
水肥条件对煤矸石土壤上两种牧草光合及生物量的影响
2021-01-12 03:37:28王进鑫张玉玉胡杭丽
水土保持研究 2021年1期
马 戌, 王进鑫, 张玉玉, 胡杭丽
(1.西北农林科技大学 水土保持研究所, 陕西 杨凌 712100; 2.西北农林科技大学 资源环境学院, 陕西 杨凌 712100 )
煤矸石是煤炭开采和洗选过程中产生的一种排放量最大、占地最多、污染最严重的固体废弃物,其资源化利用率相对较低[1-2]。在我国,煤矸石山植被恢复总是在覆土后采用工程措施和林草措施相结合而进行的。尽管煤矸石山植被恢复已初见成效,但是煤矸石覆土土层很薄,土壤物理结构不良,保水持水能力很差,养分极端贫瘠,氮磷钾及有机质含量极低,植物生长发育所需的水分和养分严重匮乏[3-4],致使林草成活率低、保存率低,植被恢复难度高,绿化工程投资大等问题还远远没有解决。目前,已有的研究多集中于水分、温度、盐害、养分等单一限制性因子[5-8],而关于各因素交互效应对煤矸石山植被恢复影响的研究鲜有报道。对西北干旱荒漠区煤矸石山植被恢复而言,水分和养分是维持植物生长的关键因子,合理的水肥条件是植物长势良好的重要保证。研究表明水肥耦合有利于提高林草生产力和水肥利用效率,促进光合作用,增加生物量积累[9-10]。谷裕等[11]认为矿区植被恢复与土壤水分和养分之间存在复杂的交互效应,两者相互依存且制约,土壤养分特性相对于土壤水分对矿区植物生长的影响更大。但是也有研究认为土壤水分比土壤养分特性对矿区植物生长的影响更大[12-13]。因此,研究西北干旱荒漠区植物光合及生物量的水肥耦合规律,明确其最优水肥方案是煤矸石山植被恢复亟待解决的核心问题。
Huang等[14]认为西北干旱荒漠区煤矸石山植被恢复,应该以草本植物为主。基于西北地区土壤结构差、水分短缺、气候恶劣等立地条件与适生植物调查[15-18],草木樨和蒙古冰草适应性和抗逆性强,耐寒、耐旱、耐贫瘠、耐盐碱,根系发达,生长迅速,生物量大,防风固沙能力强,是西北干旱荒漠区采煤迹地植被恢复的先锋牧草。因此,本文以草木樨和蒙古冰草为研究对象,探讨不同水肥条件对煤矸石土壤上2种典型草本植物光合及生物量的影响规律,从而确定西北煤矸石山植被恢复的最优水肥方案,必将为西北干旱荒漠区采煤迹地植被重建与保育提供重要的理论依据和技术支撑。
1 材料与方法
1.1 试验材料
试验选取白花草木樨(MelilotusalbusDesr.)和蒙古冰草(AgropyronmongolicumKeng)作为供试材料,其种子均购于宁夏远声绿阳草业生态工程有限公司,采摘于宁夏当地。试验所需氮肥和磷肥分别为尿素(含氮46%)和重过磷酸钙(含P2O546%),其中尿素溶于水后随水施入土壤,重过磷酸钙以穴施方式施入土壤。
1.2 供试土壤
试验供试土样由表层沙土和母质3∶1混合而成,于2019年3月取自宁夏灵武羊场湾排矸场,过0.5 cm筛作为栽培基质。土壤理化性质为:土壤田间最大持水量13.50 %,容重1.44 g/cm3,总孔隙度31.36 %,pH 8.54,土壤水溶性盐总量0.77 g/kg,有机质2.40 g/kg,碱解氮9.14 mg/kg,有效磷4.92 mg/kg,有效钾58.89 mg/kg。土壤结构性较差,蓄水保墒能力差,氮、磷元素含量较低,钾元素含量较高,因此,试验仅对氮肥和磷肥进行研究。
1.3 试验设计与方法
盆栽试验采用3因素(土壤相对含水量、施氮量和施磷量)5水平二次回归通用旋转组合设计,分别以草木樨和蒙古冰草的叶片净光合速率、水分利用效率和单株总生物量(干重)为目标函数构建二次回归模型。γ=1.682,中心试验点m0=6,每种牧草设有20个处理,每个处理10个重复。根据供试土壤基础肥力状况,结合当地降雨量、水肥施用量以及草木樨和蒙古冰草生长发发育的水肥特性,从而确定了2种牧草的三因素水平上下限,其编码值见表1。设计土壤相对含水量、施氮量和施磷量3个因素依次为Z1,Z2,Z3,其中第j个因素的上、下水平分别为Z2j,Z1j,根据二次回归通用旋转组合设计确定各因素的零水平Z0j和变化间距Δj如下:
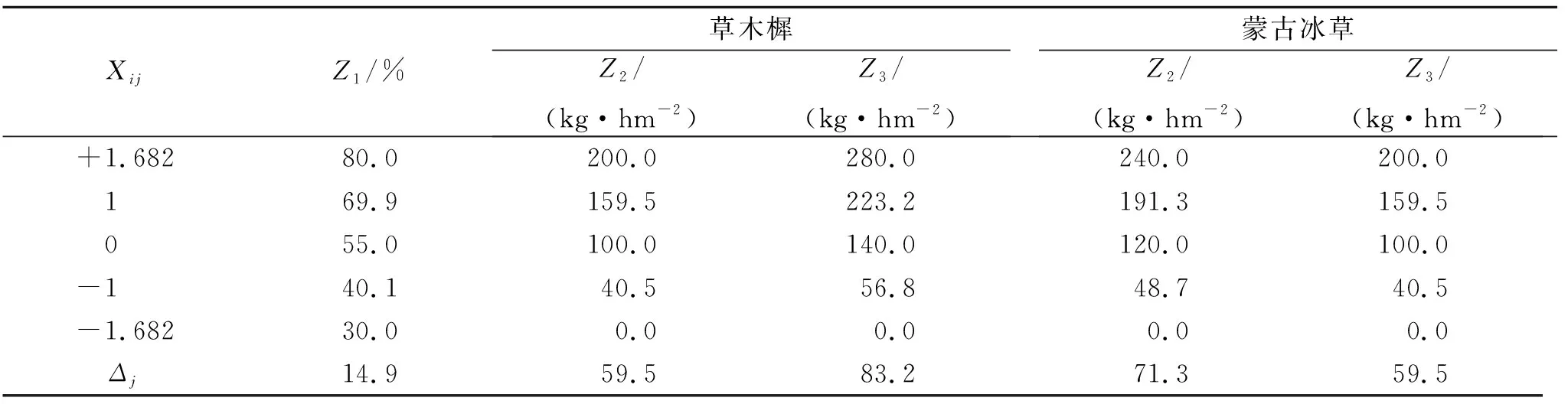
表1 试验因素水平及编码值
式中:Z1为土壤相对含水量;Z2为施氮量;Z3为施磷量;Z2j,Z1j分别为第j个因素的上、下水平;Z0j为各因素的零水平;Δj为各因素的变化间距;γ=1.682。
对各因素各水平进行编码,土壤相对含水量、施氮量和施磷量的编码值分别记为X1,X2和X3,其编码值计算公式如下:
式中:Xij为各因素各水平的编码值;X1为土壤相对含水量的编码值;X2为施氮量的编码值;X3为施磷量的编码值;Δj为各因素的变化间距。
试验地点位于西北农林科技大学南校区温室大棚,于2019年3月15日进行播种,充分灌水,待其幼苗长出3片真叶(即4月20日)时进行间苗,每盆保留10株长势相对一致的幼苗。试验从6月1日开始每天早上8:00通过称重法按照设计重量补水,使各处理盆栽土壤相对含水量维持在设定的水平范围(如80%±5%)至试验结束,同时根据试验方案,将尿素总量的30%,40%,30%分别于5月19日、6月10日、6月24日采用水溶法施入土壤,重过磷酸钙总量的一半分别于5月19日和6月10采用穴施法施入土壤。
采用便携式光合仪(CIRAS-3)分别于7月25月、8月1日对2种牧草进行两次光合测定,主要包括叶片的净光合速率(Pn)、蒸腾速率(Tr)和水分利用效率(WUE)等指标。选定一个晴朗的上午,从8:00—12:00选择草木樨和蒙古冰草各处理完全伸展的成熟叶片进行光合测定,每种牧草每个处理重复8次,求平均值即为其光合参数。
试验结束后(8月21日),挖出所有植株,清洗并用吸水纸吸干根系表面的水分,先在100~105℃杀青10 min,然后把烘箱的温度降到70~80℃左右,烘至恒重,将干样放入干燥器中冷却至室温,取出称重,重复10次,求得平均值即为单株生物量(B)。
1.4 数据处理
采用Excel 2010软件对试验数据进行统计与整理;采用Design-Expert 8.0.6软件建立2种牧草水肥耦合回归模型,用ANOVA程序做方差分析,t检验法对模型回归系数进行显著性检验,响应面分析交互效应,采用DPS数据处理系统对2种牧草的水肥耦合回归模型进行约束条件下的模型寻优。
2 结果与分析
2.1 水肥耦合回归模型的建立及检验
将不同水肥条件下草木樨和蒙古冰草的叶片净光合速率、水分利用效率和生物量列于表2中,采用Design-Expert软件对表2试验结果进行多元线性回归分析,分别建立其关于3个因素的二次回归模型(1),(2),(3),(4),(5)和(6):
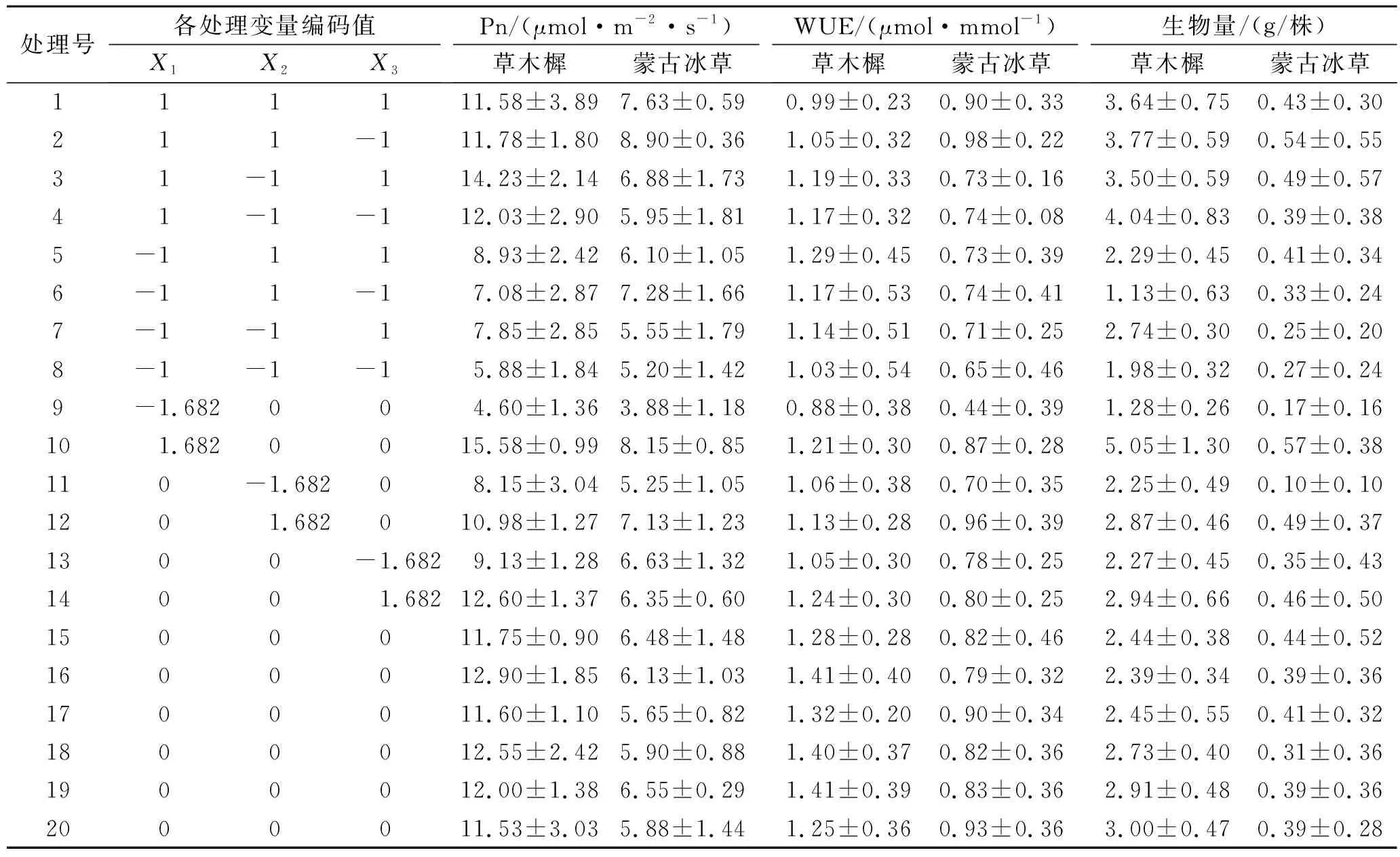
表2 试验方案设计与结果统计表
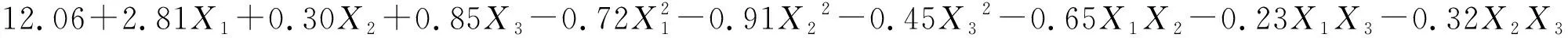
(1)
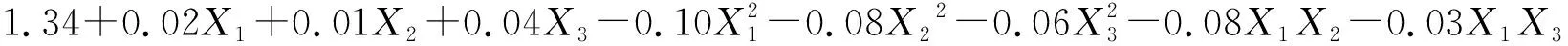
(2)
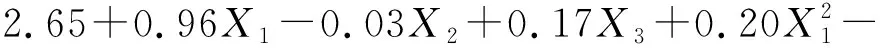
0.10X2X3
(3)
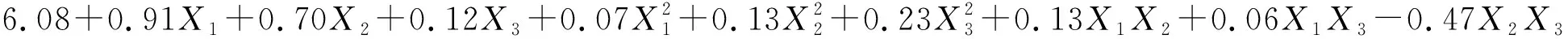
(4)
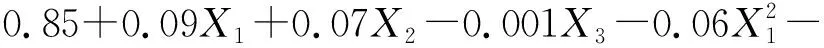
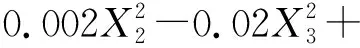
(5)
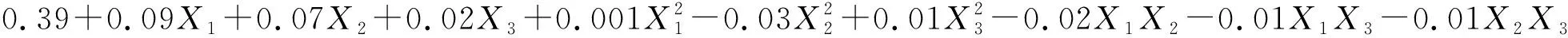
(6)
式中:Pn1和Pn2分别表示草木樨和蒙古冰草的叶片净光合速率[μmol/(m2·s)];WUE1和WUE2分别表示其叶片水分利用效率(μmol/mmol);B1和B2分别表示其单株生物量(g/株);X1为土壤相对含水量编码值;X2为施氮量编码值;X3为施磷量编码值。
水肥耦合回归模型经显著性检验,F1
2.2 主因子效应分析
通过对草木樨和蒙古冰草叶片净光合速率、水分利用效率和生物量水肥耦合回归模型的偏回归系数进行标准化,比较其绝对值大小可直接判断各因素对目标函数的影响程度。
从上述回归方程可知,草木樨和蒙古冰草水肥耦合回归模型的一次项系数为正值,说明在试验设计范围内,土壤水分、氮肥和磷肥各因子均对2种牧草叶片净光合速率、水分利用效率和生物量具有明显地促进作用。三因素对草木樨光合及生物量的影响程度从大到小为土壤相对含水量>施磷量>施氮量,蒙古冰草为土壤相对含水量>施氮量>施磷量。由此可知,在试验区,草木樨和蒙古冰草光合及生物量对土壤水分的响应最为敏感,磷肥对草木樨生长发育的贡献大于氮肥,氮肥对于蒙古冰草的贡献大于磷肥,这可能是因为豆科牧草本身具有一定固氮能力,施用少量氮肥就可以满足草木樨等豆科牧草的生长发育[19],禾本科牧草因其本身没有固氮能力,要想改变其生长特性和提高其生物量就必须增施适量氮肥,磷肥可以促进禾本科植物的固氮作用[20]。
2.3 单因子效应分析
2.3.1 净光合速率回归模型单因子效应分析 采用降维法得到土壤水分、施氮量和施磷量对2种牧草叶片净光合速率的偏回归子模型,可知草木樨的二次项系数均为负值,蒙古冰草均为正值,据其描绘出相应变化曲线(图1)。其中草木樨叶片净光合速率随着水肥用量的增加呈现开口向下的抛物线变化(图1A),说明其随着水肥用量的增加先显著升高后降低,最大值在水肥设计范围之间,符合报酬递减率。而蒙古冰草叶片净光合速率随着水氮用量的增加呈现开口向上的抛物线变化(图1B),其在水肥用量最大时达到最大值,水肥设计上限过小。由图1可以明显看出,在相对处理条件下,草木樨叶片净光合速率远高于蒙古冰草。
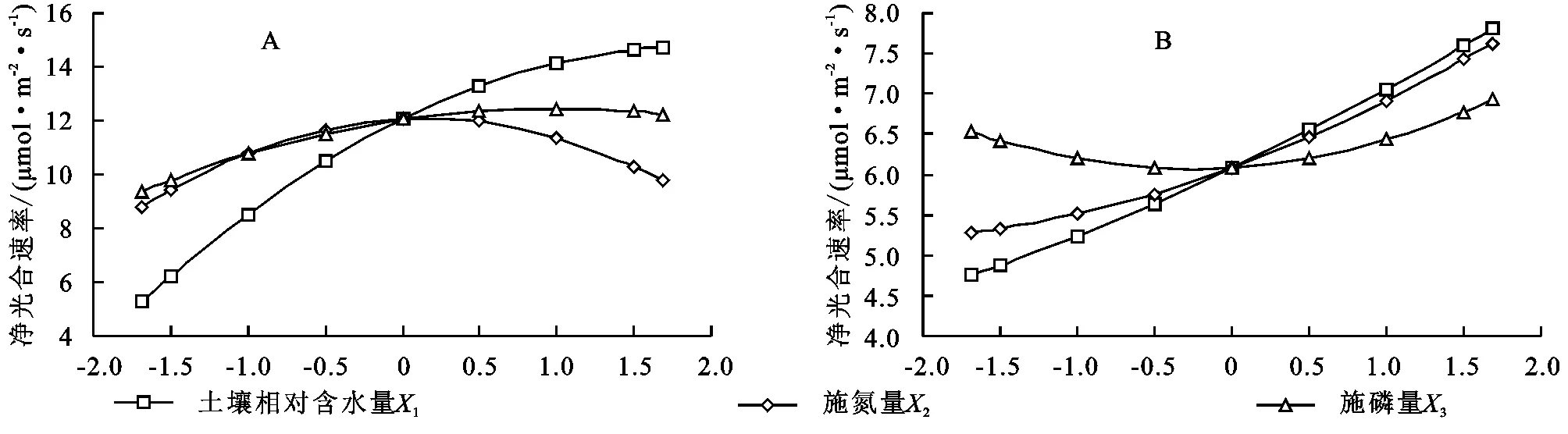
图1 土壤水分、氮肥和磷肥单因素对2种牧草净光合速率的单因子效应
2.3.2 水分利用效率回归模型单因子效应分析 根据土壤水分、施氮量和施磷量对2种牧草叶片水分利用效率的偏回归子模型,草木樨和蒙古冰草的二次项系数均为负值,描绘出其叶片水分利用效率随着水肥用量的增加均呈现开口向下的抛物线变化曲线(图2)。说明草木樨和蒙古冰草叶片水分利用效率均随着水肥用量的增加先升高后显著降低,即在适量的水肥用量时其达到最大值,此后随着水肥用量的继续增加逐渐降低。这可能是因为随着肥料的增加草木樨净光合速先增大后减小,蒸腾速率变化不大,而且蒙古冰草蒸腾速率的增加幅度大于净光合速率,致使其水分利用效率也呈现先升高后降低的趋势。
从图2可以看出;在试验设计范围内,土壤水分由-1.682增加至0.10时,草木樨水分利用效率由1.03 μmol/mmol升高至最大值为1.34 μmol/mmol,此后逐渐降低至1.11 μmol/mmol。氮肥为0.06时,水分利用效率升高至最大值为1.34 μmol/mmol,磷肥为0.33时,水分利用效率升高至最大值为1.35 μmol/mmol,此后逐渐降低(图2A)。土壤水分由-1.682增加至0.75时,蒙古冰草水分利用效率由0.53 μmol/mmol升高至最大值为0.88 μmol/mmol,此后逐渐降低至0.83 μmol/mmol。氮肥从-1.682增加到1.682时,蒙古冰草水分利用效率从0.73 μmol/mmol不断升高至最大值为0.96 μmol/mmol,磷肥为-0.03时,水分利用效率升高至最大值为0.85 μmol/mmol,此后逐渐降低至0.79 μmol/mmol(图2B)。在相同处理条件下,草木樨叶片水分利用效率明显大于蒙古冰草。
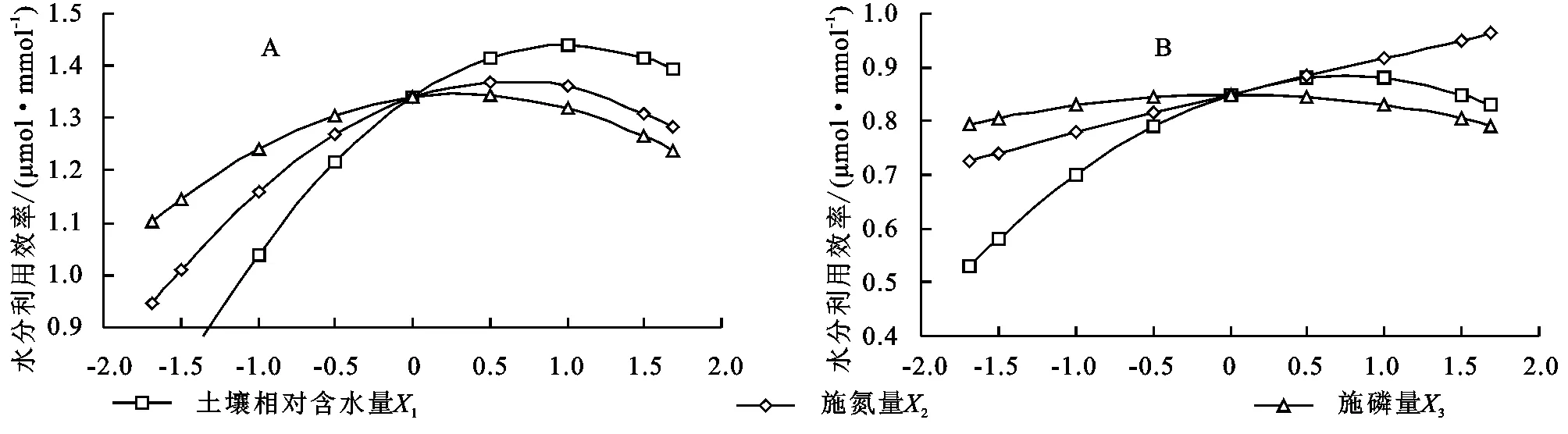
图2 土壤水分、氮肥和磷肥单因素对2种牧草水分利用效率的单因子效应
2.3.3 生物量回归模型单因子效应分析 根据土壤水分、施氮量和施磷量对2种牧草单株生物量的偏回归子模型,草木樨和蒙古冰草的水分与磷肥的二次项系数均为正值,氮肥二次项系数为负值,描绘出相应的变化曲线(图3)。
结果表明草木樨和蒙古冰草生物量均随着土壤水分和施磷量的增加显著升高,随着施氮量的不断增加先升高后降低。从图3可以明显看出,土壤水分对2种牧草的生物量具有显著的促进作用,肥料对草木樨增产不明显(图3A),氮肥对蒙古冰草的增产作用大于磷肥(图3B)。在相同处理条件下,草木樨生物量明显远大于蒙古冰草。
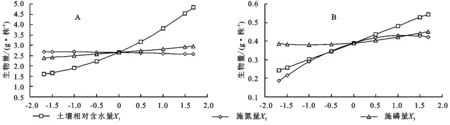
图3 土壤水分、氮肥和磷肥单因素对2种牧草生物量的单因子效应
2.4 因子间交互效应分析
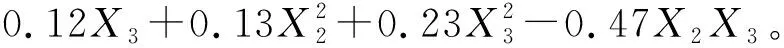
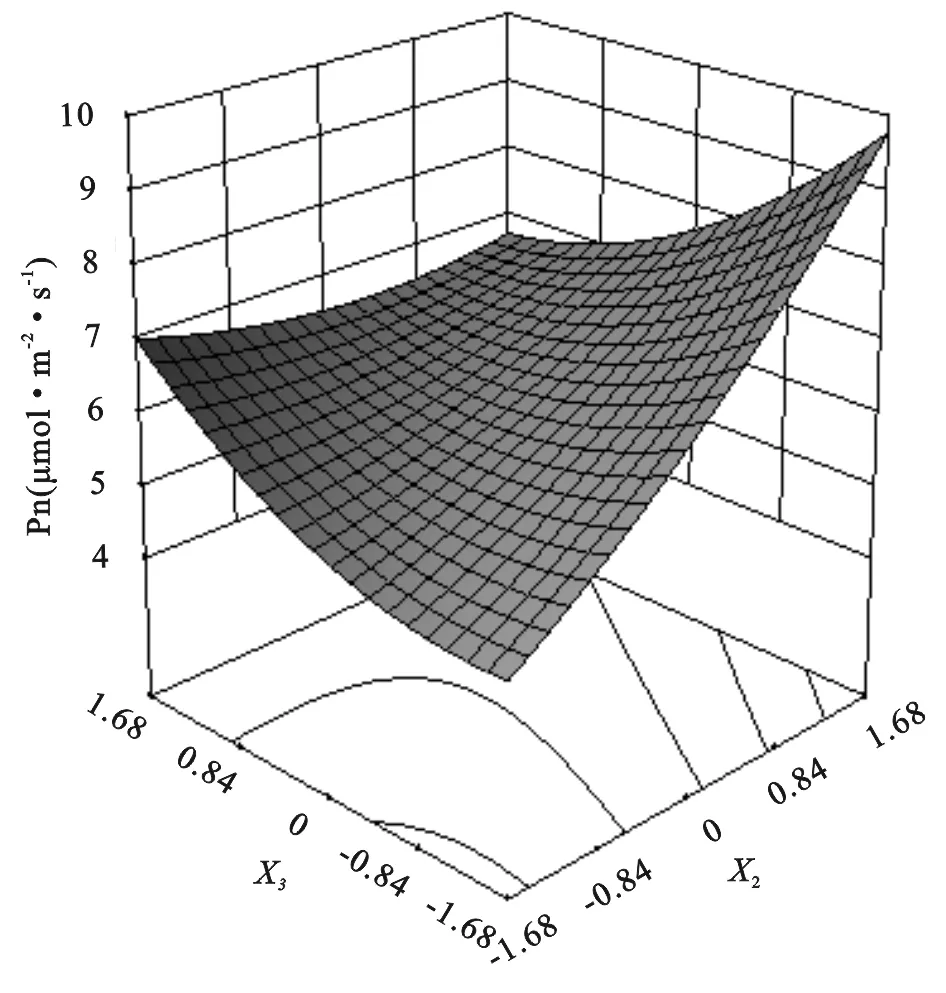
图4 蒙古冰草净光合速率氮磷交互效应
由上述方程和图4可以看出:当X2和X3均为-1.682时蒙古冰草叶片净光合速率最小,即不施氮肥和磷肥时,净光合速率最小为4.39 μmol/(m2·s)。蒙古冰草叶片净光合速率随着施氮量和施磷量的增加而缓慢增大。当X2=1.682且X3=-1.682时净光合速率最大,即施氮量为240.0 kg/hm2且不施磷量时,净光合速率达到最大值为9.40 μmol/(m2·s)。氮磷耦合作用对蒙古冰草的净光合速率表现为负效应,说明随着氮或磷的增加,蒙古冰草净光合速率会随着另一因素的增加而降低,其中,氮肥对蒙古冰草净光合速率的增幅远大于磷肥,磷肥会消减氮肥的肥效作用。
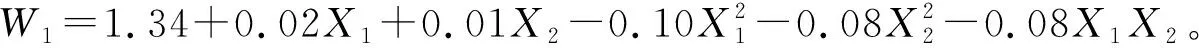
由上述方程和图5可以看出;当X1和X2均为-1.682时草木樨叶片水分利用效率最小,即土壤相对含水量为30.0%且不施氮肥时,水分利用效率最小为0.57 μmol/mmol。草木樨叶片水分利用效率随着水氮的增加先升高后降低。当草木樨水分利用效率最大时,X1=0.14和X2=-0.03是临界效应分界点,即土壤相对含水量为57.09%且施氮量为98.22 kg/hm2时,叶片水分利用效率达到最大值为1.34 μmol/mmol,此后逐渐降低。说明在临界效应分界点之前水氮与草木樨水分利用效率呈正相关,即适量的土壤水分和氮肥可以相互促进,不断提高草木樨水分利用效率,反之呈负相关。
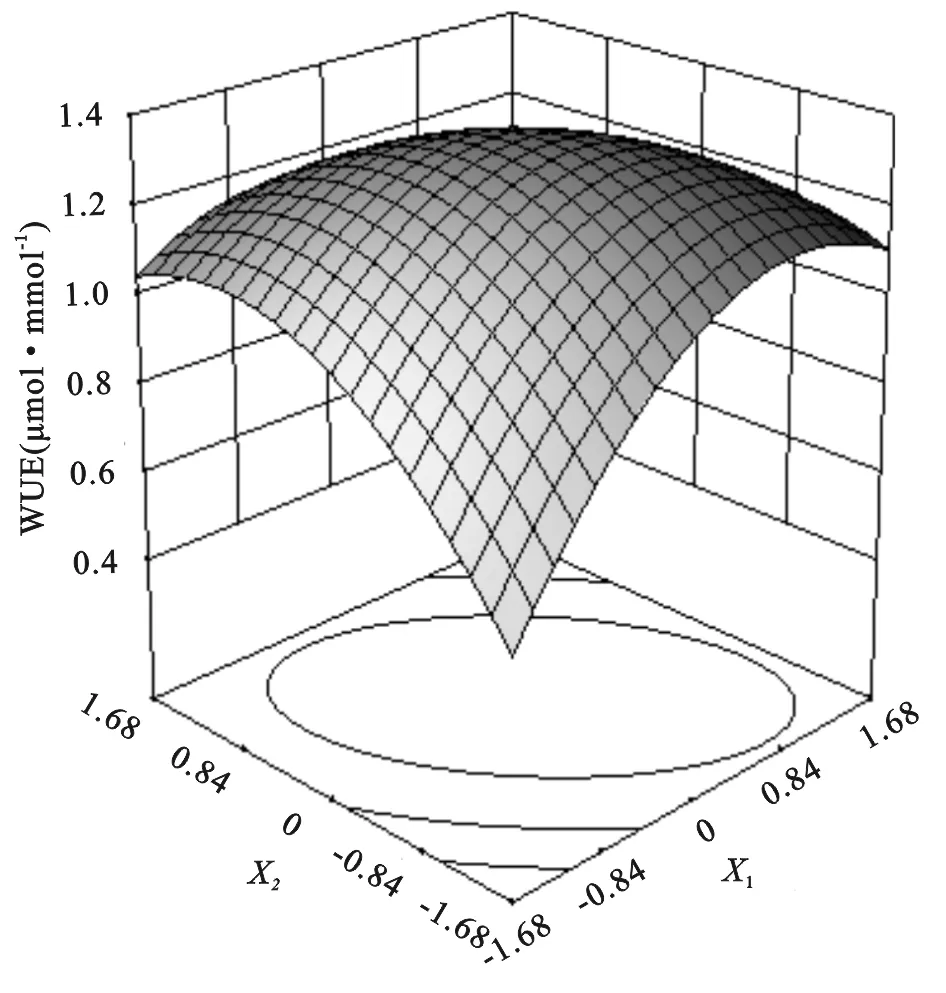
图5 草木樨水分利用效率水氮交互效应
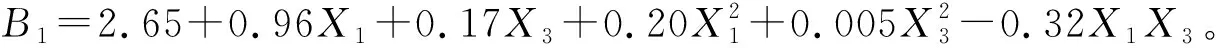
由上述方程和图6可以看出:当X1和X3均为-1.682时草木樨生物量最小,即土壤相对含水量为30.0%和不施磷肥时,生物量最小为0.42 g/株。当X1=1.682和X3=-1.682,即土壤相对含水量为80%和不施磷肥时,生物量达到最大值为5.46 g/株。说明草木樨生物量随着水磷施用量的增加不断升高,水磷交互作用对草木樨具有显著的增长作用,但是在高水分条件下,草木樨对磷肥的需求量较低。
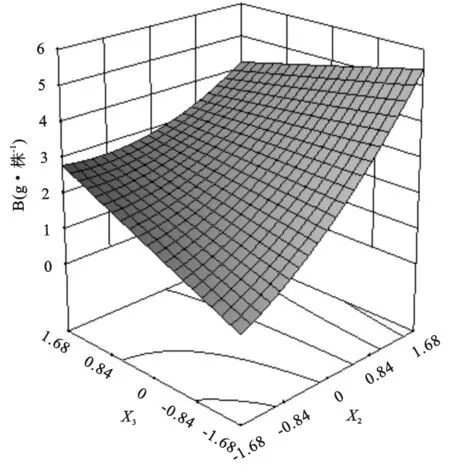
图6 草木樨生物量水磷交互效应
2.5 模型寻优
2.5.1 草木樨的最优水肥方案 在试验结果分析及模型拟合的基础上,采用Design-Expert软件对草木樨光合及生物量的回归模型在约束条件下进行模型寻优,其约束条件为水肥使用量最小的基础上使其光合及生物量达到最大,由此得到当X1=0.12,X2=-0.14,X3=-1.16时,草木樨净光合速率最大为12.61 μmol/(m2·s),水分利用效率最大为1.21 μmol/mmol,生物量最大为3.23 g/株。根据试验所得的水肥耦合回归模型,每个因素取5个水平:1.682,1,0,-1和-1.682,以所有处理的试验结果平均值作为系统优化栽培方案的指标临界值,采用DPS数据处理系统对53=125个方案进行寻优。在125个方案中,净光合速率大于10.64 μmol/(m2·s)
的组合有50个,生物量高于2.78 g/株的方案共有65个,其水、氮和磷最优组合方案见表3和表4。
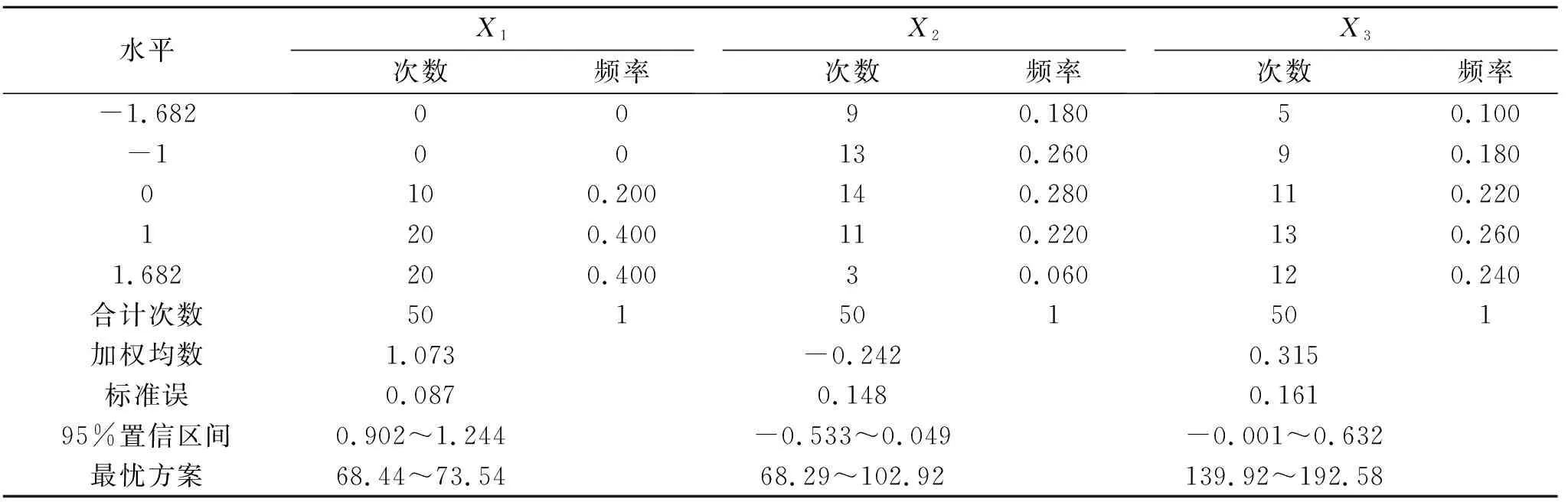
表3 草木樨净光合速率大于10.64 μmol/(m2·s)的寻优方案及频率
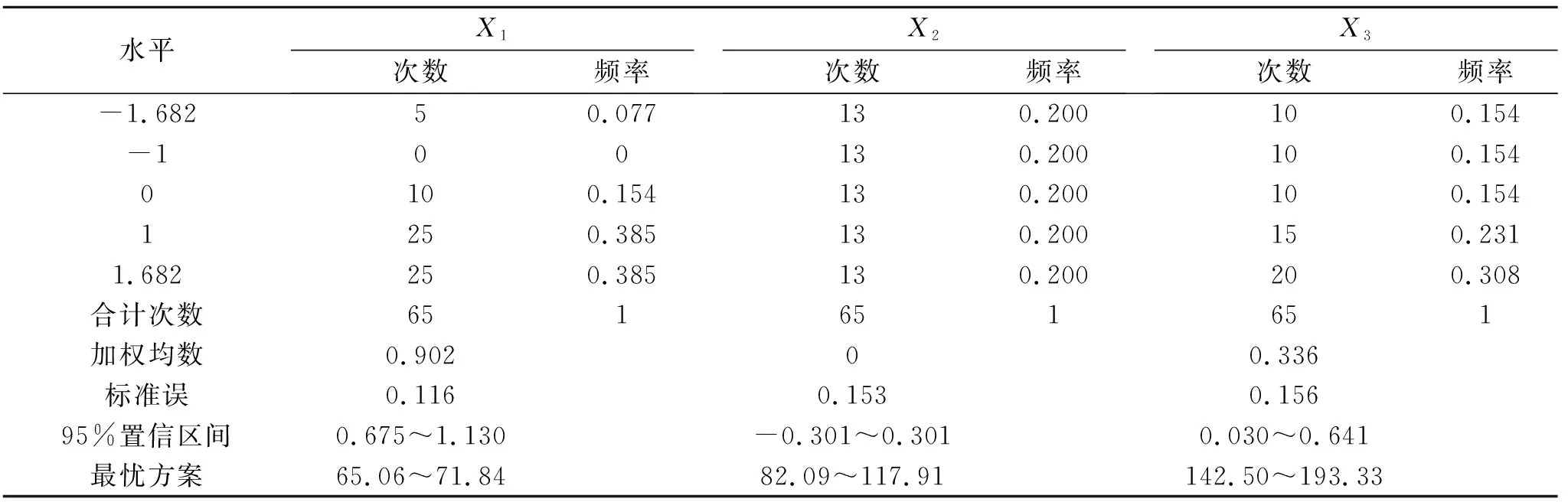
表4 草木樨生物量大于2.78 g/株的寻优方案及频率
综合表3和表4中净光合速率和生物量的最优方案,两者的水肥施用量范围基本一致,为增加试验结果的准确性与可靠性,可将其相交部分视为在水肥用量较低的基础上要使得草木樨生长发育良好的最优水肥方案:土壤相对含水量68.44%~71.84%,施氮量82.09~102.92 kg/hm2,施磷量139.92~192.58 kg/hm2。
2.5.2 蒙古冰草的的最优水肥方案 采用Design-Expert软件对草木樨光合及生物量的回归模型在约束条件下进行模型寻优,当X1=0.16,X2=-1.56,X3=-1.68时,蒙古冰草净光合速率最大为8.67 μmol/(m2·s),水分利用效率为最大0.98 μmol/mmol,生物量最大为0.34 g/株。每个因素取5个水平:1.682,1,0,-1和-1.682,组成125个方案,其中,净光合速率大于6.37 μmol/(m2·s)的组合有53个,生物量高于0.38 g/株的方案共有65个,其水、氮和磷最优组合方案见表5和表6。
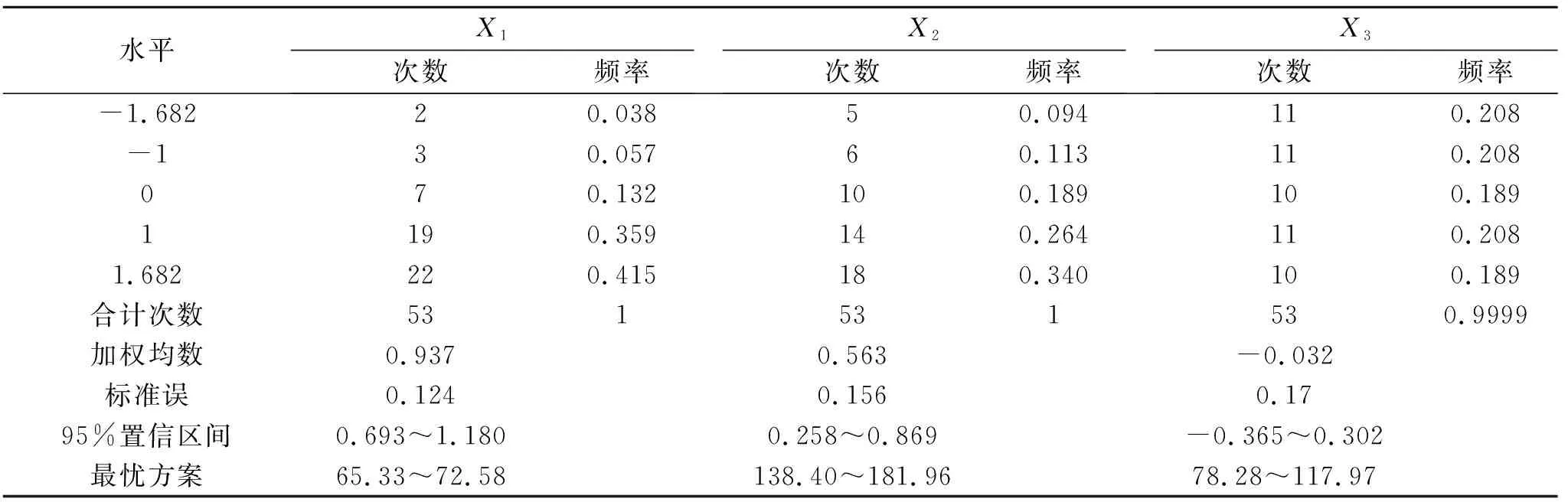
表5 蒙古冰草净光合速率大于6.37 μmol/(m2·s)的寻优方案及频率
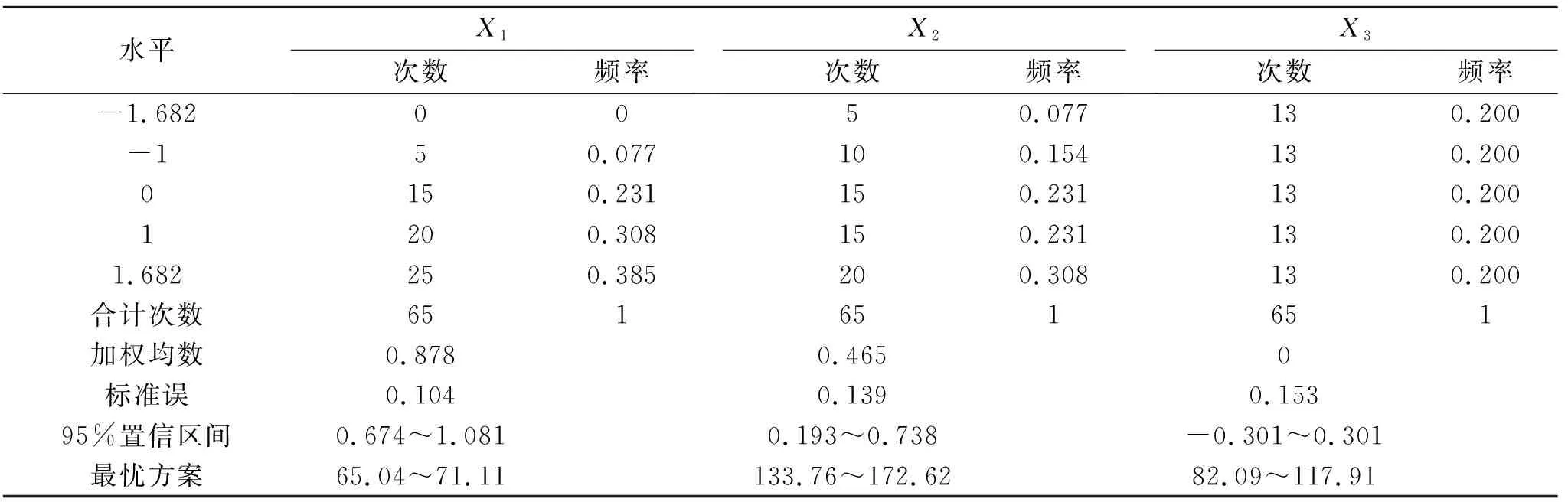
表6 蒙古冰草生物量大于0.38 g/株的寻优方案及频率
综合表5和表6中净光合速率和生物量的最优方案,两者的水肥施用量范围基本一致,为增加试验结果的准确性与可靠性,可将其相交部分视为在水肥用量较低的基础上要使得蒙古冰草生长发育良好的最优水肥方案:土壤相对含水量65.33%~71.11%,施氮量138.40~172.62 kg/hm2,施磷量82.09~117.91 kg/hm2。
3 讨 论
在西北干旱荒漠区,水分是植被恢复的限制性因子,土壤水分对林草的生长发育影响深远[21],当土壤相对含水量较低时,植物的生长生理指标都会受到不良的影响[22]。草木樨和蒙古冰草的气体交换参数多在土壤相对含水量为80%时较高,这与马宇飞[23]研究中草木樨光合作用最适宜点在相对田间持水量78.4%的结果相近。磷肥对草木樨生长发育的贡献大于氮肥,氮肥对于蒙古冰草的贡献大于磷肥,这与前人的研究结果基本一致[24-25]。在相同处理条件下,草木樨的光合及生物量都显著高于蒙古冰草,姜慧敏[26]通过比较豆科牧草与禾本科牧草的生理生态指标,也认为豆科牧草的光合参数均大于禾本科牧草。因此,西北干旱荒漠区采煤迹地植被恢复应该首选草木樨,其次是蒙古冰草。
水肥耦合效应对植物可产生3种不同的结果或现象,即协同效应、叠加效应和拮抗效应,从试验结果可以看出,不同水肥条件下草木樨的净光合速率等光合参数明显高于只灌水或只施氮量或只施磷肥。Bronson等[27]也发现水肥耦合能较好地促进植物生长,提高水分和养分的利用效率,其效果比只进行水分管理和养分管理更好。尹光华等[28]认为水肥耦合是通过扩大叶面积、提高叶片蒸腾速率、增加叶绿素含量、增大叶片气孔导度、降低胞内二氧化碳浓度来促进叶片光合作用的。杜灵通等[29]通过近17 a来宁东煤炭基地植被生态系统特征调查,表明植被生态系统蒸散增长率比生产力增长速率更快,这是导致区域水分利用效率下降的根本原因。在干旱胁迫条件下,适量施磷有利于促进植物根系发达,增强其抗旱性,在少磷或缺磷的条件下,草木樨蒸腾速率变化不大,净光合速率明显下降从而导致其生物量降低。蒙古冰草水分利用效率与土壤相对含水量或施氮量均呈正相关关系,施磷量对其水分利用效率的影响较小,因而在种植蒙古冰草的水肥管理过程中施氮量应多于施磷量,草木樨则相反,这与试验相符合。
4 结 论
(1) 试验建立了2种牧草光合及生物量的水肥耦合回归模型,均达到了显著水平且具有很强的代表性,适应于预测西北干旱荒漠区煤矸石山土壤上不同水肥条件下草木樨和蒙古冰草的光合速率、水分利用效率和生物量。
(2) 土壤水分对2种牧草的光合及生物量最敏感;磷肥对草木樨生长发育的贡献大于氮肥,氮肥对蒙古冰草的贡献大于磷肥;三因素交互作用对2种牧草水分利用效率的影响程度从大到小均是水氮>水磷>氮磷;对于生物量的三因素交互作用大小而言,草木樨为水磷>水氮>氮磷,蒙古冰草为水氮>氮磷>水磷。
(3) 草木樨的光合及生物量均远大于蒙古冰草,西北煤矸石山植被恢复的草本植物首选草木樨,其最优水肥方案为:土壤相对含水量68.44%~71.84%,施氮量82.09~102.92 kg/hm2,施磷量139.92~192.58 kg/hm2。其次是蒙古冰草,其最优水肥方案为:土壤相对含水量65.33%~71.11%,施氮量138.40~172.62 kg/hm2,施磷量82.09~117.91 kg/hm2。
猜你喜欢
莫愁(2023年25期)2023-09-14 06:34:56
天津农林科技(2022年1期)2022-02-12 03:55:16
草原与草坪(2021年6期)2022-01-24 07:17:44
中国草食动物科学(2018年2期)2018-05-25 06:10:43
新课程·下旬(2017年7期)2017-08-14 11:12:50
草业科学(2016年12期)2017-01-11 07:21:38
西北林学院学报(2015年5期)2015-01-03 05:12:20
草食家畜(2012年2期)2012-03-20 13:22:39
草食家畜(2012年2期)2012-03-20 13:22:33
植物营养与肥料学报(2011年2期)2011-10-26 03:52:30
