
子代发育管控(ODC):生物有性生殖进化的一个普遍趋势
2021-01-21 07:02刘建妮
西北大学学报(自然科学版) 2021年1期
傅 强,刘建妮,王 鑫,
(1.中国科学院 南京地质古生物研究所 生命演化与环境卓越创新中心 现代古生物学和地层学国家重点实验室, 中国 南京 210008;2.西北大学 地质学系 大陆动力学国家重点实验室 早期生命研究所, 陕西 西安 710069)
春天沙丁鱼会汇聚到西班牙北部14~15℃的水域里产卵[1]。
有些鱼会生下活生生的幼崽[2-4]。
寒武纪早期的朵氏小昆明虫在其双肢型的附肢上携带卵[5]。
有些小蜘蛛会杀死并吃掉一直携带并养育它们的母亲[6-7]。
一只雌螳螂在性交时会吞食它的伴侣[8]。
赤眼蜂把卵产在蛾子的卵里[9]。
大孢子和小孢子的分化始于泥盆纪[10]。
从泥盆纪晚期开始,胚珠就依附在它们的孢子体母株上[10]。
自中生代以来,被子植物的胚珠在受粉前就被封闭、保护起来了[11]。
红树直接生育出它们的幼苗[12]。
一些家长为他们的孩子支付名牌大学的入学费用[13]。
数百只企鹅站在凛冽的寒风中,用体温孵化处于它们包围之中的企鹅蛋。
……
动物和植物上述各式各样、令人称奇的行为引起了许多生物学家的关注,但是很少有人问,在这些不同的行为背后,有没有什么共同的规律。进化!是的。自从达尔文时代以来,进化是一个广泛接受的观念,并且人类已经花费数十亿美元研究地球生物的进化。然而,对许多人来说,进化似乎是复杂而又难以捉摸的事情。
基于对有性生殖过程和模式的研究和调查,我们认识到动植物生殖器官起源和进化中存在一个普遍的趋势,即所有的生物类群都倾向于越来越多地管控其子代的发育。我们把它称作子代发育管控(offspring development conditioning, ODC)。
如果生命被认为是有机分子间可进化的协调的相互作用模式,那么进化可以看作是穿越时间来维持和修改这种可控的相互作用模式的过程。维持谱系的延续对于所有生物来说是首要目标,也是终极目标,也是进化存在的前提。锁链容易在最薄弱的一环断开。生物中维持谱系的延续是通过持续重复有性生殖周期来实现的[14],每个个体的幼年是生命周期中最脆弱的时期。因此,对于所有生物来说,养育后代和保护后代远离危险是它们所要面对的主要挑战和任务。然而,不同的生物类群会通过不同的策略来实现这个相同的目标[11,15]。下面我们将分别浏览一下动物和植物的不同策略。
1 动物
目前,在分类学中,除了其他特征外,生物主要依据其生殖方式进行分类。在动物中,几种生殖方式是被公认的,即外卵生、内卵生、携卵生、卵胎生、组织营养型胎生和血液营养型胎生(见表1)[15]。这些模式按照上述顺序逐次为后代提供更加完善的保护,这也基本上是它们在生物演化历史中在相关类群中出现的顺序。这个顺序的背后存在着一个共同的潜在趋势,即子代的发育环境变得日益内化和受控(见表1、图1)。例如,受精部位从体外(鱼类在水中)(见图2A)到体内(爬行动物在子宫中)的转变(见图2C)。受精卵的孵化场所的位置发生了相应的变化:从鱼类的体外到小昆明虫[5]和瓦普塔虾[16]的母体表面,再到在某些两栖动物、爬行动物和哺乳动物的体内,而且在体内的驻留时间有一定的延长[17]。
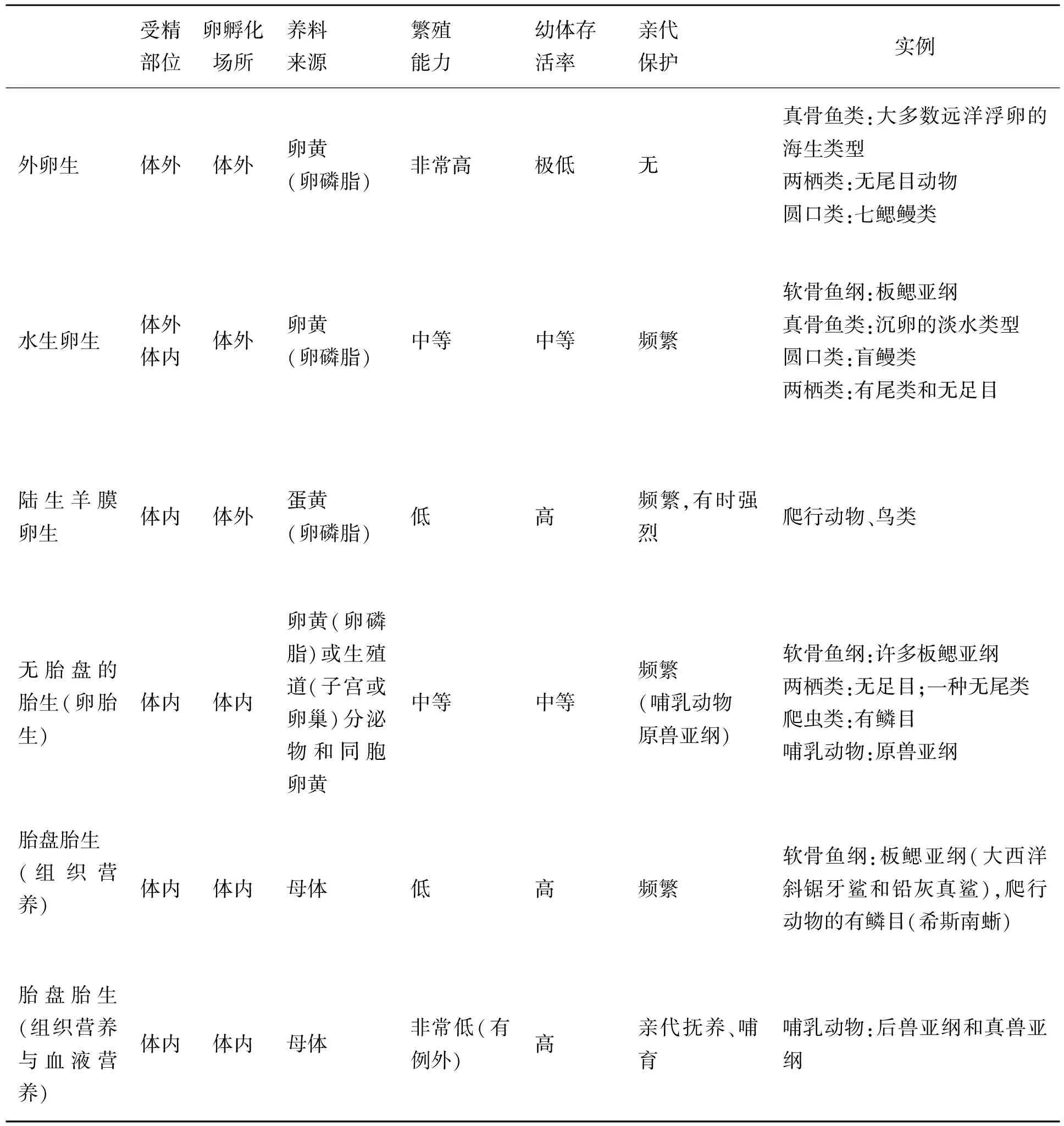
表1 脊椎动物各种生殖方式的主要特征。数据主要来源于[15]
这个过程可以称之为子代发育环境内化(offspring developmental environment internalization, ODEI),是实现子代发育管控(ODC)的途径之一,也和高等动物生殖器官起源和进化的机制密切相关。在子代发育环境内化过程中,幼体的营养得到了日益的增强、确保和延展,养料来源从简单的卵(鱼类中)、它们的同类(食卵性,蝾螈的子宫内嗜食同类[17])、输卵管的分泌物(昆虫和爬行动物中[17]),一直变化到类胎盘结构或一个真正的胎盘(哺乳动物中[17])。与此同时,后代和母亲之间的养育纽带在物理上日益加强并且在时间上有所延长。这些改变的结果是母体将会愈来愈多地控制它们孩子发育过程,而且幼崽会在出生之前获得愈来愈多的物理保护,并且会增强出生之后的存活能力。例如,小响尾蛇在出生时就有毒牙和毒液,并且很多哺乳动物在出生几分钟后就可以站立和行走。亲本动物似乎在它们幼崽生活中扮演愈来愈积极的角色。
为了应对各种生活中的挑战、对幼崽实施有效的保护,动物发展出各种适应策略并且经历了多个不同的适应阶段,包括被动适应、主动环境选择、主动环境调节。例如,鱼类经常在水体中产卵,如果水体适宜,它们的卵就会受精并且成功发育成幼鱼。然而,如果环境不理想,卵可能不会受精或根本不会发育。在这种情况下(被动的适应),卵是被动的:它们的命运更具偶然性。为了避免这种不确定性,一些鱼类在整个进化过程中进化出了一种固定的洄游习性作为应对策略。洄游使得鱼类能够在合适的时间来具有合适温度的水体中产卵、繁育。在这种情况下(主动地选择合适的环境),虽然卵的命运仍然由水体决定,但鱼类依赖从祖先那里继承下来的习性(智慧?)使得它们能够更加主动地为它们的子代选择合适乃至优越的发育环境。
一些无脊椎动物为了选择合适的环境采取了更为主动的携卵策略(见图3A),而且这种策略至少可以追溯到寒武纪早期(主动环境选择)[5,16]。“没有什么比我自身更好”,这可能是小昆明虫所采取的哲学思维(见图3B):通过随身携带卵,这些动物似乎有更多的自主性为它们的后代选择一个适宜的发育环境。通过卵胎生(见图3C)和组织营养型胎生,这种趋势在血液营养型胎生中达到顶峰,在这个过程中受精过程在体内进行。后代通过胎盘从母亲那里吸收营养并且在出生前与外界环境隔离。相比于卵生,子代发育管控程度较高的胎生使得采用这种生殖模式的动物具有成种快、物种更替率高等诸多优势,在历史上多次引发新物种和多样化的爆发(见图1)[4,18]。例如,胎生在鱼类中至少出现了29次[2,19,20],在无尾类中出现了数次[21],在蝾螈中出现了数次[22],在有鳞类中出现了6次[23]。

图1 脊椎动物的生殖模式及其在系统发育树上的分布[15]

A 鱼类的卵生; B 青蛙的卵生; C 爬行动物的卵生; D 哺乳动物(人类)的胎生

A 在体表携带卵的现代虾,插图显示附肢上的卵; B 小昆明虫(寒武纪早期),附肢上携带有卵; C 海洋卵胎生爬行动物凯科龙,其腹部有多个幼龙,插图详细展示了母亲身体里的后代(经彭光帆、金仁义、傅小平礼许,来源于《三叠纪公园》)。
虽然蛇类以卵生为特征,但并不是所有的蛇都是卵生的,例如响尾蛇就是卵胎生的[24]。在鸟类中没有胎生的生殖方式,但它们通过孵化和照顾幼鸟来进行相应的弥补。孵蛋过程中,鸟类通过长时间连续地坐在蛋上来温暖和保护它们的蛋。有几个科的鸟类属于罕见的例外:它们不孵化自己的蛋,而是在其他鸟类的巢中产卵(巢寄生)。例如,欧洲的杜鹃(鹃形目布谷鸟)和北美的牛鹂(雀形目褐头牛鹂)是众所周知的专性寄生生物。虽然它们不会孵化自己的蛋,但它们已经发展出足够先进的欺骗策略奴役其他的鸟为自己孵化蛋而且能够确保自己宝宝的安全[25]。大多数哺乳动物都是胎生的。这至少在一定程度上促成了今天哺乳动物的多样性(见图1)。单孔目动物作为哺乳动物中的一个例外,是唯一现存的卵生胎盘类动物(真兽类):就像有袋类动物一样,它们把未发育完全的幼崽放在一个育儿袋里养育、用乳汁来哺育。
很多动物通常在子代出生后继续维持着对它们的抚育,有时表现为利他主义。蜘蛛和许多哺乳动物用母乳哺育幼崽[26]。有些蜘蛛会携带它们的幼崽,并用它们本身的肉体作为哺育幼崽的营养[6-7]。螳螂在交配过程中吃掉雄性个体以获取营养和蜘蛛母体用自身作为营养哺育幼崽的行为具有异曲同工之妙[27]。我们人类更是将父母的管控延伸到极致,不仅包括青少年期的营养供给和人身保护(图2D),而且扩展到智力训练(例如大学教育)方面,而遗产作为留给子代的财富使得这种管控可能延及子孙多代[13]。
可以说,动物的类群和生殖模式之间没有严格的一一对应关系(见图1)。这不像在教科书上所教写的那样简单。相反,不同的动物谱系采用了不同的生殖策略的组合:在每个谱系中,增强了子代发育管控的策略都是后来进化出来的,这使得采取这种策略的动物在与同伴的竞争中具有更多的优势。较低等的动物类群在生殖方式上有着更宽泛的策略选择范围,而较高等的动物类群则被限制在更高级的少数几个策略选项之内。
2 植物
因为植物通常不会动,因而许多人误认为动物比植物更进化、高明。这种想法显然是错误的。如果调查植物的生殖行为,你会惊奇地发现植物和动物的生殖进化趋势完全一样(见图4A~I)。根据生殖方式,植物可分为两大类:孢子植物(早期陆生植物、蕨类)和种子植物(包括裸子植物、被子植物)。早期陆生植物和蕨类以孢子的形式传播(见图4A)。每个孢子在分散出去时,从母株中携带非常有限的营养,它的存活具有很强的偶然性,取决于它是否到达了一个适宜的环境,这个环境是否允许配子体的发育、受精以及下一代孢子体的萌发。如果一个孢子是不幸的,那么来自母株的有限的营养很快就会耗尽,这个自带的营养量就为孢子及其衍生物设定了生命力的上限。最初,所有孢子的形态都是相同的,即早期陆地植物在孢子大小、形态和营养分配上差异不大或者完全没有。然而,在泥盆纪中晚期衍生出的具异型孢子的植物在各个方面很快就出现了差异。
在异型孢子中, 大孢子获得的营养更多, 进而保证了雌配子体有更多的营养供应, 而小孢子只分配到有限的最基本的营养[28]。进化历史表明, 这种分化显然给了这些植物更多的优势, 因为目前生态系统中的最优势的植物就是异孢植物: 种子植物进一步将这种异孢趋势推向更高级的形式, 种子(见图4B)。在化石中, 有时很难从形态上区分种子和大孢子。然而, 在活的植物中很容易区分它们, 即大孢子在成熟时从母株上脱落, 而胚珠则驻留在母株上, 直到由它衍生而来的种子成熟为止[29]。与大孢子相比,种子的明显优势在于, 种子在成熟前一直保持着与母株的动态营养纽带。此外, 许多裸子植物还为它们成熟的种子提供各种额外的保护(图4C)。这种由母本植物为子代提供不断增强的营养纽带和物理保护是种子植物自起源以来一直遵循的大的进化趋势(见图4B~G)。
这种趋势在被子植物中达到了巅峰:被子植物在受粉前把它们的胚珠(种子的前身)包裹起来(被子)(图4D,F)。这种保护不仅可以保证胚珠的营养供应、保护脆弱的胚珠免受各种严酷的生物和非生物的伤害,而且通过额外的(通常是肉质的)保护层有利于种子的传播,为被子植物的生存提供了许多比较优势,使得被子植物成为当今世界上多样性最高的植物类群。沿着被子这一趋势进一步发展的是被果(见图4E,G)和胎生(见图4H)。在被果植物的果实中,包裹种子的果实被另一层的组织包裹起来(见图4E,G)。至少在现生的檬立木科[30-31]、蜡梅科[31]、桑科[32]、茄科[33],以及一些早期被子植物化石(梁氏朝阳序, 迪拉丽花)[11,34-35]中这种现象都可以看到。许多人认为胎生是哺乳动物特有的特征,他们很少知道被子植物也有这个特征。迄今为止,被子植物的近100种(23科40属)是具有胎萌行为的[36]。红树是一种典型的红树林植物。与大多数植物不同,红树的种子在其体内发芽,即在果实仍然附着在母株上时其位于果实中的种子就已萌发成幼苗(见图4H)。这意味着红树的幼苗孢子体是依靠其孢子体母株来发育的,直到幼苗达到一定的成熟度。

A 孢子植物中从孢子囊中分散出来的孢子; B 具有珠被保护的胚珠; C 开通类中有壳斗保护的胚珠/种子; D 被子植物中被子房保护的基生胚珠; E 一种酸浆属的果实里面有种子,果实周围又有一层保护层; F 被子植物中被下位子房保护的胚珠; G 被隐头果序保护的果实; H 红树在其母株上“胎生”的幼苗; I 兰科槽舌兰属植物的主动自控受粉
最后, 植物所采取的最具动物性的策略是兰科植物的主动自控受粉(PSF, proactive self-fertilization)。在主动自控受粉过程中, 通过其花药柄的有序运动, 将花粉囊插入等待花粉来受精柱头腔中[37](见图4I)。这是现在已知的植物自控行为的极致。在这种情况下, 植物不是依靠外部的非生物或生物因素来授粉, 而是通过自己雄蕊的一系列自主运动来完成授粉过程的。主动自控受粉使相关兰花在受粉过程中可以完全独立于在其他植物中不可缺少的传粉媒介, 保证了谱系的成功生殖和延续[37]。显然, 主动自控受粉给了这种兰科植物更多的自由、独立以及对生殖过程的完全控制。
3 结论
子代发育管控是大多数动植物有性生殖的基本进化趋势。首先,受精部位由低级生物的体外转移到较进化的更受生物本体控制的体内。第二,胚胎的发育部位在每个谱系中变得越来越趋向于体内。第三,在整个进化过程中,亲代和子代之间的营养纽带在物理上不断加强。第四,亲代和子代之间的营养纽带持续时间变得更长。第五,尽管方式和程度各有不同,至少大多数谱系朝着相同的方向进化出类似的生殖模式。因此,大多数生物(除某些寄生生物外)是奔着子代发育管控增强这个方向进化的。将原先的体外受精、发育场所内化为生物机体的一部分,从而实现管控子代的发育过程。这就是高等生物的生殖器官的起源方式和背后的逻辑。
缩写和术语
被子(angiospermy):受粉前胚珠(种子的前身)被包裹起来。
被果(angiocarpy):果实被其他组织包裹起来。
被子植物(angiosperms):字面意思是,种子被包裹起来的植物。严格地讲是胚珠被包裹起来的植物。后者可以确保一个植物的被子植物属性。
巢寄生(broodparasitism):一种寄生生物类型,依赖其他动物哺育自己的子代。
裸子植物(gymnosperms):一种受粉时胚珠裸露在外的植物。
血液营养型胎生(hemotrophicvivipary):受精和胚胎形成发生在雌性的生殖道,通过胎盘从母亲的血液里获取营养。
孢子异型(heterospory):一个植物产生两种不同的孢子。
结构营养型胎生(histotrophicvivipary):受精卵在雌性的输卵管中发育,从母体组织的分解和母体血液的渗漏中获得营养的胎生方式。
大孢子(megaspore):两种孢子中相对较大的一种,通常发育成雌配子体。
内卵生(ovipary):一种生殖策略,具有较少数量的较大卵,通常内部受精、直接发育,偶有亲代照料。
卵胎生(ovovivipary):幼体由受精卵发育而来,留在母体内但被卵膜与之分离的生殖策略。
携卵生(ovule-carrying):在孵化期间,将卵携带在动物的体表的生殖策略。
外卵生(ovulipary):生产大量小卵,受精和幼虫阶段都在体外的生殖策略。
寄生孵化(parasitichatching):一些寄主依靠其他动物来孵化自己子代。
亲代养育(parentalcare):亲代投入时间和精力来哺育和保护他们的子代。
主动自控受粉(proactiveself-fertilization):通过花药柄的自我驱动运动完成受粉的过程,花药柄将花粉块插入接受花粉的柱头腔。
种子(seed):由胚珠发育而来的植物繁殖器官,含有胚胎。
孢子(spore):能产生配子体的小型单细胞的繁殖体。
胎生(vivipary):在母体内长成的新的个体,或种子直接在果实里萌发成幼苗。
致谢:特别感激Jason Dunlop博士(柏林洪堡大学莱布尼茨进化与生物多样性研究所自然博物馆)提供的帮助。
猜你喜欢
中国生育健康杂志(2022年4期)2022-11-25
中国典型病例大全(2022年11期)2022-05-13
科学导报(2022年12期)2022-03-08
健康大视野(2020年17期)2020-09-15
家庭医学·下半月(2019年6期)2019-08-16
安徽农学通报(2017年9期)2017-05-19
中国医药导报(2017年6期)2017-04-06
农业与技术(2016年22期)2017-03-07
奥秘(2015年4期)2015-09-10
中学生物学(2008年12期)2008-12-27
