
肝细胞生长因子基因修饰的间充质干细胞治疗肝损伤的研究进展
2021-01-13 03:45:46修文丽毛成刚杨晨宇董蒨
中华细胞与干细胞杂志(电子版) 2020年6期
修文丽 毛成刚 杨晨宇 董蒨
肝损伤修复是指肝脏受到各种病理性因素(如感染、药物、毒物、手术和外伤等)刺激后,肝脏细胞受到破坏,继而机体启动损伤修复机制的过程。在此过程中,肝内炎症的过度反应、氧化应激、细胞过度凋亡和组织纤维化等,均可导致肝组织坏死、结构异常。严重者肝脏纤维结缔组织继续增生,假小叶形成,肝脏纤维化,进而发展为肝硬化,最终不可逆地走向肝衰竭[1]。尽管目前内科领域有各种保肝药物的研发应用,外科领域有计算机辅助手术系统更加精准指导手术[2-3],但减轻肝脏损伤、促进肝脏修复和再生、阻止肝纤维化始终是肝损伤治疗研究中的难点和热点。间充质干细胞(mesench ymal stem cells,MSCs)是一类能进行自我复制更新、具有分化为多种组织细胞潜能的干细胞,来源广泛,是目前进行基因改造优选的明星干细胞。MSCs 可旁分泌多种细胞因子,肝细胞生长因子(hepatocyte growth factor,HGF)是其中重要的一种,可发挥促进肝细胞DNA 合成、有丝分裂、抑制凋亡,调节炎症反应,促进血管再生,减轻组织纤维化等生物学效应。随着近年来对MSCs 研究的愈加深入,发现HGF 基因修饰的MSCs(HGF-MSCs)对肝损伤的治疗能够发挥更大的效果,本文就此展开综述。
一、MSCs
MSCs 是一种来源于胚胎发育早期的中胚层干细胞,拥有自我复制更新和多向分化潜能[4]。它广泛分布于骨髓、脐带和脂肪等多种组织和器官中。国际细胞治疗组织(International Society for Cellular Therapy)对鉴定人的MSCs提出了以下3 点标准[5]:(1)在标准且适宜的培养条件下,必须贴壁生长;(2)细胞表面标志物表达CD105+/CD73+/CD90+,同时造血干细胞表面标志物 CD34、CD45、CD11a、CD19 和人类白细胞抗原-DR 等阴性表达;(3)在体外经特殊条件诱导,能分化为机体多种细胞(如成骨细胞、成软骨细胞和成脂肪细胞等)。MSCs 除具有以上所述的生物学特性外,还具有来源途径广泛、类型多样、表面抗原性弱、免疫原性低和致瘤性极低等优势,从而使MSCs 具有广阔的发展前景。
在肝脏研究方面,有学者发现不同组织来源的MSCs 不仅形态和表型相似,而且都具备向功能性肝细胞分化的潜能[6]。MSCs 移植入肝脏受损的动物或人体内,可定植于受损肝组织,分化为正常肝脏细胞并在一定程 度上取代受损细胞[7-8],并且旁分泌大量细胞因子[9],通过多种信号通路诱导肝细胞增殖,调节炎症反应,促进血管再生,减轻纤维化[10-13],从而为肝脏疾病开辟了新的治疗思路和途径。
随着研究的深入,学者们发现移植后损伤部位的MSCs存活率低、分化效率低,单纯依靠MSCs对肝损伤的治疗作用有限。但是增加MSCs 的移植剂量可能会带来栓塞和组织坏死等未知风险,因此基因修饰MSCs 为科学家们提供了新思路[6]。
二、HGF
1984年Nakamura 等[14]首次从部分肝切除术后的大鼠血清中分离到HGF,研究发现HGF 是一种具有促进增殖效应的细胞因子。科学家们进一步研究发现,(1)HGF 系由α链(69 kDa)和β 链(34 kDa)通过一对二硫键链接构成的异二聚体。α 链中有类似于纤溶酶原和凝血酶中的kringle 结构,β 链中有类似丝氨酸蛋白酶的催化结构域。HGF 的β 链蛋白序列还与扩散因子同源;(2)HGF 主要由肝脏间质细胞分泌,如肝星状细胞、Kupff er 细 胞和血管内皮细胞等;(3)HGF 可与特异性跨膜受体c-Met 结合,激活细胞内信号转导分子,继而发挥HGF 的生物学效应。
HGF 是目前已知的促肝细胞有丝分裂最强的细胞因子,Mars 等[15]向无血清培养的原代肝细胞加入1 ng/mL 的外源性HGF,即可观察到其显著促进DNA 合成。除此之外,HGF还具有强有力的促进内皮细胞增殖和毛细血管管腔形成能力;抑制细胞凋亡的能力,减轻组织损伤;促迁移效应,能够提高多种细胞的运动能力;高浓度条件下的肿瘤抑制效应,抑制某些癌和肉瘤细胞系的生长[16-19]。
尽管HGF 具有如上众多的正面效应,但是HGF 的临床应用仍然有很多限制:(1)HGF 生理分泌量少,难以分离、富集和纯化;(2)HGF 是大分子的糖基化蛋白,在血液中半衰期短,需要反复注射才能达到有效血药浓度;(3)运输和保存困难;(4)致癌、促癌转移和诱导耐药等负面效应[20-21];(5)肝外器官的血液大量分流,直接影响了外源性HGF 对肝损伤的治疗效果[17]。
三、HGF-MSCs 治疗肝损伤
HGF 是MSCs 分泌的发挥生物学效应的主要细胞因子之一[22]。王荣等[23]研究发现,体外多次传代培养的人脐带间充质干细胞(human umbilical cord mesenchymal stem cells,hucMSCs)均可分泌细胞因子HGF,但分泌的HGF 量很少且随着MSCs 传代次数的增加而降低。加上MSCs 在损伤部位存活能力低下,严重影响MSCs 治疗效率。近年来,随着基因修饰技术的发展,针对HGF 基因修饰MSCs 获得的HGF- MSCs 的研究越来越多,学者们也逐渐发现HGF-MSCs对于肝损伤修复可能有更好的治疗效果(图1)[24-52]。
(一)抑制凋亡
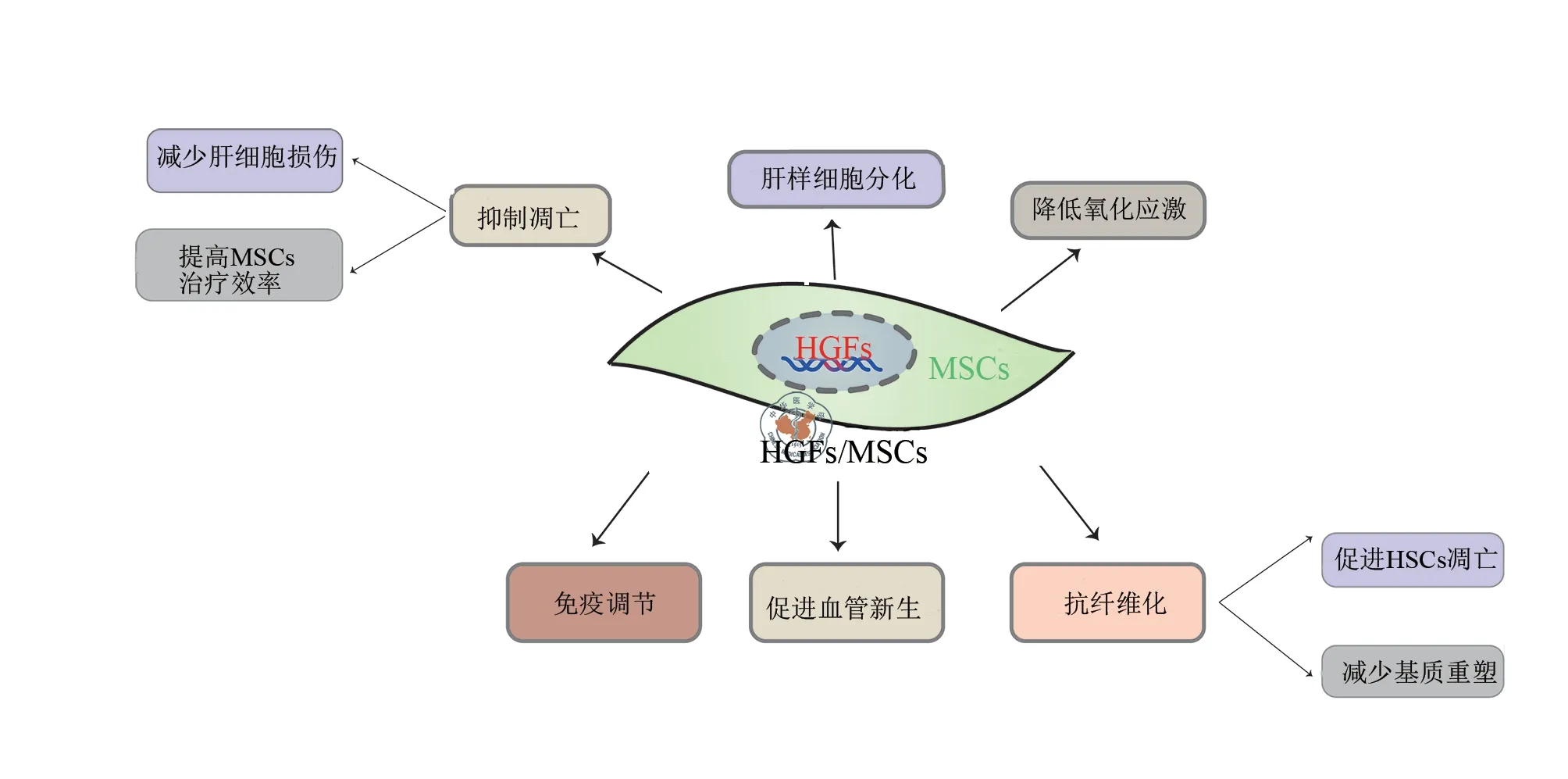
图1 HGF-MSC 治疗肝损伤
Zhang 等[24]在研究过表达HGF 的脂肪间充质干细胞(adipose mesenchymal stem cells,ADSCs)对放射线诱发的肝损伤大鼠的治疗作用中发现,HGF-ADSCs 治疗组染色质浓缩明显逆转,肝细胞凋亡明显减少,肝细胞再生相较于ADSCs 治疗组更显著。Jang 等[25]在研究HGF-MSCs 减轻对乙酰氨基酚诱导的肝细胞损伤中发现,HGF-MSCs 上清液处理的肝细胞活力增加,抗凋亡蛋白(Mcl-1)表达增加,促凋亡蛋白(Bad,Bik 和Bid)表达降低。Yu 等[26]将HGF-MSCs经门静脉注射至小体积肝移植术后(移植百分比为30%)的大鼠体内,结果发现大鼠死亡率下降,肝再生增强;同时HGF及其受体c-Met 的表达量增加,且与肝再生呈正相关关系。同样另有研究发现,当HGF/c-Met 信号途径中断时,再生肝脏的体积较小并且表现出广泛的肝细胞死亡[27]。另外,过量表达的HGF 还可以抑制MSCs 的凋亡。Choi 等[28]在研究过表达HGF 对过氧化氢(H2O2)诱导的MSCs 保护作用时发现,HGF-MSCs 内凋亡相关蛋白减少,凋亡细胞的数量下降,细胞活力提高了1.68倍。以上研究提示HGF-MSCs 通过增强HGF/c-Met 途径,发挥更显著的抑制肝细胞凋亡和保护MSCs 作用,提高MSCs 治疗效率,最终促进肝再生。
(二)诱导MSCs 向肝样细胞分化
HGF 是诱导MSCs 向功能性肝细胞分化重要且有效因子之一[22]。无论是HGF-MSCs 还是在MSCs 中加入外源性的HGF[29],MSCs 均可分化为功能性肝细胞。HGF 与其他细胞因子(胰岛素样生长因子1、白细胞介素1β 等)联合诱导可提高MSCs 分化的效率和质量[30-31]。由于HGF 是诱导分化的关键因子,因此HGF-MSCs 可能更有利于诱导MSCs 分化为肝样细胞,从而提高MSCs 的替代治疗效果。
(三)降低氧化应激
氧化应激是指在某些病理状态下,机体内氧化与抗氧化的动态平衡失衡,导致细胞死亡和组织损伤的过程[32]。Moon 等[33]将HGF-MSCs 移植到二甲基亚硝胺诱导的肝纤维化大鼠中发现,H2O2诱导肝细胞的毒性降低,肝细胞内活性氧种类减少,促凋亡蛋白和肝细胞凋亡减少。Choi 等[28]在H2O2诱导的HGF-MSCs 中发现,HGF 的过量表达使细胞内活性氧水平减少了27%,细胞活力明显增加,提示HGFMSCs 可增强MSCs 抗氧化能力。因此HGF- MSCs 在治疗肝损伤的过程中,可通过降低氧化应激抑制凋亡,而损伤局部MSCs 的存活数量的增加,将发挥更大的移植效果。
(四)免疫调节
MSCs 通过分泌HGF 减轻免疫反应,或使受体处于免疫耐受状态来调控炎症反应,从而减轻肝脏损伤[34]。MSCs 来源HGF 可抑制单核细胞发育成熟为树突状细胞[35],可有效抑制CD4+及CD8+T细胞增生[36]。Bian 等[37]研究显示,过表达HGF 的骨髓间充质干细胞(bone marrow mesenchymal stem cells,BMSCs)在体外混合淋巴反应中仍维持BMSCs原本的增殖和分化能力,但明显抑制了异常淋巴细胞增殖,并且对体外的细胞免疫具有更强的抑制效果。他们后续的实验发现HGF-BMSCs 和未经转染的BMSCs 免疫抑制功能相似,但HGF-BMSCs 针对部分免疫细胞(如NIH3T3 和CD11b+/-细胞等)具有更强的免疫调节作用。朱金海等[38]研究发现,HGF-BMSCs 注射与单独BMSCs 注射相比,更加显著减轻急性移植物抗宿主病中的急性排斥反应,并且大幅度延长受者的生存时间。由此可见,HGF 基因修饰明显增强了MSCs 的免疫抑制功能,更有效地减轻免疫排斥反应。
(五)抑制内皮细胞凋亡、促进血管新生
各种病理性刺激因素、炎症、氧化应激等在造成肝细胞损伤的同时,往往也造成血管结构破坏。肝内缺血不仅加剧肝损伤,而且不利于肝脏损伤修复,因此减轻血管损伤或者促进血管重建在肝损伤修复中具有重要意义。刘滋康 等[39]通过结扎大鼠股动脉来模 拟局部肢体缺血,并且以它为模型研究HGF-BMSCs 在体内促血管新生效应时发现,HGF-BMSCs组血流灌注水平明显高于MSCs组,而且HGF- BMSCs 与MSCs 相比,内皮细胞促增殖、促迁移及重建血管结构等效应更显著。Su 等[40]也有相似的发现,即HGFMSCs组毛细血管密度最高,提示这可能与HGF-MSCs 诱导更强的血管新生效应密切相关。陈安平等[41]研究HGFBMSCs 改善慢性缺血心肌时发现,缺血区血管生成显著增加,从而改善了心肌缺氧微环境。因此在肝脏损伤时,MSCs旁分泌的HGF 可能也存在以上机制抑制血管内皮细胞凋亡,促进肝脏新生血管的形成,从而改善肝脏缺血,而HGFBMSCs 可能增强了这种效应。
(六)抗纤维化
肝纤维化是慢性肝病发展为肝硬化的中间环节,逆转肝纤维化是防止肝衰竭的关键阶段。目前普遍认为,肝纤维化进展的首要因素和中心环节是肝星状细胞(hepatic stellate cells,HSCs)的激活。在肝细胞损伤、炎症等始动因素作用下,HSCs 被激活,转变为肌成纤维细胞并产生大量细 胞外基质[42],基质重塑,最终导致肝纤维化。因此促进HSCs 的凋亡、减少基质重塑可能是逆转肝纤维化的重要途径[43]。以往的研究表明,在肝硬化大鼠模型中应用外源性HGF 或单独移植MSCs 均可减轻肝脏纤维化[44],其原因可能与抑制HSCs 的激活和减少胶原在肝脏沉积有关[45],HGF-MSCs 增强了这些效应[46]。
1.诱导及促进HSCs 的凋亡:目前认为HSCs 的凋亡水平同肝纤维化的逆转程度基本一致[42]。Kim 等[47]研究报道慢性肝损伤时发现,HGF 可能通过JNK 磷酸化促进活化的HSCs 凋亡。苏思标等[48]发现MSCs 抑制HSCs 增殖的机制可能与RhoA-P27通路使HSCs细胞周期停滞有关。随后学者们发现MSCs 发挥HSCs 的促凋亡作用可能与HGF有关[49],作用于神经生长因子P75 信号通路、Fas/FasL通路等逆转肝纤维化[50]。HGF-MSCs 也可能从以上途径促进HSCs 的凋亡,甚至发挥更好的效果。
2.减少基质重塑:当肝组织受损时,HSCs 被激活,转化为肌成纤维细胞,同时基质和胶原合成增加,降解减少,合成和降解平衡被打破,导致基质重塑和肝脏纤维化[43]。Yu等[51]在研究减轻小体积肝移植大鼠模型中缺血再灌注和慢性损伤时发现,HGF-MSCs 明显抑制小体积肝移植大鼠肝纤维化的形成,从而促进肝再生,降低大鼠死亡率;在这一过程中,HGF-MSCs 的抗纤维化作用不仅与调节HSCs 活化和凋亡有关,还与显著减轻胶原的沉积有关。Pulavendran 等[52]也发现HGF-BMSCs 可使Ⅰ型胶原明显减少,细胞外基质纤维溶解,进而逆转肝纤维化。Zhang 等[24]在研究改善辐射诱导的肝纤维化过程中发现,HGF-ADSCs 可能通过下调α-平滑肌肌动蛋白和纤维连接蛋白等,减少基质重塑。
综上所述,HGF-MSCs 不仅不会影响MSCs 原有的干细胞生物学特性,而且与单纯的MSCs 或HGF 移植相比对肝损伤有明显优势。HGF-MSCs 中过量表达的HGF 对MSCs抗氧化和抗凋亡的保护作用,解决了移植区MSCs 高凋亡率问题。HGF-MSCs 作为HGF 基因的细胞载体在损伤部位增加HGF 的分泌[53],解除了HGF 的临床应用部分限制。因此HGF-MSCs 移植在肝损伤修复中充分发挥了MSCs 和HGF优势,弥补它们在临床应用上的部分劣势,发挥 “1+1>2”的治疗效果[54]。
四、问题和展望
上述众多的体内外实验和临床试验均展示MSCs 在促进肝细胞再生和修复方面具有广阔的前景,同时也证实了MSCs 的安全性,未发现致瘤性。但HGF-MSCs 能够高表达HGF,HGF 的配体c-Met 在肿瘤中也是高表达,因此HGF- MSCs 是否在致瘤方面有更大的风险,HGF-MSCs 能否精准高效地调控HGF 分泌等还有待进一步研究。相信随着干细胞研究技术的进一步成熟,一定能够给肝病患者带来新的曙光。
猜你喜欢
今日农业(2022年13期)2022-09-15 01:21:20
传染病信息(2022年3期)2022-07-15 08:24:28
昆明医科大学学报(2022年2期)2022-03-29 00:52:18
肝博士(2021年1期)2021-03-29 02:32:16
生物学通报(2020年10期)2020-08-13 08:52:26
知识经济·中国直销(2017年10期)2017-11-07 02:39:52
癌变·畸变·突变(2016年3期)2016-02-27 06:15:36
哈尔滨医药(2015年4期)2015-12-01 03:57:54
医学研究杂志(2015年6期)2015-07-01 17:40:08
中国卫生(2014年2期)2014-11-12 13:00:14
