穗期均呈负相关。千粒重与粒型的相关性较高,而与其他表型的相关性较低。
2.2 GCA 与SCA 对性状及杂种优势的不同贡献

图1 NCII群体遗传结构

表2 NCII群体之间农艺性状的表型变异
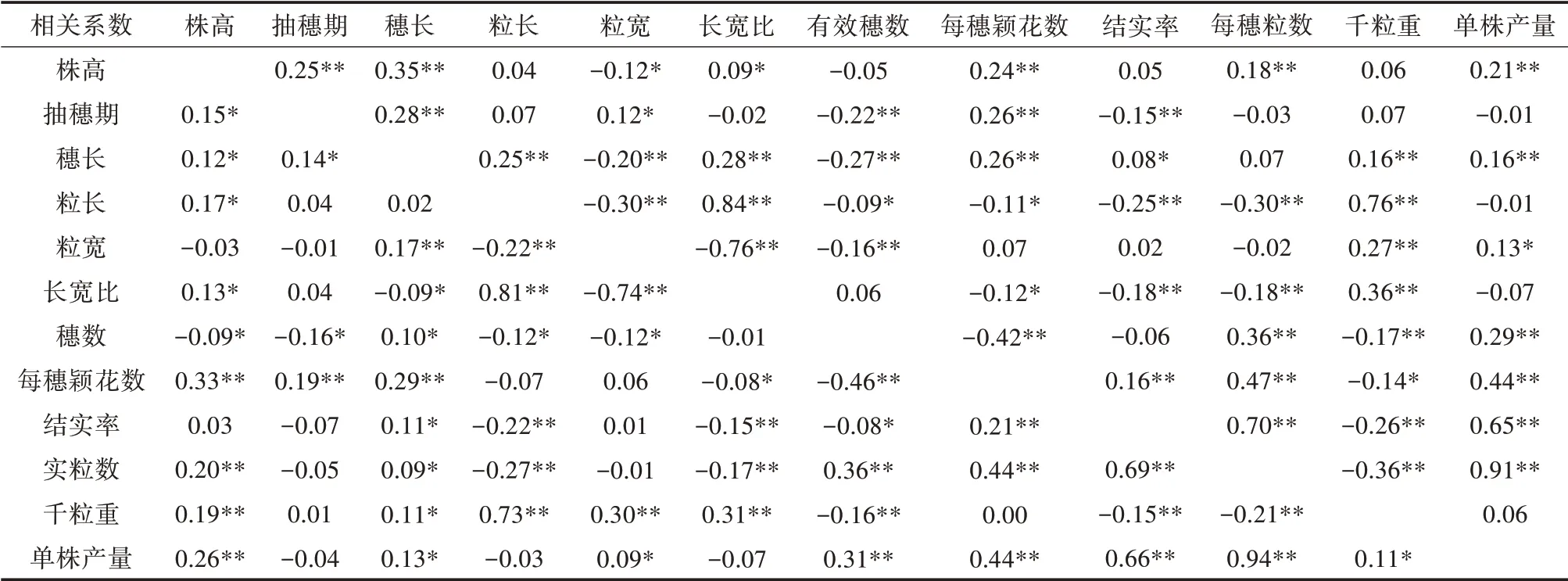
表3 NCII群体之间农艺性状的遗传系数和表型相关系数
利用双因素方差分析了父本、母本、杂交种以及环境因素之间对产量相关性状的效应,发现父本、母本及杂交材料在所有的性状中均有显著差异。进一步计算所有产量相关性状的父本加性效应(δ2m)、母本加性效应(δ2f)、杂交材料的非加性效应(δ2mf)、环境变异(δ2e)、狭义遗传力(Hn)和广义遗传力(Hb)。在所有的性状中,子粒长宽比表现出最高的遗传力,而单株产量表现出最低的遗传力。如果性状具有较高的狭义遗传力,表明它们更多的是受加性效应决定的。相反的,若性状具的狭义遗传力很低且与广义遗传力差值较大,则表明它们更多的是受非加性效应影响。
为了阐明各产量相关性状间的表型、配合力和杂种优势之间的关系,对每个性状估算了双亲的GCA、F1的SCA 和杂种优势效应(HA)。此外,引入了亲本的遗传距离(PGD)和杂合度(OH)2 种参数进行分析。所有性状的表型值,亲本的一般配合力、杂交材料的特殊配合力和杂种优势间有高度的相关性。杂种优势与亲本间的遗传距离和杂合度之间没有显著的相关性(图2)。

图2 12 个水稻产量相关性状的表型、杂种优势和配合力之间的相关性分析
以杂交种的单株产量为例(表4、表5),特殊配合力与表型之间的相关性最高,相关系数为0.75;特殊配合力与杂种优势之间也有较高的相关性,相关系数为0.71。一般配合力与单株产量间也表现出较高的相关性,相关系数为0.61;但是一般配合力与杂种优势间的相关系数为0.21。结果表明,非加性效应对杂种优势的产生具有更大的贡献,加性效应和非加性效应对产量本身均有重要的作用。这些参数对不同农艺性状的表型和杂种优势具有不同的贡献,具有较高遗传力的性状比如粒长,一般配合力的贡献比特殊配合力更高;每个性状的平均杂种优势与广义遗传力和狭义遗传力均表现为负相关(图3),其中杂种优势与狭义遗传力的相关系数为0.71,杂种优势与广义遗传力的相关系数为0.80。
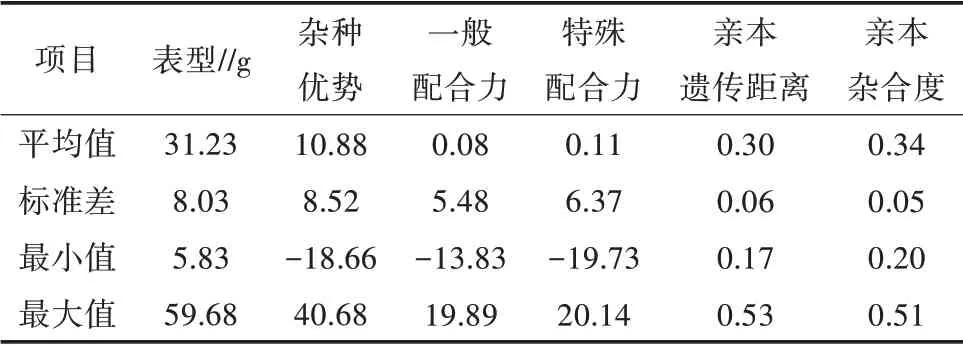
表4 NCII群体的产量性状表型参数

表5 NCII群体的产量性状表型参数的相关性
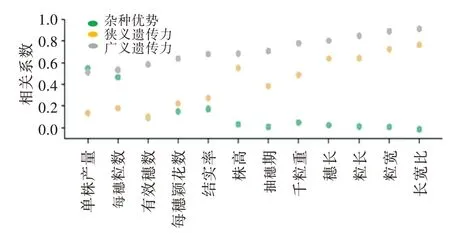
图3 水稻各产量性状的平均杂种优势与狭义遗传力和广义遗传力之间的关系
2.3 不同参数在育种中的选择效率
在实际育种中,通常会以亲本的表型为主要参考标准培育优良的杂交水稻,随着对分子遗传机理的不断研究,亲本的一般配合力和杂交材料的特殊配合力及杂种优势也作为参考标准应用于遗传育种当中。为了进一步探索不同参数间的选择效率,从NCII 群体中划分了4 个不同的F1群体,每个群体包含40 个家系。4 个群体分别是:①具有最高产量表型的10 个亲本衍生的杂交种群体;②具有最高杂种优势的杂交群体;③具有最高产量一般配合力的10个亲本衍生的杂交种群体;④具有最高特殊配合力的杂交种群体(图4a)。
所有的组合均表现出比群体平均产量更高的表型。其中,具有最高特殊配合力的杂交群体表现出更高的产量优势,而具有最高产量表型的10 个亲本衍生的杂交种群体表现出最低的产量优势(图4a)。同时具有最高特殊配合力的杂交群体与具有最高杂种优势的杂种群体表现出高度的重合性(22 个家系是一致的),表明特殊配合力与杂种优势之间具有高度的一致性(图4b)。具有较高产量的杂交种群体在4 个群体组合中均有表现,具有最高产量表现的4 个杂交群体的单株产量均在50 g 以上,且在具有最高产量一般配合力的10 个亲本衍生的杂交种群体和具有最高特殊配合力的杂交种群体中均有分布,表明一般配合力和特殊配合力均对水稻的高产量和杂种优势具有贡献(图4b)。

图4 NCII群体各组分产量比较(a)和各组分之间相互关系的维恩图解(b)
3 讨论
杂种优势的应用大大提高了水稻、玉米等作物的产量,在杂交水稻育种过程中,选择优良亲本材料被认为是重要的成功因素。但是在1 个丰富多样性的群体中选取到理想的亲本材料是比较困难,具有较大的随机性并且费时费力,需要育种家们丰富的田间经验[8]。SCA 和GCA 已经成功地应用于优良亲本的鉴定进而培育优良的杂交组合材料。利用双列杂交的方法将遗传变异分解为加性变异和非加性变异,并以此估算GCA 和SCA 的遗传效应[7],为估算不同的杂交组合设计中的遗传变异和配合力的效应提供了理论基础。研究指出,亲本间的遗传距离和配合力及杂种优势表现正相关[9,10],而本研究结果表明,杂种优势与亲本间的遗传距离和杂合度之间并没有极显著的相关性。
杂交稻的产量性状的遗传力较低,易受包括环境因素在内的复杂成分的影响,另外,人类长期的育种选择所造成的加性遗传变量也可能与此相关。依据Kelly[11]提出的理论R=ih2s(R表示经过选择的一代的平均增加优势,i表示选择强度,h2表示遗传力的大小,s表示亲本的表型标准偏差),对特定性状的选择响应与该性状的遗传力是显著相关的。多次的人工选择趋向于优势加性效应基因的整合,进而导致遗传力和表型标准偏差下降。一旦优势基因被整合,就不会对选择进一步地响应。随着不断的选择性育种,越来越多的优势加性效应位点被整合,加性遗传变异和遗传力会越来越少。这将会造成加性遗传方差与非加性遗传方差的比值越来越小,导致对一般配合力的选择效率降低;为了使得后期选育的品种不断优于前期的杂交品种,对特殊配合力的选择效率要求则必然越来越高。
为了有更好的杂种优势表现,在实际育种中,建议结合分子标记辅助选择(MAS)体系,选择具有较高一般配合力的亲本进行随机杂交,然后在杂交材料中选择具有较高特殊配合力的材料进行定向育种改良,可以降低时间和人力成本,减小盲目性和随机性,实现分子层面的杂交育种和分子改良育种[12]。
本研究分析了GCA、SCA 对表型性状和产量杂种优势的相互关系和不同贡献率。如果性状具有较高的狭义遗传力,它们更多的是受加性效应决定的;相反,若性状具有的狭义遗传力很低,且与广义遗传力差值较大,则它们更多的是受非加性效应影响。表型值总是和GCA 和SCA 呈正相关,而杂种优势和SCA 呈正相关,GCA 与SCA 之间呈负相关或者没有相关性。即非加性效应对杂种优势的产生具有更大的贡献,加性效应和非加性效应对产量性状本身均有重要作用。通过比较不同参数在育种中的选择效率,可对以后的亲本选择、优良杂交材料构建和效应评价提出建议。其他作物如玉米可能具有与水稻相似的配合力遗传机制,但仍需要大规模的遗传设计试验进行研究。
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
养猪(2022年4期)2022-08-17
蔬菜(2021年7期)2021-11-27
猪业科学(2018年5期)2018-07-17
河北农业科学(2016年2期)2016-09-09
山西农经(2016年5期)2016-02-28
湖北畜牧兽医(2015年11期)2016-01-11
山东农业科学(2014年1期)2015-03-09
茶叶通讯(2014年2期)2014-02-27
生物学杂志(2013年1期)2013-04-11
