
基于不同基质刺芹侧耳菌丝生长与木质素降解酶的相关性研究*
2021-01-18 08:31赵翠敏邹亚杰胡清秀郑素月
中国食用菌 2020年11期
赵翠敏,杜 芳,邹亚杰,胡清秀,郑素月
(1.河北工程大学园林与生态工程学院,河北 邯郸 056000;2.中国农业科学院农业资源与农业区划研究所,北京 100081)
我国是农业生产大国,每年都会有大量的秸秆类、修枝类等农林废弃物产生。这些农林废弃物大多富含木质纤维素,是来源丰富、成本低廉、再生时间短的生物质资源,其堆积不仅会造成环境污染,而且是一种巨大的资源浪费[1-2]。食用菌属于大型真菌,可以以富含木质纤维素的材料为主要培养料[3],通过分泌一系列胞外酶将纤维素和半纤维素物质分解为可供菌体直接吸收利用的小分子碳源。但在木质纤维素中,纤维素、半纤维素和木质素相互交联,形成牢固的网络结构[4]。纤维素及半纤维素很容易被环境中的微生物降解,但木质素却不容易被分解。降解木质素能有效地促进纤维素、半纤维素的释放,是木质纤维素物质高效利用的关键。参与木质素降解的酶系主要由木质素过氧化物酶(lignin peroxidase,LiP)、锰过氧化物酶(manganese peroxidase,MnP)和漆酶(laccase)等组成,这些酶类合称为木质素降解酶或木质素酶[5-6]。
刺芹侧耳 [Pleurotus eryngii(DC.et Fr.)Quèl.],又名杏鲍菇,隶属于伞菌目(Agaricales) 侧耳科(Pleurotaceae) 侧耳属 (Pleurotus),是一种白腐真菌,具有较强的木质纤维素降解能力,可以在木屑、甘蔗渣、棉籽壳、玉米芯等多种农林废弃物上生长[7]。真菌对基质的利用和转化能力可以用其木质纤维素降解酶的活性来指示。鞠洪波[8]研究报道杏鲍菇对云杉木质素的降解能力在香菇、金针菇、平菇等9种食用菌中最强,而从多种新鲜食用菌子实体中提取漆酶,发现杏鲍菇漆酶的酶活性最高[9-10],工厂化杏鲍菇菌渣漆酶活性最高可以达到7.55×104U·g-1,明显高于桃红侧耳、香菇、白灵菇菌渣中的漆酶活性[11]。由此可见,杏鲍菇之所以生长速度快、生产周期短、易于栽培,除了其本身的生物学特性之外,还与其能够分泌高活性的木质素酶类紧密关联[12]。但在杏鲍菇的生产过程中,不同农林废弃物诱导菌丝体生产木质素酶类的酶活水平尚不清楚。通过以刺芹侧耳2611为试验材料,研究其在7种农林废弃物培养基质上的菌丝生长速度,并且测定其液体发酵胞外产漆酶、木质素过氧化物酶的情况变化,探索刺芹侧耳对不同栽培基质的降解利用规律,为优化刺芹侧耳新型栽培基质提供依据。
1 材料与方法
1.1 材料
1.1.1 供试菌株
刺芹侧耳(Pleurotus eryngii)2611,来自于中国农业微生物菌种保藏管理中心,保藏号ACCC52611。
1.1.2 供试材料
棉籽壳、木屑、玉米芯、玉米秸秆、甘蔗渣、花生壳、柠条7种农林废弃物,各栽培基质中的纤维素、半纤维素和木质素的含量[13-16]如表1所示。

表1 供试原料及组成Tab.1 Test material and composition
1.1.3 培养基
PDA培养基:15.6 g DifcoTM Potato Dextrose A-gar(美国 BD公司) 溶于 400 mL的 ddH2O中,121℃灭菌 15 min。
农林废弃物培养基各组分见表2。
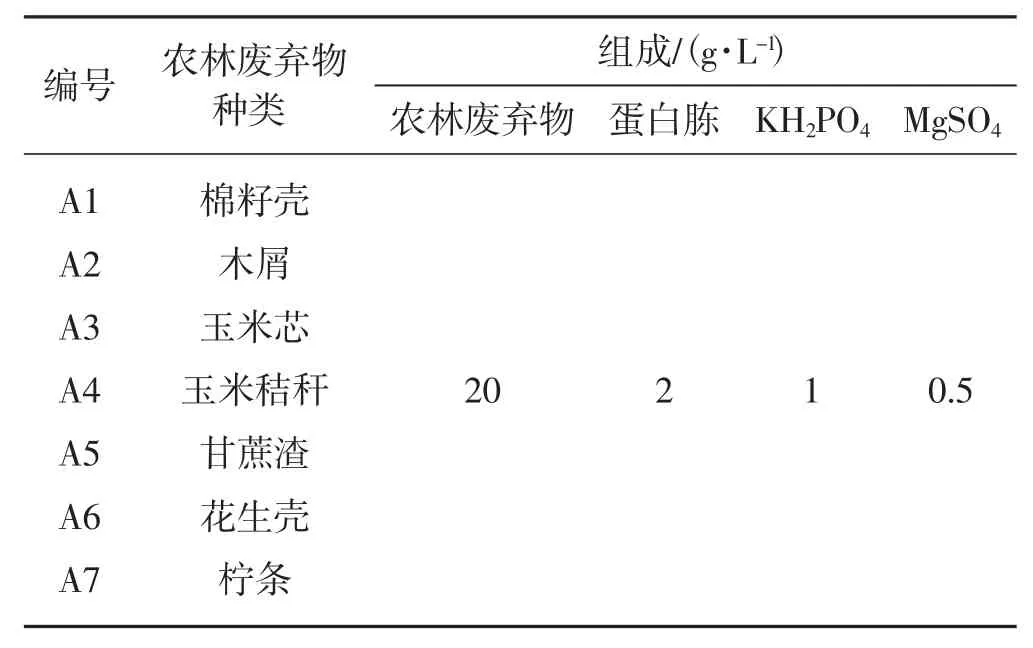
表2 培养基组分Tab.2 The medium formula
如表2所示,固体和液体培养基组分相同,固体培养基配制时加入20 g·L-1的琼脂。定容至1 L,121℃,灭菌30 min。
1.2 方法
1.2.1 平板培养试验
从PDA平板上取0.5 cm×0.5 cm菌种块接种于不同基质的固体培养基的平板。28℃恒温黑暗培养5 d观察记录菌丝生长情况,并测定菌丝生长速度。
1.2.2 发酵培养试验
在250 mL的三角瓶中加入100 mL相应的发酵液灭菌,在无菌环境中每瓶接种10 mL的原种,置于28℃摇床内黑暗振荡(180 r·min-1)培养5 d。原种由2个刚长满菌丝90 mm×90 mm PDA平板加入100 mL农林废弃物培养基中,经匀浆仪20 s混匀得到。
1.2.3 粗酶液制备
将发酵液转移至离心管中,12 000 r·min-1离心10 min,上清液即为粗酶液样品。
1.2.4 漆酶活性测定
ABTS法测定发酵液漆酶酶活[17]。将100 μL的粗液样品与190 μL的底物ABTS混匀,30℃反应10 min后,加入300 μL的5℅ TCA溶液终止反应,420 nm处测定OD值。灭活酶液作为对照,对照管提前终止。酶活单位定义为每分钟氧化1 μmol底物所需的酶量为一个酶活力单位(U·mL-1),漆酶活力(E,U·mL-1)计算公式为:
式中:ε为ABTS摩尔消光系数(36 000 L·mol-1cm-1);Δt为酶反应时间(min);ΔOD为吸光度的变化值;V1为酶反应中反应液的总体积(mL);V2为酶反应中,酶液的总体积(mL);N为酶液稀释倍数。
1.2.5 木质素过氧化物酶活性测定
木质素过氧化物酶活性的测定使用北京索莱宝生物科技有限公司的木质素过氧化物酶试剂盒。每个处理3个生物学重复。酶活性定义:每升培养液每分钟氧化1 nmol藜芦醇所需的酶量为一个酶活力单位。木质素过氧化物酶活性(P,nmol·min-1L-1)计算公式为:
式中:ε为藜芦醛摩尔消光系数,9 300 L·mol-1cm-1;d为比色皿光径1 cm;V反总为反应总体积1 mL;V样为反应中样本体积0.1 mL;T为反应时间5 min;ΔA为OD310的变化值。
1.2.6 数据分析
生长速度、酶活和生物量变化数据均为3次重复试验的平均值±标准偏差(SD),小写字母为使用IBM SPSS Statistics 25软件计算的显著性差异(P<0.05),采用Duncan方法进行多重比较。
2 结果与分析
2.1 不同培养基质对菌丝生长的影响
不同栽培基质上的菌丝生长速度、生长势和菌丝的浓密度不同,见图1、表3。
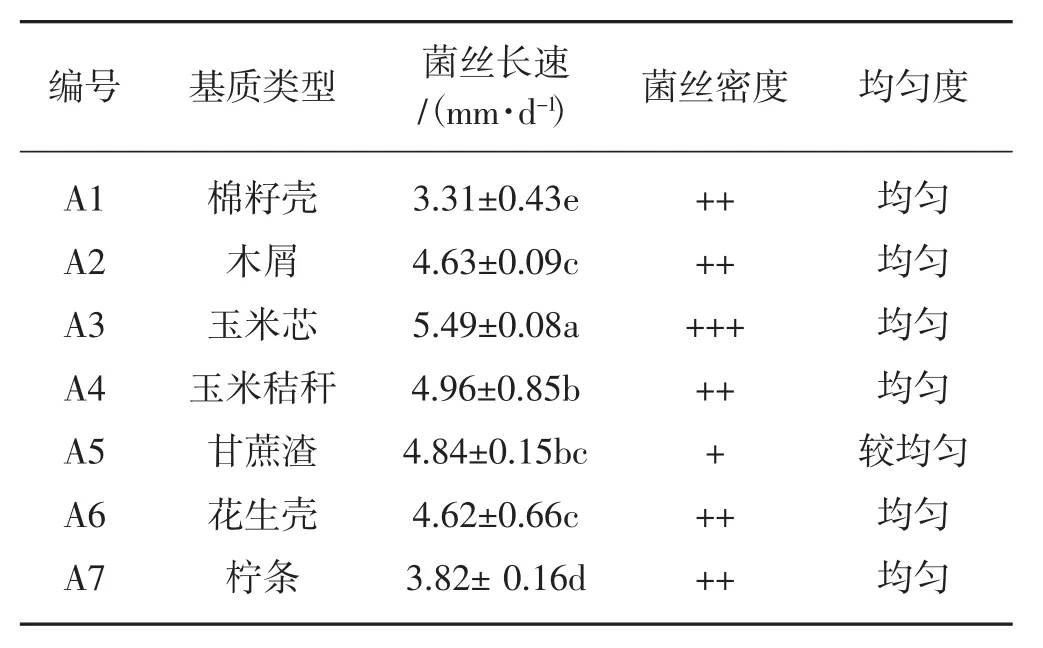
表3 不同基质对菌丝长势的影响Tab.3 The effects of different media on mycelium growth
从图1、表3可以看出,在玉米芯基质上,菌丝生长速度最快(5.49 mm·d-1),且菌丝整齐浓密,长势良好;其次菌丝生长速度由快到慢依次是玉米秸秆、甘蔗渣、木屑、花生壳、柠条、棉籽壳。其中玉米秸秆基质生长速度(4.96 mm·d-1)和甘蔗渣基质生长速度(4.84 mm·d-1)差异不显著,木屑基质生长速度(4.63 mm·d-1)和花生壳基质(4.62 mm·d-1)没有显著差异。甘蔗渣基质上菌丝稀疏,菌丝较均匀。综合菌丝生长速度、菌丝浓密度、菌丝均匀度分析,以棉籽壳、柠条、甘蔗渣为碳源的培养基质杏鲍菇的菌丝长势明显弱于以其他材料为碳源基质的杏鲍菇菌丝长势,说明以玉米芯、木屑、玉米秸秆和花生壳为碳源有利于菌丝的生长,杏鲍菇对4种基质的降解利用效果更好。
2.2 不同培养基质诱导菌丝体产漆酶的能力比较
不同培养基质液体发酵液中的漆酶酶活性有显著的差异,见表4。

表4 不同基质对漆酶酶活的影响Tab.4 The effects of different substrates on laccase activity
从表4可以看出,在花生壳培养基质中,漆酶的酶活最高(109.15 U·mL-1),其它培养基质中漆酶酶活由高到低依次是玉米秸秆、玉米芯、木屑、柠条、甘蔗渣、棉籽壳,在棉籽壳培养基质中漆酶酶活最低(30.08 U·mL-1)。其中玉米芯基质中的漆酶酶活(76.62 U·mL-1)与玉米秸秆基质中的漆酶酶活(86.62 U·mL-1)无显著差异,木屑基质中的漆酶酶活(63.73 U·mL-1)与柠条基质中的漆酶酶活(57.60 U·mL-1)差异也不显著。白腐菌的生长与漆酶的分泌具有一定的相关性,一般情况下,生长状况良好的白腐菌分泌的漆酶酶活性高,生长状况差的白腐菌分泌的漆酶酶活性低,但是白腐菌的生长和产漆酶并不同步,没有明显的正相关性[18]。在本研究中,杏鲍菇在花生壳、玉米秸秆、玉米芯基质中分泌漆酶的酶活性高,而在棉籽壳、柠条、甘蔗渣基质中分泌胞外漆酶的酶活性低。和已有的研究结果基本一致,漆酶的酶活性与菌丝长势相关,菌丝长势越好漆酶酶活越高[19]。
2.3 不同培养基质诱导菌丝体产木质素过氧化物酶的能力比较
不同培养基质液体发酵液中的的木质素过氧化物酶的酶活性有显著的差异,见表5。
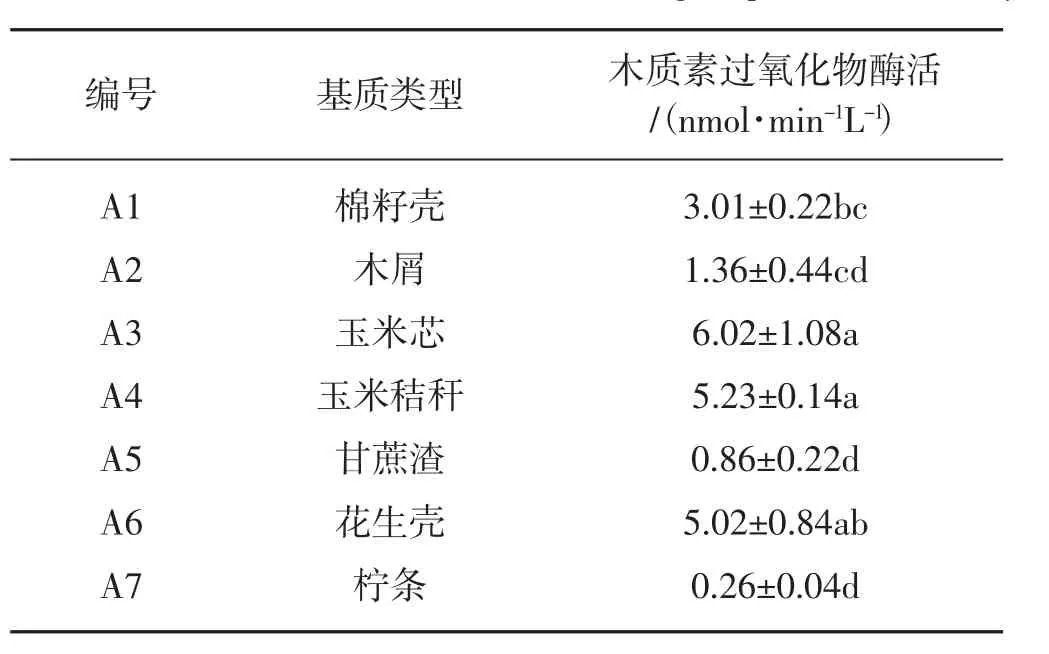
表5 不同基质对木质素过氧化物酶酶活的影响Tab.5 The effects of different substrates on lignin peroxidase activity
从表5可以看出,在玉米芯培养基质中,木质素过氧化物酶的酶活性最高(6.02 nmol·min-1L-1)。其它培养基质中木质素过氧化物酶的酶活性由高到低依次是玉米秸秆、花生壳、棉籽壳、木屑、甘蔗渣、柠条。其中玉米芯培养基质中的酶活性(6.02 nmol·min-1L-1)与玉米秸秆培养基质中的酶活性(5.23 nmol·min-1L-1)无显著差异,甘蔗渣培养基质中的酶活性(0.86 nmol·min-1L-1)与柠条培养基质中的酶活性(0.26 nmol·min-1L-1)无显著差异。柠条培养基质中酶活性最低(0.26 nmol·min-1L-1)。木质素过氧化物酶活性在柠条、甘蔗渣、木屑、棉籽壳的培养基中,活性明显低于其它培养基。以上结果表明,胞外木质素过氧化物的酶活性与菌丝长势具有一定的相关性,酶活越高菌丝长势越好。
3 讨论
通过对刺芹侧耳在不同培养基质上菌丝生长速度、菌丝的长势和胞外酶的变化规律进行研究,发现不同培养基质对菌丝的生长和木质素降解酶的活性均有明显影响(P<0.05)。菌丝长势参差不齐、菌丝生长速度差异大,在玉米芯基质上菌落最大,菌丝浓密,其次是玉米秸秆基质,说明玉米的废弃物适合刺芹侧耳菌丝的生长。已有研究表明食用菌菌丝的生长速度不仅与栽培料的C/N和N浓度有关,更与栽培料种类及其透气性密切相关;菌丝生长良好是食用菌高产的基础,但是不同栽培基质的菌丝满袋时间及菌丝浓密度与其产量相关性不显著,菌丝的生长情况与产量不呈正相关[20-22]。根据本研究结果,可以在杏鲍菇的生产中栽培基质中加入合适比例的玉米芯和玉米秸秆将有助于菌丝的生长。
漆酶是与木质素等大分子物质降解有关的主要酚氧化酶,在木质素的分解过程中起着重要的作用,属于诱导酶类,其活性会受到诸多因素的影响,如碳源、氮源、金属离子和激活物、pH及菌株等[20]。杂木屑、棉籽壳、玉米芯、甘蔗渣都是食用菌生产的主要碳源,其都含有大量的木质素,但含量不同,木质素含量高更有利于漆酶的诱导表达[23]。韩嘉钰等[24]对不同栽培基质对灵芝胞外酶活的影响开展研究,发现在菌丝生长阶段玉米芯培养基质中漆酶的酶活性比棉籽壳和木屑中的漆酶酶活性要高,与本试验结果一致。赵风云等[25]研究以蔗渣和棉籽壳为基质的灵芝酶活性中,发现棉籽壳培养基质中的漆酶酶活性高于蔗渣,与本试验中甘蔗渣漆酶酶活性高于棉籽壳结果不一致。安琪等[26]针对不同碳源和氮源开展对金针菇降解木质纤维素酶活性的影响的研究,表明与纤维素酶和半纤维素酶相反,金针菇在复杂氮源培养基上生长时漆酶活性相应低于简单氮源培养基。在本研究中,花生壳培养基质中漆酶的酶活性最高(109.15 U·mL-1),棉籽壳基质中漆酶酶活最低(30.08 U·mL-1)。花生壳的木质素含量最高,菌丝生长需要分泌更多的漆酶来降解木质素,因此在花生壳培养基质中酶活最高。但是棉籽壳的木质素含量在7种农林废弃物中不是最低的。木质素含量只是其中一个因素,不是决定性的因素,可能还和栽培基质的结构等其它因素有关。
木质素过氧化物酶是第一个从黄孢原毛平革菌(Phanerochaete chrysosporium) 发现的木质素降解酶,在木质素降解中起关键性作用[27]。丁少军等[28]研究了不同培养条件对云芝木质素降解酶产量的影响,发现木质素过氧化物酶、锰过氧化物酶和漆酶的产生与培养条件关系很大。彭木等[29]在研究帽儿山采集的禅剥管菌时表明不同底物能极大地刺激相应酶的产生与分泌。据报道,木质素含量越高越能促进木质素分解酶的产生[30]。在本研究中,玉米芯培养基质中木质素过氧化物酶的酶活性达到最高(6.02 nmol·min-1L-1),柠条培养基质中木质素过氧化物酶的酶活性最低(0.26 nmol·min-1L-1)。总体来看,除了少数差异以外,木质素过氧化物酶的酶活性与漆酶的酶活性表达趋势基本一致。
综上所述,通过对刺芹侧耳在不同培养基质上菌丝生长势、长速分析并进行了对应的液体培养,测定了木质素降解的关键酶,初步明确了不同种类的农林废弃物的木质纤维素的组成影响刺芹侧耳的降解能力,哪种类型的农林废弃物更有利于刺芹侧耳的吸收利用,为进一步筛选适宜刺芹侧耳利用生长的栽培基质提供参考。
猜你喜欢
造纸信息(2022年8期)2022-11-10
大连工业大学学报(2022年1期)2022-03-14
重庆科技学院学报(自然科学版)(2021年5期)2021-11-10
恋爱婚姻家庭·养生版(2021年7期)2021-08-23
恋爱婚姻家庭(2021年21期)2021-08-03
新能源进展(2020年1期)2020-03-09
农家致富顾问·上半月(2019年6期)2019-09-10
中国造纸(2019年6期)2019-09-10
分析化学(2018年7期)2018-09-17
农家顾问(2016年7期)2016-05-14
