
中国南方生长于不同基质的天然林植物叶片元素含量特征比较
2021-01-15 10:18:26崔培鑫申智骅付培立白坤栋姜艳娟曹坤芳
生态学报 2020年24期
崔培鑫,申智骅,付培立,白坤栋,姜艳娟,曹坤芳,*
1 广西大学, 亚热带农业生物资源保护与利用国家重点实验室, 南宁 530004 2 广西大学林学院, 广西森林生态与保育重点实验室, 南宁 530004 3 中国科学院西双版纳热带植物园, 热带森林生态重点实验室, 勐腊 666303 4 广西壮族自治区中国科学院广西植物研究所, 桂林 541006
自然界中的植物有机体都是由多种元素按照一定比例构成,养分元素的含量状况在植物生长和发育过程中有着重要的作用,而植物体的元素成分可以反映其在某种土壤环境下对元素的吸收或积累[1- 4]。因成土母质和风化环境等差异,不同生境的土壤发育和养分状况有明显差异,改变了植物对矿质元素的吸收和利用。基于特定生境基质上生长的森林生态系统,生态学家对植物养分吸收和利用的特征、机理进行了大量探索。
喀斯特森林是生长在可溶性碳酸盐岩(石灰岩、白云岩)上的生态系统,由于母岩母质中富含Ca、Mg,土壤呈碱性,pH值高,且土层浅薄不连续,持水能力差,养分贫瘠,生境十分脆弱[5]。周运超[6]对贵州喀斯特植物的研究中,得出喀斯特植物的Ca、Mg含量要明显高于酸性土壤植物,并且根据Ca+Mg量化归类,定义了不同钙生特性植物的吸收范围。罗绪强等[7]研究发现,喀斯特钙生植物主要受P的制约,具有低P、K和高Ca、Mg的特点,主要受到特殊土壤地质背景和环境因素的影响。杨成等[8]对喀斯特山区植物研究表明,不同生境下植物的养分含量存在差异,石灰土上植物的N、P、K、Ca、Mg、Al、Cu、Mo元素高于酸性黄壤上的植物,S、Fe和Mn反之。
我国南方地区降雨量大温度高,土壤淋溶作用强烈,非喀斯特地区分布大量的酸性土壤(pH值低),由于土壤中铁铝氧化物的吸附作用降低了养分的有效性,加之土壤本身养分缺乏、离子不平衡,严重影响植物的生长。Vitousek[9]对各气候带植物养分利用效率的分析认为P是限制热带植物生长的关键营养元素;Güsewell[3]也认为植物的生长多数会受P元素的限制,主要因为相对N,环境为植物提供的可直接被吸收利用的P较少。前人对酸性土植物的研究也显示植物主要受到P素的限制,莫江明等[10]对鼎湖山植物的研究提出,P和Mg对南亚热带常绿阔叶林植物的生产力有限制作用。阎恩荣等[11]和黄小波等[12]分别对浙江天童常绿阔叶林和云南季风常绿阔叶林的研究表示其受到P的限制。而吴统贵等[13]对珠三角典型森林乔木叶片的分析发现,常绿阔叶林和部分阔叶树种生产力受到N的限制。
红树林是生长在滨海湿地的典型森林系统,土壤为高度盐渍化的砂质土,含有呈还原态的酸性硫酸盐[14]。王文卿和林鹏[15]研究认为,秋茄和红海榄叶片高Ca和高Na、Cl含量的特点是它们应对盐胁迫的策略,可以减轻盐分对植物的伤害。刘滨尔等[16]对秋茄幼苗的盐分梯度研究发现,随着盐度增高幼苗对Na和Cl的选择性吸收增强,同时制约K、Ca、Mg的吸收。而邹晓君等[17]对3种华南园林植物的研究也得出,在盐胁迫下植物会增强对Na和Cl的吸收。可以看出红树林土壤基质中高盐的特点是影响植物体养分吸收的重要因素。
植物有机体在环境养分的平衡供给和体内代谢的内稳态中得以生长和发育,其元素含量都是按照一定比例存在的,某种元素的变化一般会对其它元素产生协同影响,在生态化学计量学中通常会以有机体的元素比值来表示元素间的相互作用[18]。植物体内的N/P比值可以作为植物生长的N、P营养限制指标[11,19],当N/P<14时,暗示植物生长受N限制,当N/P>16时,植物生长受P限制,当N/P比值介于14—16时,表明植物生长受N与P共同限制[20]。Du等[21]通过对中国西南喀斯特地区植物叶片N/P比值的研究,认为喀斯特地区植物一般受到P限制或N和P的共同限制。樊月等[22]对漳江口红树植物和黎洁[23]对广西北仑河口红树植物的研究均表明红树受到N限制。而在酸性土壤低pH的环境中易降低P的可溶态,减少植物对P的吸收,使植物的N/P比多大于16,表现为P限制。
植物营养元素在植物生长过程中相互作用,主要表现为拮抗作用和协同作用[24]。有研究认为,K+与Ca2+两个阳离子具有拮抗关系,在低钾时会有利于钙的吸收,高钾时K+大量进入细胞,会阻碍了Ca2+的吸收[25],而当土壤Ca含量丰富时,植物吸收大量的Ca2+会对K+产生拮抗作用[26]。酸性土植物Mn毒问题严重,在低pH环境下,吸收P元素能显著减轻Mn对植物的毒害[27],增加Ca和Fe的吸收也能有效拮抗Mn过量积累[28-29]。盐生环境下,K会对Na的吸收产生拮抗,使植物保持较高的K/Na比[30],避免盐胁迫伤害。总体上看,我国对于植物叶片养分含量特征的研究主要基于某一特定类型的生境,而对多种基质上的森林系统的比较研究还鲜有报道,对不同森林植物叶片养分含量的丰度和元素计量关系的研究,可以从元素计量学角度揭示植物对特殊环境的适应特点。本研究对基质条件迥异的3种生境,热带喀斯特森林、红树林和亚热带常绿阔叶林的植物叶片养分含量进行比较分析,试图回答以下问题:(1)各生境植物对哪些营养元素存在富集或缺乏状况?(2)不同生境植物叶片养分计量关系具有什么样的差异?
1 研究方法
1.1 研究区概况
(1)中国科学院西双版纳热带植物园(101°25′E,21°41′ N)绿石林森林公园位于云南省西双版纳勐仑镇(图1),属热带季风气候,年平均气温21.4℃,最热月均温25.3℃,最冷月均温15.6℃,年降雨量在1556.8 mm。该地区为典型热带喀斯特地貌,多峰丛、沟谷,土壤为石灰岩基质上发育淋溶腐殖质碳酸盐土,呈碱性(pH在7.29左右),在岩石露头地段,地面土壤复盖物少,土层薄,有机质层薄。受断块差异抬升及河流强烈下切的影响,这些石灰岩山地形成一种顶部保存有峰丛,边缘坡度较陡的中山和低山地貌,河谷多为深而陡的峡谷或嶂谷[31]。森林属典型的石灰岩山森林植被,生长有石灰山特征种如:榆科的油朴(CeltisphilippensisBlanco)、漆树科的清香木(PistaciaweinmanniifoliaJ. Poiss. ex Franch.)、百合科的开口箭(CampylandrachinensisBak.)、翅子藤科的扁蒴藤(Pristimeraindica(Willd.) A. C. Smith)、龙舌兰科的龙血树(Dracaenadraco(L.) L.)等。

图1 研究地点区位图Fig.1 The location map of study sites
(2)选取纬度相近的热带喀斯特森林对比分析,该对照点为广西弄岗国家级自然保护区(106°42′28″—107°04′54″E,22°13′56″—22°39′09″N),位于崇左市龙州县和宁明县境内(图1),属北热带季风气候区,年平均气温22℃,年平均降雨量1150—1550 mm。保护区为北热带裸露型喀斯特区域,岩性为灰岩、白云质灰岩、白云岩等,地貌以峰丛深切圆洼地为主,由多个石山山峰和镶嵌其中的洼地(谷地)组成。土壤呈碱性,pH值在7.87左右,在山谷中,土壤类型为水化棕色石灰土为主,在山坡上,土壤类型是棕色石灰土,山峰以黑色石灰土为主[32]。植被为热带喀斯特季节性雨林,在山谷和洼地中,分布着一些喜湿耐阴物种,如唇形科的垂茉莉(Clerodendrum_wallichiiMerr.)、桑科的对叶榕(FicushispidaL. f.)、叶下珠科的日本五月茶(AntidesmajaponicumSieb. et Zucc.)等;坡地阳光充足,但保水性差,分布有大戟科的肥牛树(Cephalomappa_sinensis(Chun et How) Kosterm.)、楝科的割舌树(Walsura_robustaRoxb. Hort.)、桑科的米扬噎(Streblus_tonkinensis(Dub. et Eberh.) Corner)、锦葵科的苹婆(Sterculia_monospermaVentenat)和蚬木(Excentrodendron_hsienmu(Chun et How) H. T. Chang et R. H. Miau)等;山顶部高温干旱,生境较为恶劣,有漆树科的清香木(PistaciaweinmanniifoliaJ. Poiss. ex Franch.)等耐旱植物。
3)海南清澜港红树林自然保护区(110°30′ —110°02′E,19°15′ —20°09′N)位于海南省文昌市内(图1),是我国红树林种类最多、面积最大、生长较茂盛的地区,红树林面积8.35 km2[33]。该区属于热带季风气候类型,年均气温在24℃,年平均降水量1623.5 mm。土壤成土母质主要为沙土,发育成细粉砂软泥和含淤泥的粗粉砂及细砂土[34],pH值在7.89左右,偏碱性。该红树植物群落主要以海桑(Sonneratiacaseolaris(L.) Engl.)和海莲(Bruguierasexangula(Lour.) Poir.)占优势,主要有以下群系:海桑群系、杯萼海桑(SonneratiaalbaSm.)群系、黄槿(HibiscustiliaceusLinn.)+海桑群系、榄李(LumnitzeraracemoseWilld.)群系,海莲+木果楝(XylocarpusgranatumKoenig)群系等[35]。
4)广东南岭国家级自然保护区(112°30′ —113°04′ E,24°37′ —24°57′N)位于广东省北部(见图1),属亚热带季风气候和山地气候,年均温17.7℃,年均降水量1705 mm。受强烈造山运动的影响,保护区内以皱褶构造发育的中山地貌为主,基岩主要为砂岩、页岩、花岗岩等,土壤为红壤和黄壤,呈强酸性(pH在4.89左右)。植被由常绿阔叶树与一定数量针叶树混交的中山亚热带常绿阔叶林为主,它们当中的壳斗科的丝栗栲(CastanopsisfargesiiFranch.)、甜锥(Castanopsiseyrei(Champ.) Tutch.)、水青冈(FaguslongipetiolataSeem.),樟科的竹叶木姜子(LitseapseudoelongataLion)、黄樟(Cinnamomumparthenoxylon(Roxb.) Kosterm.)、凤凰润楠(MachilusphoenicisDunn),金缕梅科的马蹄荷(Exbucklandiapopulnea(R.Br.) R. W. Brown)、阿丁枫(Altingiachinensis(Champ. ex Benth.) Oliv. ex Hance)、枫香(LiquidambarformosanaHance)等是南岭保护区植被的表征类群[36]。
1.2 采集与测定
本研究选取生境内的优势物种和频繁出现的常见树种进行研究,2006年 8至9月,在西双版纳绿石林森林公园采集了23个物种(每种选取3—5株成年植株),2013年7月,在弄岗热带喀斯特森林采集40个物种(每种选取3—10株成年植株),2015年8至9月,于海南清澜港红树林采集植物12种,(每种选取3株成年植株)。在雨季采集植物中上层新鲜成熟叶片,将同种植物叶片混合后装入干净自封袋,带回实验室。叶片洗净、晾干,100℃杀青后,烘干至恒重,用自动球磨仪研磨粉碎过60目筛保存备用。植物叶片化学元素含量分析依据来自《森林植物与森林枯枝落叶层全硅、铁、铝、钙、镁、钾、钠、磷、硫、锰、铜、锌的测定》 (LY/T 1270—1999)。植物叶片C、N含量采用Vario MAX CN分析仪(Elementar Analysensysteme, Hanau, Germany)测定,样品粉末通过HNO3-HClO4消煮,盐酸酸解,过滤,然后滤液用感应耦合等离子发射光谱仪(ICP AES-iCAP6300, Thermo Fisher Scientific, MA, USA)测定P,K,Ca,Mg,S,Fe,Mn,B,Zn和Na元素含量。而Si的测定采用质量法,酸解过滤后,滤纸连同残渣一起放入坩埚中105℃烘干,然后移入高温炉中800℃灼烧,冷却称重。广东南岭国家自然保护区植物元素含量数据来自许伊敏、旷远文等的研究[37],该研究对象为16个物种的成熟叶,每个种采集3—5个个体。
1.3 数据处理
使用Excel 2016对数据进行初步整理,应用SPSS 20软件对实验数据进行处理和统计分析。采用One-Sample T Test来分析植物各元素与N的比值和全国水平[38]之间有无显著性差异,因文献[38]中缺少C、Zn、B元素与N的计量比数据,本文使用陆生高等植物需求浓度中C、Zn、B和N的数据[39],并计算其计量比作为补充对照。认为显著性水平为P<0.05时才具有显著差异,P<0.01时为极显著差异。在对叶片元素之间进行线性相关性分析时进行了以10为底的对数转换,以改善数据的正态性。图形制作使用SigmaPlot 12软件。
2 结果与分析
2.1 各生境植物叶片元素的富集与缺乏
2.1.1喀斯特森林植物叶片元素丰缺状况
由表1看出,西双版纳喀斯特森林23种植物叶片中Ca与N的计量比值显著高于全国水平(P<0.001),具有Ca富集特征,研究的物种中,油朴、南酸枣、鸡骨香、槟榔青、豆果榕、大果山香圆、石筋草的Ca含量较为丰富,均高于30 g/kg(表2)。C、K、Na、Zn元素与N的计量比值显著低于对应的全国和陆生高等植物的计量水平(C、K、Na(P<0.001),Zn(P<0.05)),存在缺乏的状况。像油朴、石筋草的C含量均不超过400 g/kg;大苞藤黄、闭花木、南亚岩豆树、团花树、番龙眼、浆果楝、清香木的K含量较低,均少于5 g/kg;油朴、团花树、藤春、鸡骨香、浆果楝、榕树Na的含量均低于0.03 g/kg;毛紫薇、油朴、闭花木、团花树、藤春、槟榔青、坡垒、榕树、清香木、大果山香圆、披针叶楠、石筋草、蛛毛苣苔的Zn含量低,均少于0.02 g/kg(表2)。西双版纳喀斯特森林植物N/P均值为14.27,介于14至16之间,说明存在着N与P的共同限制。
由表1,弄岗喀斯特森林40种植物叶片K、Ca、Mg、S、Zn与N的计量比值有显著高于对应的全国和陆生高等植物的计量水平(K、Ca、Mg、S(P<0.001),Zn(P<0.05)),含量丰富。其中,白饭树、清香木、红背山麻、米扬噎4种植物含有丰富的K,均高于20 g/kg;白饭树、黄梨木、清香木、野漆、红背山麻、米扬噎、南酸枣的Ca含量高,均高于30 g/kg;白饭树、清香木、红背山麻、闭花木、南酸枣富Mg,含量均高于10 g/kg;火果、斜叶榕、构树、木蝴蝶、灰毛浆果、南酸枣的Zn皆高于0.15 g/kg;S含量高的植物有火果、构树、灰毛浆果、南酸枣,均大于4 g/kg(表3)。弄岗植物P、Fe与N的计量比值显著低于全国水平(P<0.001),表明含量缺乏。如鲫鱼胆、米仔兰、桑枝米碎、山榄叶柿、铁榄、鱼骨木、云南倒吊笔、清香木、紫弹树、闭花木、秋枫、蚬木几种植物的P含量较低,均小于1 g/kg;细子龙、铁榄、鱼骨木、黄梨木、割舌树的Fe含量较低,均小于0.10 g/kg(表3)。弄岗喀斯特森林植物N/P均值是18.26,受到P的限制。

表1 不同生境植物叶片元素含量及与N比值的百分比的平均值/(g/kg)
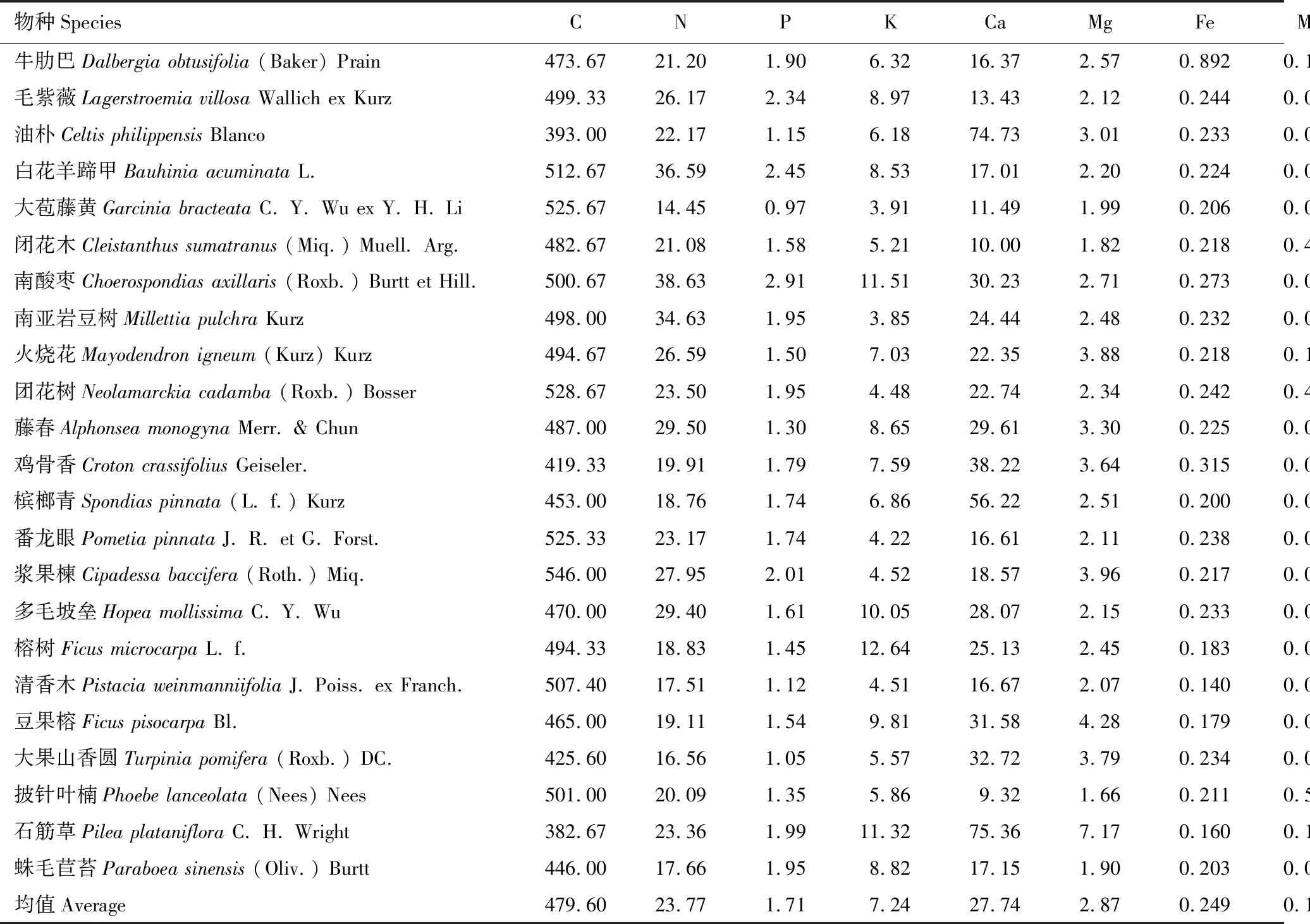
表2 西双版纳喀斯特森林植物叶片元素含量(3—5个个体的平均值)和N/P比值/(g/kg)
2.1.2红树林叶片元素丰缺状况
由表1看出,红树林12种植物叶片P、Ca、Mg、Na、S与N的计量比值显著高于全国水平(Mg、Na、S(P<0.01),P、Ca(P<0.05)),存在富集特征。所研究物种中,海桑与海南海桑有较高的P含量,都大于2 g/kg;木果楝、海漆、尖瓣海莲、榄李的Ca含量高,均大于15 g/kg;Mg含量较高的植物有瓶花木、榄李、黄槿,均高于6 g/kg;除银叶树、黄槿两种半红树,其他红树植物普遍富集大量的Na,均在10.38—3.15 g/kg范围内;海漆、尖瓣海莲、海莲的S含量较高,均大于4 g/kg(表4)。红树植物Fe、Si、Zn与N的计量比值显著低于全国和陆生高等植物的计量水平(P<0.01),说明其含量缺乏。如木果楝、瓶花木、银叶树、正红树、海莲、黄槿的Fe含量较低,均少于0.10 g/kg;瓶花木、榄李、海莲、杯萼海桑的Si含量低,均少于0.50 g/kg;Zn的含量普遍较低,除瓶花木、海漆、海桑、海南海桑4种植物,其他物种的Zn含量都低于0.02 g/kg(表4)。测得的12种红树植物的N/P平均值为13.12,小于14,主要受N元素限制。
2.1.3亚热带常绿阔叶林植物叶片元素丰缺状况
由表1得,亚热带常绿阔叶林16种植物叶片的P、Na与N的计量比值显著低于全国水平(P<0.001),说明其含量缺乏,如五列木、马蹄荷、阿丁枫、凤凰润楠4种植物,P的含量均不超过0.50 g/kg,甜锥、岭南槭、黄樟、青榨槭、凤凰润楠、岭南酸枣的Na含量较低,均不超过0.10 g/kg[37]。而Mn与N的计量比值显著高于全国水平(P<0.001),含量丰富,如岭南槭、拟赤杨、阿丁枫、枫香4个树种Mn的含量均大于1.00 g/kg[37]。Al与N的平均计量比远高于全国水平,但分析结果表明并未有显著差异,这是由于五列木物种有极高的Al含量,说明酸性土上的植物存在Al的富集状况。16种常绿阔叶林植物的N/P比值均大于16,平均值为26.27,其中五列木和水青冈分别高达41.66和35.52,表明南岭亚热带常绿阔叶林植物存在明显的P限制。
2.2 不同生境植物叶片元素计量关系
从相关性散点图(图2)可以看出,4个研究地点的植物叶片N与P的含量均表现出显著的正相关,作为两种重要的代谢元素,N、P在植物营养吸收方面表现出稳定的协同性;从点的分布看,常绿阔叶林的N与P含量总体低于其他生境。在Ca和Mg的关系上,喀斯特和亚热带常绿阔叶林植物均表现出显著的正相关,而红树林植物未表现出相关关系;数据双对数转化后的Ca和Mg线性回归线斜率弄岗的明显高于西双版纳的,并且弄岗的Mg含量处于高值区间。常绿阔叶林植物叶Mg含量的分布在低值范围。N与K在红树林和常绿阔叶林植物中有显著的正相关,而在喀斯特生境中均无相关关系;西双版纳喀斯特植物叶片K含量的总体分布范围低于其他生境,呈现出低K的特点。由图3看出,P与Zn在两种喀斯特森林和红树林植物中表现都表现出显著正相关关系,具有良好的协同性;从点的分布看出,弄岗喀斯特植物的Zn含量明显高于西双版纳和红树林植物。K和Mn在红树林中呈现出负相关关系,而在喀斯特森林和常绿阔叶林中未出现相关关系;亚热带常绿阔叶林植物Mn元素的分布明显高于西双版纳和红树林植物,差异显著。P与S在弄岗喀斯特森林中为正相关,而在红树林中没有相关关系。
3 讨论
3.1 不同生境植物叶片元素含量丰缺及特殊性
3.1.1喀斯特森林叶片元素分析
本研究中,西双版纳和弄岗两个地点的喀斯特森林植物叶片含有丰富的Ca、Mg含量,特别是弄岗喀斯特植物的Mg高度富集。这是由于喀斯特生境中的基岩多为碳酸盐岩类,其主要化学成分为CaCO3、MgCO3等可溶性盐类,土壤中富含的大量Ca、Mg被植物体吸收,使其普遍具有高Ca、Mg的特性。弄岗喀斯特植物叶片的Mg含量显著高于西双版纳(Anova,F=26.75,P<0.001),这可能与生境内基岩分布有关,弄岗地区含有一定的白云岩、白云质灰岩,相比石灰岩,白云岩较高的Mg含量[40]。Ca是细胞壁中间层的组分[41],在维持细胞膜的结构和功能,调节酶活性方面具有重要作用[42],Mg元素参与植物叶绿素的合成及一些与光合作用有关的酶的合成[43]。虽然Ca和Mg是植物生长的重要营养元素,但过量会对植物产生毒害作用,特别是Mg元素[44]。曾有研究表明,土壤中Ca/Mg比例小于1时,会对一些非钙土植物产生毒害,比例越低, 毒性越强[45-46]。本研究中的西双版纳绿石林地区表层土的Ca/Mg比例为3.5,5—50 cm土壤的Ca/Mg比例为1.6,弄岗喀斯特森林0—10 cm土壤的Ca/Mg比例为2.72,两地植物应该都没有受到Mg的毒害。喜钙植物能通过细胞外形成草酸盐结晶,减少Ca在原生质中富积造成细胞伤害,嫌钙植物会因Ca过量造成毒害。运用周运超[6]使用Ca+Mg对植物钙生特性进行分类的方法,发现西双版纳喀斯特植物中,油朴、鸡骨香、槟榔青、石筋草的Ca+Mg含量大于40 g/kg,为嗜钙型植物,南酸枣、藤春、坡垒、豆果榕、大果山香圆的Ca+Mg在30—40 g/kg之间,为喜钙型植物。弄岗喀斯特植物中,白饭树、清香木、野漆、红背山麻、南酸枣为嗜钙型植物,日本五月茶、构树、米仔兰、桑枝米碎、云南倒吊笔、黄梨木、闭花木、肥牛树、米扬噎、苹婆、菜豆树、灰毛浆果、山石榴为喜钙型植物,这些都对喀斯特环境有良好的适应性。
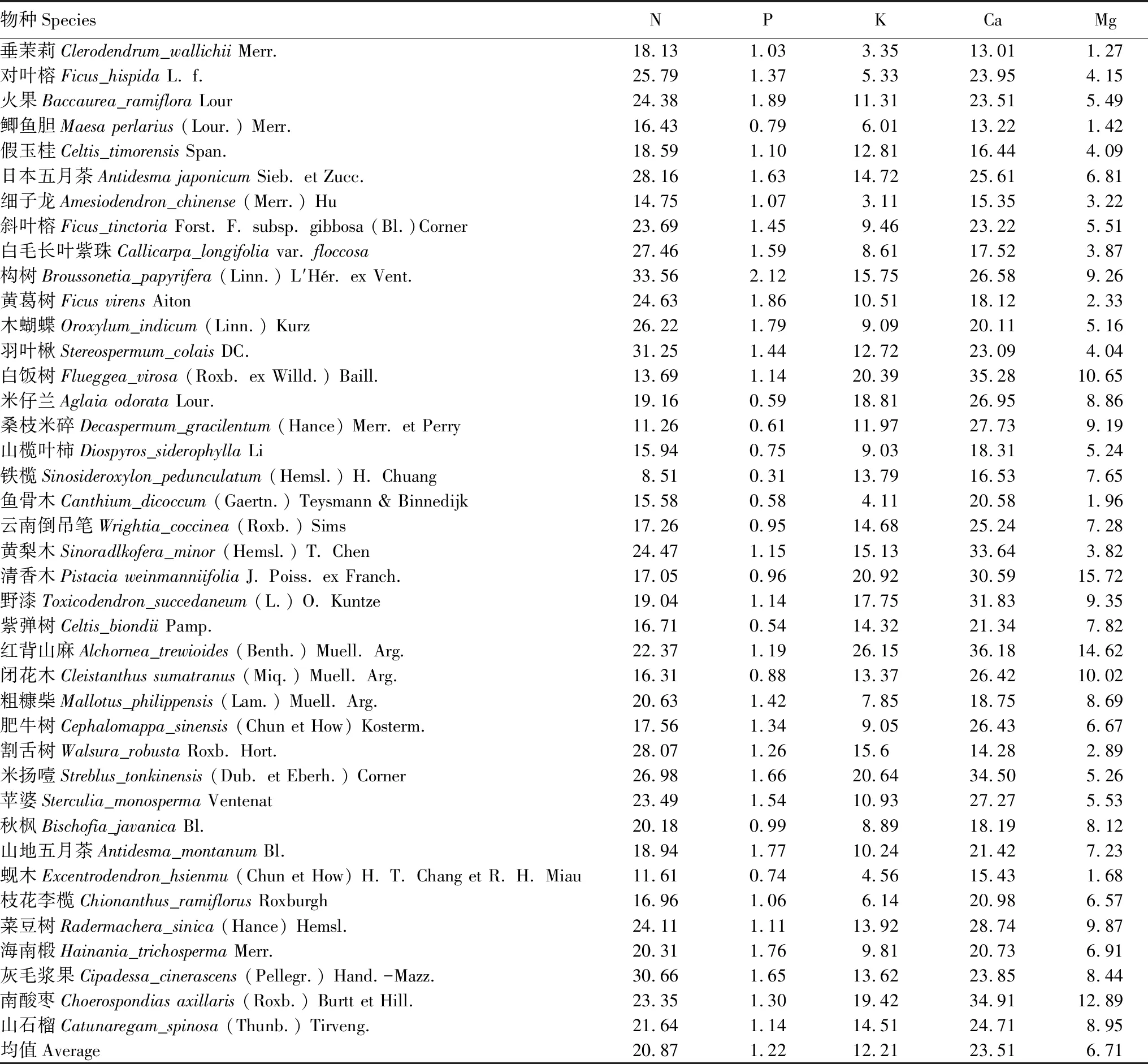
表3 弄岗喀斯特森林植物叶片元素含量(3—10个个体的平均值)和N/P比值/(g/kg)
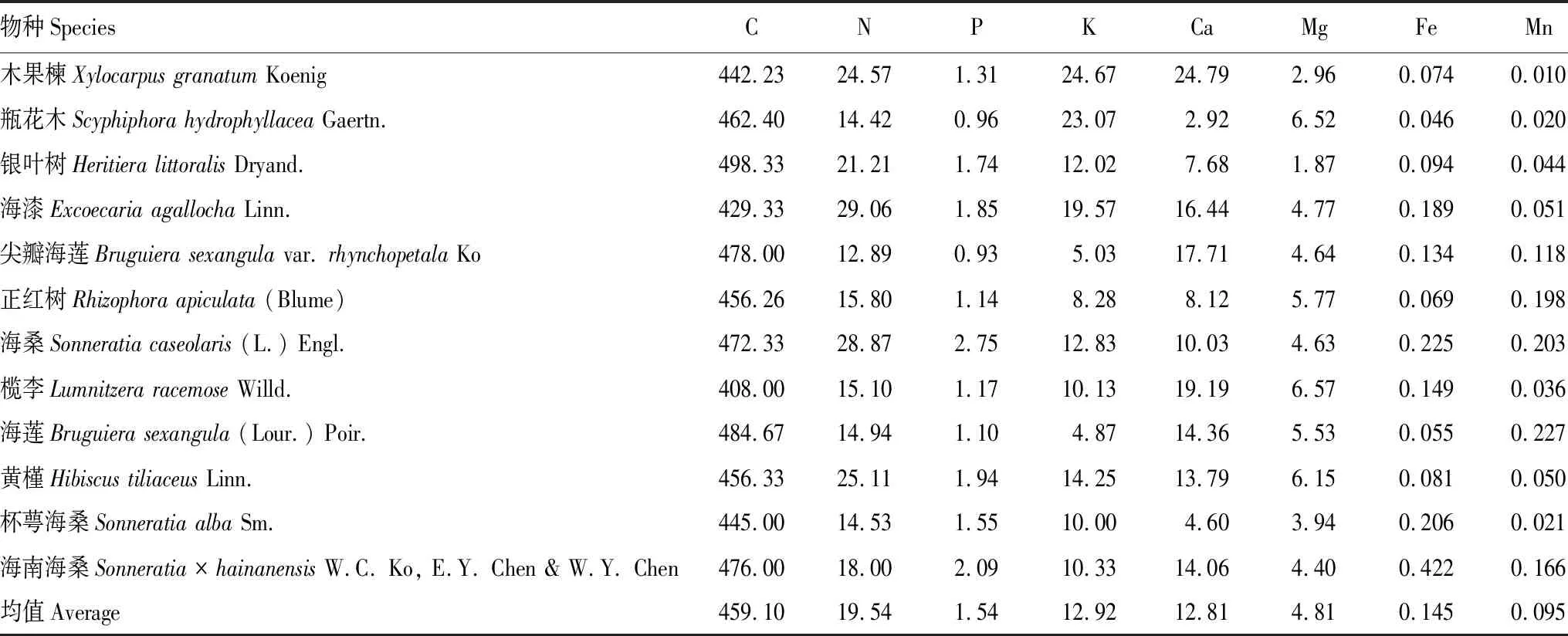
表4 海南红树林物种植物叶片元素含量(3个个体的平均值,g/kg)和N/P比值
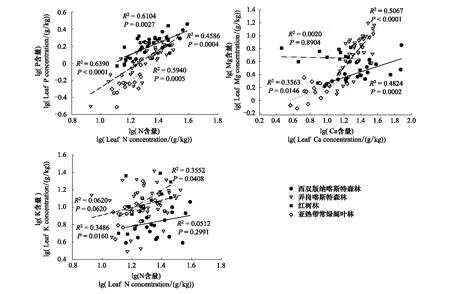
图2 不同生境植物元素含量相关散点图Fig.2 The correlation between elements in different habitats

图3 不同生境植物元素含量相关散点图Fig.3 The correlation between elements in different habitats
研究结果显示,西双版纳喀斯特植物叶片的C含量表现出明显的低量特征,这可能是因为此地喀斯特植被拥有较多的落叶树种[24]。西双版纳每年有半年的旱季,并且喀斯特土壤浅薄,持水能力差,落叶是对干旱的适应。常绿植物叶寿命长,需要投资更多碳骨架构建叶片,因而C含量比落叶树高。西双版纳植物的K和Zn含量相对缺乏,这是由于碳酸盐岩特别是石灰岩化学淋溶强烈,风化快,元素易流失,不容易留下较粗的矿物颗粒,成土速度缓慢,使得碳酸盐岩中处于痕量水平的K[47]不易通过土壤对植物进行充分的供给。同时,K+与Ca2+两个阳离子可竞争质膜上的吸收部位,在植物吸收上表现为拮抗作用,当土壤中存在丰富的Ca时会对K的吸收产生拮抗[48]。喀斯特石灰性土壤较高的pH是缺Zn的重要原因,土壤pH值是重要的土壤理化指标,已有前人研究发现土壤pH值对土壤有效Zn含量的显著影响,pH值升高会降低土壤Zn的有效性[49];Pardo等[50]对土壤pH值与土壤有效Zn吸附解吸的关系做了分析,发现土壤Zn随着pH值的上升,吸附量上升,解吸量下降;随着pH值的下降,吸附量下降,解吸量上升。而弄岗喀斯特植物的K与Zn含量显著高于西双版纳喀斯特植物,且达到富集水平,这可能与白云岩沉积过程中,常混入含K的长石[51]和含Zn的方解石矿物有关。碳酸盐岩多由海洋中所沉积的碳酸钙在压力作用下结合形成沉积岩石,这种海相沉积岩相对拥有更多的S[52],这可能导致了弄岗喀斯特植物有较高的S含量。Fe的含量在两种喀斯特生境植物中都不高,弄岗植物甚至表现出缺乏状况。这与石灰性土壤缺Fe的症状有关,土壤pH值提高会降低Fe的有效性[53],而碳酸钙与Fe也会形成难溶的化合物,影响对植物的供应[54]。
本研究中,西双版纳喀斯特森林植物的N/P平均值为14.27,介于14—16之间,受N和P的共同限制,弄岗喀斯特森林植物的N/P平均值为18.26,大于16,表明受到P限制,可以看出N和P都对喀斯特植物产生了限制作用,特别是P。Du等[21]对中国西南喀斯特地区植物叶片N/P比值的研究说明植物可能受到P的限制或N、P的共同制约,与本研究结果相同。这与喀斯特土壤中较高的Ca和pH值有关,高Ca、高pH值会导致土壤P-Ca化合物的形成和稳定性提高,使得P的有效性降低[55]。
3.1.2红树林叶片元素分析
所研究的海南清澜港红树林植物有极高的Na含量,Na/N比值超过全国水平的13倍,其平均含量高于北海11种红树植物Na含量(13.19 g/kg)[56]。红树林是生长在热带、亚热带海滨的群落类型,海水中的盐分浸渍、滞留在红树林土壤,迫使红树植物被动地吸收、积累大量的Na+。西双版纳喀斯特和常绿阔叶林植物的Na含量特别低,相比盐生境土壤,碱性石灰土和酸性土的可溶性Na盐含量少,而高Na则不利于这两种基质上的植物生长。前人研究发现[57],K/Na比可以反映植物的抗盐碱能力,有些植物通过增加K/Na比例来提高抗盐性。本研究的12种红树植物K/Na比在 0.30—25.58之间,其中以银叶树和黄槿两种半红树为高,说明这两种植物具有良好的离子调控能力,能将Na+停留在根部,减少向植物地上部分的运输。

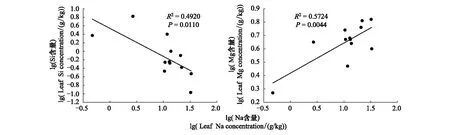
图4 红树林植物Na与Si、Mg元素相关散点图Fig.4 The correlation between Na and Si、Mg in mangrove
3.1.3亚热带常绿阔叶林叶片元素分析
该生境植物的Ca、Mg含量在不同生境中相对较低。这是因为酸性土壤低pH值提高了Mn2+、Al3+的溶解度,植物吸收大量的Mn2+、Al3+会与Ca2+、Mg2+在吸收部位发生竞争,影响物对Ca、Mg的吸收,同时高温多雨的环境也使土壤环境中Ca2+、Mg2+部分淋失。亚热带常绿阔叶林植物的N、P含量分布总体低于喀斯特和红树林植物,N/P比值为26.27,明显受到P的限制。P在酸性土壤中易与Fe、Al结合,形成不溶性的磷酸铁(FePO4)和磷酸铝(AlPO4)[63],从而减少了植物对P的吸收。
3.2 不同生境植物叶片元素之间的计量关系差异
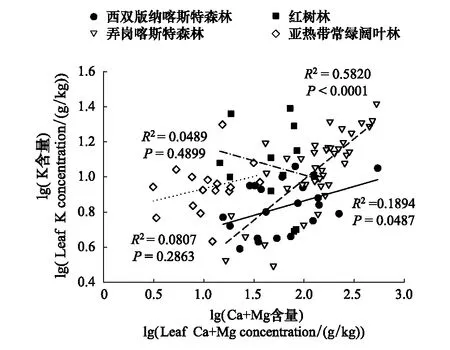
图5 不同生境植物K与Ca+Mg元素含量相关散点图 Fig.5 The correlation between K and Ca+Mg in different habitats
N和P是构成蛋白质和遗传物质的重要组成元素,对植物的光合作用和细胞生长分裂等生理活动有重要贡献,是植物生长发育的重要限制元素。Sterner和Elser[2]研究认为植物叶片的N与P含量间存在显著的正相关关系。本研究中的四个生境的植物叶片N、P含量存在一定的差异,但都表现出显著的正相关关系,这决定了不同生境植物N与P比例的变异性要小于N、P含量的变异,反映了不同基质上植物叶片N、P含量间良好的协同性,是植物适应环境的普遍规律。Ca2+和Mg2+是植物体中含量最丰富的两种二价阳离子,二者具有相似的吸收方式,从生物地球化学属性上看,Ca和Mg同为碱土元素,在化学性质上具有相似性。因此本研究中两种喀斯特森林和亚热带常绿阔叶林植物的Ca、Mg含量之间都表现出较好的协同性,但红树植物的Ca、Mg却没有相关关系。分析发现,红树植物的Mg与Na之间具有显著的正相关(R2=0.5724,P<0.01)(图4),表明高量的Na盐离子吸收可以提高Mg的吸收,某一元素的不足或过量吸收都会影响植物养分的平衡关系,所以过量的盐离子可能造成了红树植物对Ca、Mg养分的不平衡利用。N和K的计量关系在红树林和亚热带常绿阔叶林植物中表现为协同性,这与两种元素的生理功能有关,N是合成蛋白质的主要元素,而K可以提高植物对N的利用,加快转化成蛋白质的速率[64]。但是在两种喀斯特森林植物中,N与K并未表现出相关关系,这可能与碳酸盐岩母质高Ca、Mg的含量改变了植物对K元素的吸收利用有关。元素在植物生长代谢过程中要按照一定的比例分配,混合比率K/[Ca+Mg]可以表征植物体内K与Ca和Mg的互作关系[65]。所研究的两种喀斯特森林Ca+Mg的含量与K含量均为正相关(P<0.05,P<0.001),红树林和常绿阔叶林均未有相关关系(P>0.05)(图5),这说明喀斯特森林植物高Ca、Mg的吸收量协同影响了K的利用,进而打破了N与K的元素平衡关系。
在喀斯特森林和红树林植物中,P与Zn的含量都表现出协同关系,这可能与植物的代谢过程中,P和Zn都参与了酶的合成有关。Zn主要通过酶对植物代谢起作用,如铜锌超氧化歧化酶、乙醇脱氢酶、碳酸酐酶和RNA聚合酶等,需要锌的参与才能发挥其正常的生理功能[66],而酶的产生需要大量核酸(P库)的复制[67]。在K和Mn的计量关系上,只有红树植物体现出拮抗作用,而喀斯特植物和常绿阔叶林植物都没有相关关系。有相关研究表示盐分可以降低土壤的pH值,改变氧化还原电位,转化Mn的形态,同时,Na+和Ca2+的离子交换作用增加了Mn的有效性,以诱导土壤中Mn释放[68]。K、Mn之间的拮抗作用应该是高盐环境的红树植物避免Mn过量吸收的重要方式,然而在高Mn的酸性土植物中却没有这种拮抗关系,这可能与植物的遗传适应性和合适的K/Mn浓度比有关。有研究表明,S可以影响植物体内P的含量水平[69],本研究中的弄岗喀斯特森林植物S与P之间表现为正相关,喀斯特土壤高pH值可以降低土壤对S的吸附,增加植物的吸收利用,而S与P之间的协同作用可以缓解喀斯特植物缺P的症状,这种关系则在缺N高P的红树植物中未有体现。
3.3 结语
通过对比喀斯特森林、盐碱土的红树林以及酸性土的常绿阔叶林植物叶片的养分含量,并且将每种生境植物元素计量比与全国植物和陆生高等植物的元素数据进行比较,发现不同生境基质上植物的元素吸收、利用方式有明显差异,特定的生境植物富集不同的元素。通过元素之间的关系分析发现,不同生境植物叶片的元素计量关系发生了改变,是植物与环境长期适应的结果。研究可为多种森林生态系统植被恢复和生物地球化学循环提供重要参考。
猜你喜欢
儿童故事画报·自然探秘(2022年3期)2022-04-27 00:17:10
落叶果树(2021年6期)2021-02-12 01:29:10
阅读(低年级)(2020年8期)2020-11-06 06:23:19
乡村地理(2019年2期)2019-11-16 08:49:32
————水溶蚀岩石的奇观
家教世界(2019年4期)2019-02-26 13:44:20
疯狂英语·初中天地(2018年6期)2018-11-24 02:39:24
歌海(2018年4期)2018-05-14 12:46:15
体育科技(2016年2期)2016-02-28 17:06:07
西华师范大学学报(自然科学版)(2015年3期)2015-02-27 15:31:17
文化月刊·下旬刊(2014年6期)2014-08-28 00:58:59
