
陆地棉种子萌发期对低温胁迫的响应及耐冷性鉴定
2021-01-14 03:02张陇艳程功敏魏恒玲王寒涛芦建华马峙英喻树迅
中国农业科学 2021年1期
张陇艳,程功敏,魏恒玲,王寒涛,芦建华,马峙英,喻树迅✉
1河北农业大学/华北作物改良与调控国家重点实验室,河北保定 071001;2中国农业科学院棉花研究所/棉花生物学国家重点实验室,河南安阳 455000
0 引言
【研究意义】棉花(Gossypium hirsutumL.)原产于热带和亚热带地区,低温冷害是限制其品种产区分布的重要环境因素[1]。中国的棉花产区主要分布于长江流域、黄河流域和西北内陆地区[2]。由于粮棉争地矛盾日益加深,棉花种植区域逐步向西北内陆等瘠寒地区拓展[3]。低温冷害是棉花西进和北上拓展所面临的重要非生物胁迫之一,在播种出苗阶段,低温冷害轻则延迟种子萌发和幼苗发育,重则降低出苗率和幼苗的存活率[4-6]。因此,鉴定和筛选吸胀萌发期耐冷品种、探究棉花吸胀萌发期耐冷性机理,可以为耐冷性遗传改良,以及棉花生产区域拓展提供重要依据。【前人研究进展】棉花为双子叶植物,种子包含有胚乳层,萌发首先从种子吸胀开始[7]。双子叶植物种子萌发需要完成种皮破裂(testa rupture)和胚乳层破裂(endosperm rupture)2个物理过程,胚根从破裂的珠孔胚乳层处出现标志着萌发的结束,拟南芥(Arabidopsis thaliana)萌发阶段分别以种皮破裂和胚乳层破裂作为前期和后期萌发结束的标志[8-9]。在一些研究中,棉花种子萌发期的非生物胁迫鉴定通常还包括胚根突破胚乳层到胚芽伸长这个阶段。植物萌发期和幼苗期面临的低温胁迫通常是有别于冻害温度(<0℃)的非结冰温度(0—15℃)[10];棉花萌发最低温度为10—12℃,最适温度为 20—30℃[3,11-12]。针对植物萌发期耐冷性鉴定,国内外研究人员从生理生化和形态指标等方面进行广泛的研究。陈昊等[13]采用吸水速率、相对吸水量和相对发芽率等指标对 64份花生材料进行吸胀萌发阶段的耐冷性鉴定,发现冷胁迫能降低吸胀速率,吸胀阶段冷处理 12 h可以区分不同种质间抗冷性差异;李霞等[14]通过测定吸胀阶段的吸水速率、电导率、脱氢酶和α-淀粉酶活性及发芽指标研究不同耐冷基因型玉米对低温的响应,发现冷敏感玉米籽粒吸胀迅速且电导率相对增大,而耐冷性吸胀缓慢。HUANG等[10]利用萌发率和胚芽数指标来鉴定 125份玉米自交系的萌发期耐冷性,发现萌发期指标具有显著正相关(R=0.41),而萌发期和幼苗期指标之间相关性不显著(-0.05≤R≤0.16)。SCHLÄPPI等[15]采用低温发芽力和低温胚芽生长速率指标对 202份水稻品种萌发期进行耐冷性鉴定,发现萌发期指标彼此相关性较高,而其与幼苗期耐冷指标不相关或相关性较弱。以上研究表明,在作物的不同生育时期,其耐冷性存在差异。在棉花耐冷性形态指标鉴定方面,高利英[16]利用发芽率、发芽指数和活力指数等萌发期指标对38份陆地棉材料进行耐冷性鉴定和综合评价;王俊娟[17]利用低温萌发后的子叶平展率作为萌发期低温鉴定的指标对 13份陆地棉品种进行耐冷性鉴定。在棉花耐冷性生理生化指标鉴定方面,武辉等[18]测定了低温胁迫下15份陆地棉材料幼苗叶片的光合参数及叶绿素、可溶性糖、丙二醛和游离脯氨酸含量以及相对电导率等12项生理指标,采用隶属函数法进行耐冷性综合评价;李志博等[19]测定了 4份耐冷性不同的北疆主栽陆地棉品种低温胁迫下的超氧化物歧化酶(superoxide dismutase,SOD)、过氧化物酶(peroxidase,POD)、过氧化氢酶(catalase,CAT)酶活性及丙二醛(malondialdehyde,MDA)、可溶性蛋白、可溶性糖含量,通过回归分析发现相对CAT酶活性、相对MDA和可溶性蛋白含量与棉花幼苗耐冷性密切相关;夏军等[20]发现耐冷棉花材料中SOD、POD和CAT酶活性在低温胁迫下比冷敏感材料的要高;高利英等[21]发现在不同冷处理水平下耐冷棉花中SOD和POD酶活性均能保持较高水平,而在耐冷材料中CAT酶活性会逐渐超越冷敏感品种。棉花萌发期耐冷性是复杂的综合性状,仅依靠单一的指标很难筛选到抗冷性综合性能强的材料。因此,近年来基于因子分析和主成分分析的隶属函数综合评价方法在棉花萌发期非生物胁迫鉴定方面得到了广泛的应用,这对棉花吸胀萌发阶段的耐冷性鉴定提供了很大的帮助[22-24]。【本研究切入点】尽管前人对棉花萌发期的耐冷性多有研究,但是由于试验材料、鉴定指标和鉴定方法的局限性而不能对吸胀萌发阶段的耐冷性有全面和深入的认识。因此,利用综合评价方法系统考察吸胀期和萌发期的耐冷性关系,并且从生理和分子水平对综合评价所得的不同抗冷基因型材料进行研究,必然会为深入理解棉花吸胀萌发期耐冷性机理和遗传改良提供帮助。【拟解决的关键问题】本研究通过对主要来自黄河流域和西北内陆棉区的53份陆地棉材料进行室内低温萌发鉴定,综合评价材料的吸胀萌发期耐冷性。研究差异材料低温胁迫处理下抗氧化物酶活性、渗透调节物质浓度及抗氧化物酶基因表达规律,明确之前未知的一些供试材料的耐冷特性,为深入解析棉花抗冷机理与鉴定耐冷种质以及育种提供理论基础。
1 材料与方法
1.1 试验材料
利用56份陆地棉常规栽培品种(系)作为试验材料,所有材料均由中国农业科学院棉花研究所早熟育种实验室提供。其中,用于萌发期耐冷性鉴定的 53份材料包括黄河流域材料28份、长江流域材料2份和西北内陆棉区材料23份,材料名称及详细信息见电子附表1。
1.2 试验方法
每个材料选取饱满种子100粒,3个重复,称重后在4℃和25℃条件下分别吸胀0和6 h,拭去种子表面水分后分别再次称重,计算不同温度下的吸胀速率。其中,中50、中425和中915均吸胀12 h,每小时称重一次,参考前人方法计算相对吸水量和吸水速率[13]。从每个品种(系)中挑选400粒饱满种子,清水冲洗干净后置于25℃浸泡12 h。挑选已完成吸胀的种子(外种壳的珠孔处开裂并露出白色胚乳层)各100粒,拭去其表面水分并称重,然后均匀地平铺到垫有双层滤纸的培养皿中,设3个重复。置入12℃冷光源培养箱内黑暗萌发12 d。每天准时统计萌发种子数,萌发以胚根顶破珠孔处的胚乳层并伸出 2—3 mm为标准。低温萌发12 d后,人工将种胚的芽和胚用镊子分离并分别称鲜重,烘干后分部称干重,得到低温萌发后芽和胚的鲜重及干重。
分别从新陆中4号和中103026中挑选200粒饱满的种子,25℃清水浸泡12 h后冲洗3—5遍,挑选露白种子各50粒均匀平铺到垫有双层滤纸的培养皿中,置于25℃冷光源培养箱内暗培养24 h。分别从每个材料的50粒种子中挑选30粒萌发一致的种子,将种子均匀平铺到垫有双层滤纸的培养皿中,置于 4℃冷光源培养箱内暗培养24 h。在低温处理0、3、6、12和24 h时分别取出3粒种子,去除外种壳及胚乳层后投入液氮冷冻保存,另外在0、12和24 h时分别取出3粒种子用于生理指标测定。
1.3 指标测定与方法
1.3.1 形态指标 根据吸胀前后及萌发前后的种子重量,以及每天统计的各品种(系)的萌发种子数,利用以下公式或方法计算各个指标值:
种子吸胀速率(imbibition rate,IR):IR=(B-A)/A×100%,式中,A、B分别为吸水前和吸水后的种子质量[25];
相对吸胀速率(relative imbibition rate,RIR):RIR=低温吸胀速率/常温吸胀速率;
物质增长率(material growth rate,MGR):MGR=(M2-M1)/M1。式中M1、M2分别为吸胀结束后百粒鲜重和萌发结束后百粒鲜重;
物质效率(productive efficiency,PE):PE=m2/(m1-m2)。式中,m1、m2分别为吸胀前百粒重和萌发结束后百粒重;
发芽率(germination rate,GR):GR=发芽种子数/供试种子数×100%。试验中选择低温萌发第7天统计种子发芽率[22];
发芽势(germination potential,GP):GP=发芽达到高峰期时发芽种子数/供试种子数×100%,本试验中选择低温萌发第5天调查种子发芽势[26];
发芽指数(germination index,GI):GI=∑(Gt/Dt)。式中Gt为第t天的发芽种子数,Dt为至t天的发芽天数(d)[22];
活力指数(vigor index,VI):VI=S×GI。式中S为萌发种子鲜质量,GI为发芽指数[22];
平均发芽时间(mean germination time,MGT):MGT=∑(Gt×Tt)/∑Gt。式中,Gt为在不同时间的发芽数,Tt为发芽日数[27];
平均发芽速度(average germination speed,AGS):AGS=(∑Dn)/∑n。D为种子放置到培养皿后算起的天数,n为相应各天发芽种子数[28];
种子萌发指数(seed germination coefficient,SGC):SGC=1.00×nd2+0.75×nd4+0.50×nd6+0.25×nd8。nd2、nd4、nd6、nd8分别指第2、4、6和8天的种子发芽率[26]。
以上指标涉及重量的测定,主要用到ME104E电子分析天平(上海梅特勒-托利多)及JY2002电子天平(上海衡平)。
1.3.2 生理指标 将4℃处理0、12和24 h的去壳萌发种子样品按照苏州科铭生物技术有限公司提供的试剂盒说明进行预处理,测定SOD、POD和CAT 3种抗氧化物酶活性,同时检测样品中可溶性蛋白含量。
1.4 数据处理和分析
采用Microsoft Excel 2007软件进行数据整理,利用SPSS19.0软件进行方差分析、因子分析、相关性分析和主成分分析,运用R语言进行聚类分析,图片利用Excel和R软件绘制。使用模糊隶属函数法评价各材料种子萌发期耐冷性的差异,各个值的计算方法分别为:
隶属函数值:μ(Xi)=(Xi-Xmin)/(Xmax-Xmin),i=1,2,...,n。式中,Xi为第i个综合指标,Xmin为第i个综合指标的最小值,Xmax为第i个综合指标的最大值[29]。
综合指标权重:wi=pi/∑pi,i=1,2,...,n。式中,wi为第i个综合指标在所有综合指标中的权重,pi为各品种第i个综合指标的贡献率[18]。
综合评价值:D=∑[μ(Xi)×wi],i=1,2,...,n。式中,D值为各材料在冷害胁迫条件下用综合指标计算所得的耐冷性综合评价值[18]。
1.5 抗氧化物酶基因的表达分析
1.5.1 种子RNA的提取和检测 将4℃处理0、3、6、12和24 h的去壳萌发种子样品按照RNApre Pure多糖多酚植物总RNA提取试剂盒(TIANGEN,北京)说明方法提取胚根和子叶中的总RNA,利用1% agarose凝胶检测其完整性,120 V电泳20 min。
1.5.2 cDNA的合成 利用 TaKaRa的 PrimeScript™RT reagent Kit with gDNA Eraser试剂盒说明书进行(宝生物,大连)反转录。该试剂盒反应分2步进行:去除RNA样品中残留的基因组DNA(gDNA);cDNA单链合成(所有操作都需在冰上进行)。将反转录后的cDNA放置到-20℃保存备用。
1.5.3 荧光定量PCR及数据分析 利用Oligo7软件设计内参基因和抗氧化物酶基因的荧光定量引物。qRT-PCR反应在Applied Biosystems 7500仪器中进行,体系采用TaKaRa的SYBR Premix Ex Taq(2×)试剂盒。反应结束后,将数据导出,使用 2-ΔΔCt方法计算基因的相对表达量。
2 结果
2.1 低温冷害对棉花种子吸胀的影响
为了研究低温冷害对棉花种子萌发期吸胀阶段的影响,分别测定3份陆地棉栽培品种在常温(25℃)和低温(4℃)条件下0—12 h的相对吸水量和吸水速率(图1)。与常温相比,中50、中425和中915在低温下吸胀12 h后相对吸水量分别只达到常温的76.44%、78.93%和 71.90%。常温条件下,中 50、中425和中915分别在3、2和4 h达到最大吸水速率(分别达到1.1408、1.5156和0.8566 g·h-1);低温条件下,其最大吸水速率分别出现在7、2和4 h时,仅达到0.5367、0.8041和0.5049 g·h-1。说明低温能够显著降低棉花种子萌发期吸胀阶段的相对吸水量和吸水速率。
2.2 棉花萌发期耐冷性指标的差异
常温(25℃)条件下的萌发预试验发现,完成吸胀后所有材料的露白种子(外种壳珠孔处开裂并露出白色胚乳层)在一天之内均可悉数萌发,说明选择露白种子进行萌发试验能够消除材料之间除温度因素以外的所有差异,因此,低温下相关萌发指标就能直接地反映不同材料的耐冷性强弱。这些指标中,低温种子吸胀速率和相对吸胀速率被用来反映棉花吸胀阶段的耐冷性,低温物质增长率、物质效率、发芽指数、活力指数、平均发芽时间、平均发芽速度、发芽势、发芽率、种子萌发指数、芽鲜重、芽干重、胚鲜重、胚干重被用来反映萌发阶段耐冷性。从表1可以看出同指标在不同材料间存在较大变幅,其中低温吸胀速率变化幅度为 0.07%—0.62%,低温相对吸胀速率变化幅度为 0.24%—1.00%,芽鲜重变化幅度为0.31%—4.06%,胚鲜重变化幅度为 13.11%—24.17%,芽干重变化幅度为0.09%—0.65%,胚干重变化幅度为5.89%—11.93%,物质增长率变化幅度为 0.01%—0.24%,物质效率变化幅度为0.04%—0.35%,平均发芽速度变化幅度为5.10%—10.55%,发芽指数变化幅度为2.34%—19.99%,活力指数变化幅度为 0.86%—78.31%,平均发芽时间变化幅度为4.99%—10.18%,发芽势变化幅度为 0.00—0.82%,发芽率变化幅度为0.03%—1.00%,种子萌发指数变化幅度为0.07%—1.72%。53份陆地棉材料间的各个萌发期耐冷指标的变异系数也存在差异,其中,发芽势(CV=1.235)和活力指数(CV=0.551)的变异系数最高,而胚鲜重(CV=0.130)和胚干重(CV=0.141)的变异系数最低,说明各个耐冷指标对材料基因型的依赖性不同。由于各个萌发期耐冷指标反映的信息可能存在重叠,因此,有必要进行指标间的相关性分析。
2.3 主成分分析

表1 不同棉花材料萌发期各耐冷指标的差异Table 1 Differences in chilling-tolerance indices of different cottons at germination stage

表2 萌发期各综合指标的系数及贡献率Table 2 Coefficients and contribution of comprehensive indexes at germination stage
主成分分析可以将多维变量简化为少数几个相互独立的综合指标。15个萌发期单项耐冷指标的主成分分析结果见表2,以特征值大于1为阈值共得到3个主成分,对应的特征根分别为2.877、1.656和1.148,其贡献率分别为55.17%、18.27%和8.79%,累计贡献率达到82.23%,说明这3个主成分能够解释绝大部分信息,而其余成分最大贡献率仅为6.723%,可忽略不计。这样就将原来的15个单项指标转化为3个独立的综合指标,命名为CI1、CI2和CI3,而这 3项综合指标代表了原有单项指标的耐冷信息。其中,第一主成分CI1的特征向量主要描述的是活力指数(0.335)、低温萌发指数(0.333)、平均发芽速度(-0.333)、发芽指数(0.331)、平均发芽时间(-0.330)、发芽率(0.321)、芽鲜重(0.315)、芽干重(0.284)、物质增长率(0.258)和发芽势(0.238);第二主成分CI2的特征向量主要描述的是胚干重(0.516)和胚鲜重(0.490);第三主成分CI3的特征向量主要描述的是低温吸胀速率(0.481)、相对吸胀速率(0.632)和物质效率(-0.553)。
2.4 品种综合评价
2.4.1 隶属函数分析 按照隶属函数计算公式求得各品种综合指标的隶属函数值(表 3)。对于同一综合指标如CI1来说,棉花萌发期的各品种在低温条件下,新陆中 4号的隶属函数值μ(X1)最大(1.000),说明在此综合指标下其萌发期耐冷性最强;中103026的隶属函数值μ(X1)最小(0.000),说明在此综合指标下其萌发期耐冷性在最差。对于综合指标CI3来说,中 679的隶属函数值μ(X1)最大(1.000),说明在此综合指标下其萌发期耐冷性最强;品种川01的隶属函数值μ(X1)最小(0.000),说明在此综合指标下其萌发期耐冷性最差。根据综合指标权重公式以及各综合指标的贡献率可计算得到3个综合指标的权重,分别为6.532、4.427和2.641(表3)。
2.4.2 综合评价及聚类分析 按照综合评价指标公式计算各棉花品种(系)萌发期耐冷性综合指标值(表3),并根据求得的综合评价指标D值对53份材料的耐冷性进行排序,采用最大距离法对综合指标进行聚类分析,依据D值大小及聚类结果可将材料划分为4类(图2)。新陆中4号、中棉所14等5份材料为第一类(Ⅰ),占供试材料总数的 9.43%,属于低温萌发强耐冷型。新陆中20号、黑山棉1号等13份材料为第二类(Ⅱ),占材料总数的24.53%,属于低温萌发耐冷型。新陆中14号、邯2490、新陆中60号等26份材料为第三类(Ⅲ),占总材料的49.05%,属于低温萌发不耐冷型。新陆中34号、新陆中28号、川01等9份材料为第四类(Ⅳ),占材料总数的16.98%,属于低温萌发冷敏感型。从比例来看,耐冷品种(系)比较少,说明获得棉花萌发期耐冷性种质的来源途径比较狭窄且多数材料适合在更温暖的季节播种。在这53份陆地棉材料中,新陆中4号的萌发期耐冷综合指标值最高(D=0.938),而中103026的萌发期耐冷综合指标值最低(D=0.166),因此,选择这两份萌发期耐冷性存在显著差异的材料用于后续生理水平的研究。
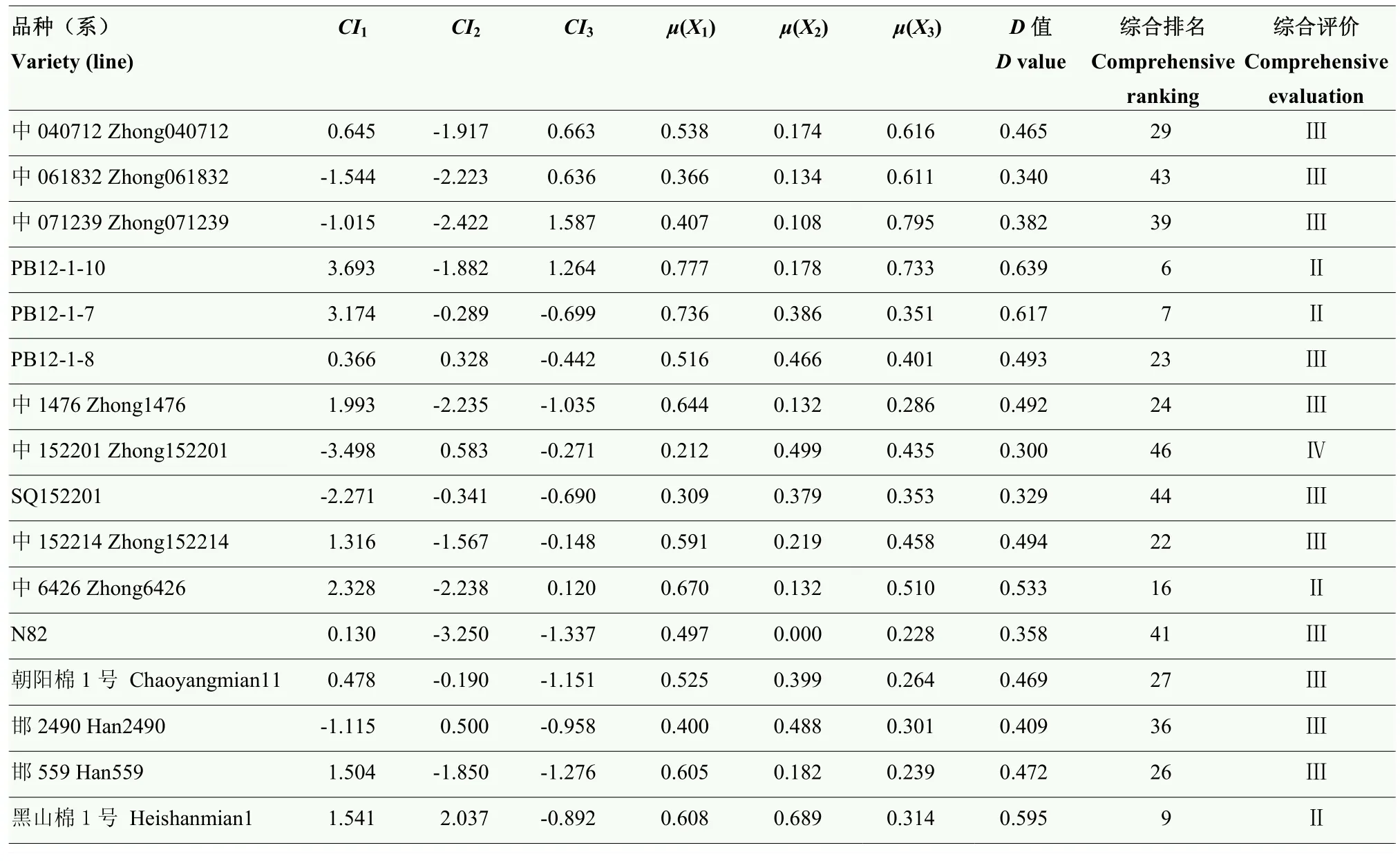
表3 各品种的综合指标值、权重、μ(X)、D值及综合评价Table 3 Value of each comprehensive index (CI), index weight, μ(X), and comprehensive evaluation value (D value)
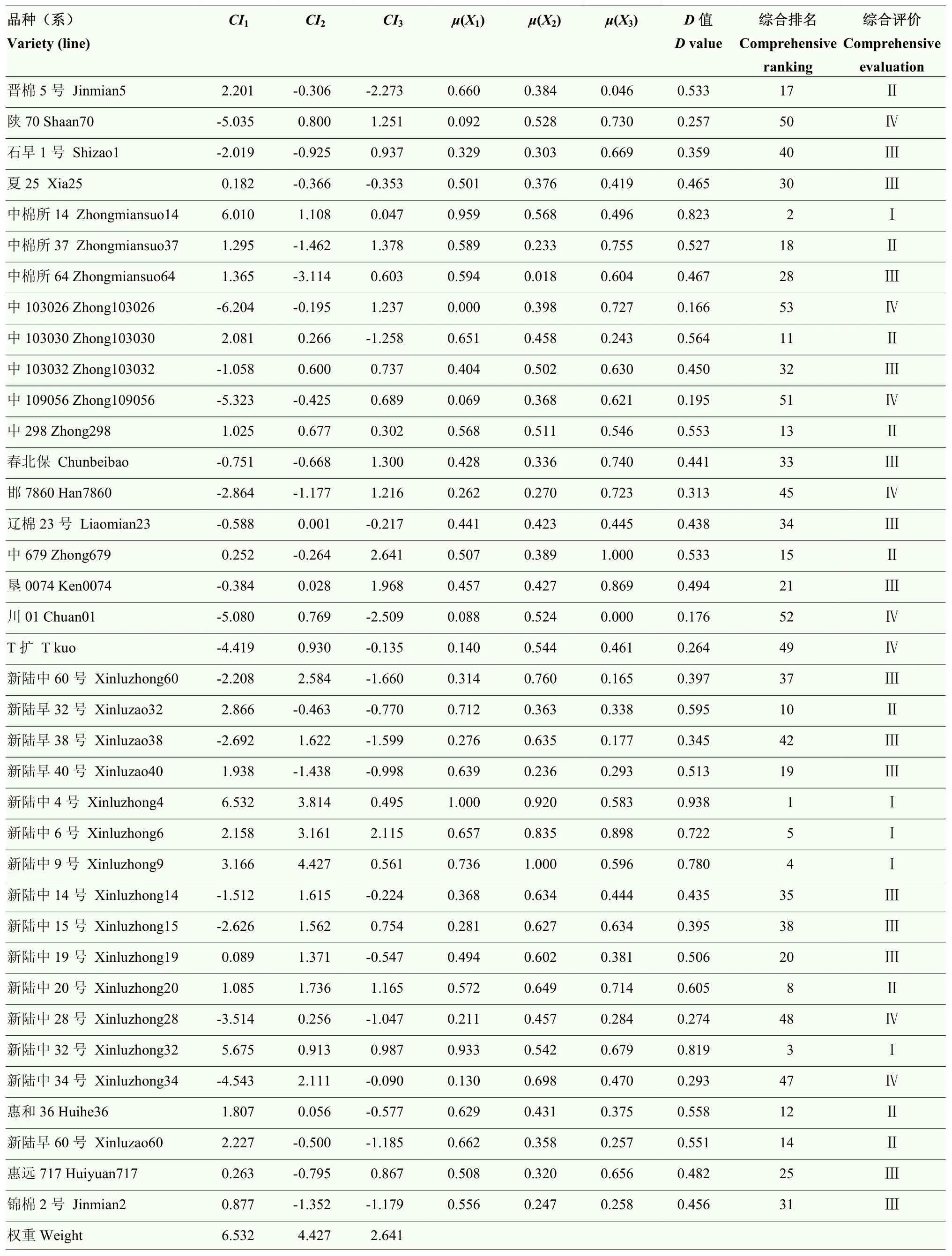
续表3 Continued table 3
2.5 棉花萌发期耐冷性指标的相关性分析
萌发期耐冷性指标间的相关性如图3所示,低温萌发后胚鲜重、胚干重、芽干重、芽鲜重、活力指数间呈极显著正相关,其中,胚鲜重与胚干重间(R=0.96)、芽干重与芽鲜重间(R=0.94)、芽鲜重与活力指数间(R=0.96)相关性都较高。芽干重、芽鲜重、活力指数、发芽势、物质增长率、发芽率、发芽指数、种子萌发指数间呈极显著正相关,其中除芽干重、芽鲜重、活力指数间相关性较高外,发芽率、发芽指数、种子萌发指数间相关性也较高(R=0.94—0.96),它们与平均发芽速度和平均发芽时间呈极显著负相关(R=-0.98—-0.67)。种子低温吸胀速率与低温相对吸胀速率间呈极显著正相关(R=0.58),它们与其他萌发指标间的相关性不显著或负相关。这说明萌发期的吸胀阶段与其他阶段可能具有不同的耐冷机制,而萌发阶段的指标所反映的耐冷信息具有重叠性。耐冷性综合评价指标(D值)与胚鲜重、胚干重、芽干重、芽鲜重、活力指数、发芽势、物质增长率、发芽率、发芽指数、种子萌发指数间均呈极显著正相关(R=0.60—0.95),其中,与芽干重、芽鲜重、活力指数之间的相关性最强(分别为R=0.88、R=0.92和R=0.95),与平均发芽速度、平均发芽时间之间呈极显著负相关(R=0.83)。这说明芽干重、芽鲜重、活力指数、平均发芽速度、平均发芽时间能较好反映不同棉花材料萌发期的耐冷性强弱。
2.6 生理指标分析
由图4可知,在对2个棉花材料的萌发期种子进行冷胁迫处理后,耐冷材料新陆中4号在处理12 h时的SOD、POD和CAT酶活性均降到最低,而在胁迫24 h时又有所上升。对于冷敏感材料来说,SOD、POD和CAT酶活性均呈现先上升后下降的趋势,并且在处理前和处理24 h时均显著地低于耐冷材料的酶活性。冷处理12 h后,冷敏感材料中的SOD和CAT 2种抗氧化物酶活性均显著高于耐冷材料,而POD酶活性在两种材料间差异不显著。这说明棉花在萌发期进行低温胁迫后,耐冷材料种子的SOD、POD和CAT酶活性能够在短时间内恢复到接近对照的水平或超过对照,这些特性均有利于耐冷材料萌发期的种子在面临冷胁迫时通过有效地降低活性氧积累对机体造成的伤害,因此,耐冷材料表现出比冷敏感材料更强的耐低温萌发能力。对于可溶性蛋白浓度而言,耐冷材料的可溶性蛋白浓度始终显著高于冷敏感材料,并且可溶性蛋白的含量均呈增加趋势。这说明耐冷材料在萌发期能维持较高的可溶性蛋白浓度,在受到低温胁迫后不同耐冷性材料的种子都会借助增加可溶性蛋白浓度而增强其自身抵御低温伤害的能力。
2.7 抗氧化物酶基因的表达量分析
由图5可知,萌发期的种子受到低温胁迫12 h后,SOD酶基因GhCSD1在两材料子叶中均下调表达,低温胁迫24 h时在耐冷材料中表达量迅速上调,而在冷敏感材料中保持持续下调趋势,且在耐冷材料中的表达量均显著地高于冷敏感材料,这与SOD酶活性的变化基本一致。POD酶基因GhPrx53在低温胁迫0—24 h,在耐冷材料中的表达量始终高于冷敏感材料,这与过氧化物酶的活性变化相一致。在胚根遭受低温胁迫0—12 h,GhCSD1在两材料中的表达量均维持在较低水平,但是在耐冷材料中其表达量始终高于冷敏感材料;冷胁迫24 h时,在冷敏感材料中的表达量显著地高于耐冷材料,说明在种子的子叶和胚根中GhCSD1的表达趋势有所差异,而POD酶基因GhPrx53的表达趋势与子叶中的基本一致。在低温胁迫前后,POD酶基因的表达量在耐冷材料的子叶和胚根中的表达量都远高于冷敏感性材料子叶和胚根中的表达量,说明两材料萌发期的耐冷性很可能与二者自身间的遗传背景差异有关。
3 讨论
3.1 低温冷害对棉花种子萌发期相关性状的影响
温度是影响棉花种子萌发的重要环境因素,温度过低会严重影响种子的萌发率和出苗率。研究表明,棉花种子萌发的最低温度为 10—12℃,温度低于 10℃时棉花种子不能萌发[12,30]。不同温度条件下材料间种子萌发鉴定结果可能会不同,说明不同温度影响种子萌发的机制存在差异[12]。植物的耐冷性是一种受多基因调控的复杂数量性状[31]。冷胁迫下不同棉花品种在种子萌发期的表现存在差异[32],遗传背景的差异往往决定种子萌发期的耐冷性。棉花萌发期的耐冷机制的研究首先依赖于代表性耐冷指标和不同耐冷性材料的筛选。为了准确反映不同棉花品种的萌发期耐冷性,普遍采用基于对照的低温相对指标[30]。本研究利用完成吸胀的露白种子进行萌发试验,它们在常温下24 h内均可以正常萌发。因此利用露白种子进行低温萌发试验可以消除材料本身其他因素的影响,从而直接反映出低温下不同材料的萌发期耐冷性。前人研究中,发芽势、发芽率、发芽指数、活力指数、萌发指数等常作为鉴定萌发性强弱的指标[33-34]。本研究发现,综合指标除与这些指标极显著正相关外,与低温萌发后胚芽鲜干重呈极显著正相关,与平均发芽时间和发芽速度呈极显著负相关,这些指标在前人的研究中还鲜见报道。这说明胚芽鲜干重可以与其他常见萌发指标一起作为耐冷鉴定的正向指标,而平均发芽时间和平均发芽速度可以作为萌发期耐冷鉴定的负向指标。
3.2 棉花萌发期与吸胀期耐冷性的比较
棉花的不同材料在不同生长发育阶段的耐冷性往往存在差异,说明棉花在不同生育阶段应对低温胁迫的机制不同。棉花春播后经常面临突发的低温天气,这对种子吸胀和萌发都会产生影响。研究发现,低温能够显著降低种子的相对吸水量和吸水速度;低温吸胀速率与低温相对吸胀速率呈极显著正相关(R=0.58,P<0.001),说明低温下的吸胀速率能够部分地反映种子吸胀阶段的耐冷性。低温吸胀速率和低温相对吸胀速率作为吸胀阶段的2个耐冷鉴定指标与萌发期的耐冷鉴定指标间不相关或呈较弱的负相关性,这可能是由于吸胀阶段的耐冷性和萌发阶段的耐冷性调控机制不同所致。王俊娟等[30]发现棉花萌发期和芽期的耐冷性存在差异,芽期比萌发期对低温冷害更敏感。黄贺等研究发现,不同甘蓝型油菜萌发期耐冷性与成苗期耐冷性不相关[35]。HUANG等[10]发现玉米不同发育时期内部的指标间相关性较强,而不同发育时期间耐冷指标的相关性不显著。在本研究中,即便是萌发期,早期的吸胀阶段与萌发期其他指标也呈现出不相关或较弱的相关性,而萌发期指标与芽期的有关指标(比如胚、芽鲜干重)则呈显著相关,这些都暗示着棉花萌发期耐冷性的复杂性。
3.3 低温冷害对棉花萌发期种子生理生化的影响
在低温胁迫下,植物在遭受伤害时其内部发生一系列变化,比如活性氧产生速度加快、膜脂氧化加速、可溶性蛋白含量积累等等,而植物体内会通过形成SOD、POD、CAT等抗氧化酶来降低胁迫对植物产生的伤害。本研究表明,在4℃低温处理12 h时,SOD、POD和CAT酶活性在耐冷材料中达到最低,胁迫后24 h时酶活性又上升到接近对照;冷敏感材料中的酶活性在胁迫后均呈先上升后下降的趋势,且在胁迫24 h时显著地低于耐冷性材料,超氧化物歧化酶基因与过氧化物酶基因的表达量变化与酶活性变化的趋势基本一致,说明SOD和POD酶活性与其在低温诱导下的基因表达关联性较高,耐冷材料的抗氧化能力比冷敏感材料更强。王冀川等[36]通过研究低温冷害对海岛棉幼苗生理生化特性的影响发现,SOD和POD是清除棉花体内活性氧以减轻膜脂伤害的重要保护酶类,这与耐冷材料中SOD和POD的酶活性与其相关基因表达量均显著高于对照及冷敏感材料的结果相一致。两材料中可溶性蛋白浓度均高于对照,且在耐冷材料中的浓度显著高于冷敏感材料,说明可溶性蛋白浓度与棉花种子萌发期耐冷性相关。姜艳丽等也有类似的研究报道,可溶性蛋白能够降低植物细胞因冷害而受伤致死的机会,从而提高植物的抗寒性[3]。综上所述,在低温胁迫下新陆中4号比中103026能够更有效地清除活性氧,其可溶性蛋白浓度相对也更高,抗氧化能力更强,拥有较高的耐低温萌发性,是研究棉花萌发期耐冷性的重要候选材料。
3.4 棉花萌发期耐冷性鉴定及品种耐冷性比较
通过对种子萌发期耐冷性综合评价值进行聚类分析,将53份棉花供试材料分为强耐冷、耐冷、不耐冷和冷敏感4个类型。其中,新陆中4号耐冷性最强而中103026耐冷性最差。在本研究中,新陆中14号萌发期种子不耐冷,其亲本中棉所19在苗期抗冷性较差[37],表明新陆中14号可能在发育的早期(萌发幼苗期)表现出不耐冷性。贝尔诺斯的2个衍生品种,新陆中19和新陆早 31分别在萌发期和幼苗期同时表现出不耐冷特性[38],这暗示贝尔诺斯可能具有不耐冷的遗传背景,而其耐冷性有待进一步研究。邯559属于萌发期不耐冷型,而其亲本邯郸109表现子叶期抗冷性[30];石早 1号的萌发期种子不耐冷,其亲本双价抗虫棉SGK321据报道在幼苗期抗冷性较强[39],这可能是育种导致耐冷信息丢失或生育时期不同所致。在耐冷品种中,新陆早 60与李星星等[40]在萌发期鉴定的耐冷性结果一致,说明该品种具有稳定的萌发期耐冷性。根据综合指标可以看到,新陆中4号、新陆中32号、新陆中9号、新陆中6号为萌发期强耐冷品种,新陆中20号、新陆中60号为萌发期耐冷品种,以上6个品种的早熟性综合指标值均较小(相对早熟性较差)。而姜艳丽等[3]通过对6个不同熟性的陆地棉品种幼苗进行4℃处理,发现早熟品种中50和N52的耐冷性最强;尹晓斐[41]通过进一步分析这6个品种二叶期幼苗4℃胁迫下的超氧化物歧化酶基因(CSD2a)表达量,得出与之前相同的结论。本研究表明处理温度的不同和时期的差异可能是导致以上结果存在出入的主要原因。这也说明,棉花的耐冷性是受多种因素(温度、遗传背景、生育时期等)共同影响的,单一因素的改变可能导致耐冷鉴定结果出现显著差异。
4 结论
低温冷害能够显著降低棉花种子萌发期吸胀阶段的相对吸水量和吸水速率,且材料之间存在差异。活力指数、萌发后芽干重、萌发后芽鲜重、平均发芽时间及平均发芽速度五项指标可作为陆地棉低温萌发鉴定的主要指标。品种新陆中 4号耐冷性最强,而中103026耐冷性最差,它们可以作为进一步研究棉花萌发期耐冷性机制的材料。POD酶活力、SOD酶活力、CAT酶活力和可溶性蛋白浓度可作为棉花萌发期抗冷鉴定的生理生化指标。
猜你喜欢
少儿科学周刊·儿童版(2022年10期)2022-06-27
军事文摘(2021年16期)2021-11-05
中学生数理化(高中版.高一使用)(2021年3期)2021-06-09
今日农业(2020年23期)2020-12-15
小读者(2020年4期)2020-06-16
劳动保护(2018年8期)2018-09-12
中学化学(2016年10期)2017-01-07
山东工业技术(2016年15期)2016-12-01
红领巾·萌芽(2015年6期)2015-08-14
小天使·一年级语数英综合(2014年7期)2014-06-26
