
河流大型底栖动物对环境压力的响应:以太湖、巢湖流域为例*
2021-01-12 06:58蔡永久高俊峰李宽意
湖泊科学 2021年1期
张 又,蔡永久,张 颖,高俊峰,李宽意,3
(1:中国科学院南京地理与湖泊研究所,湖泊与环境国家重点实验室,南京 210008) (2:江苏省水利工程规划办公室,南京 210029) (3:中国科学院大学中丹学院,北京 100049)
大型底栖动物是淡水生态系统的一个重要生物类群,是物质循环和能量流动的主要环节,有着承上启下的关键作用. 底栖动物可以促进有机碎屑分解,调节泥-水界面的物质交换,维持生物完整性等,在淡水生态系统中发挥着重要的作用[1-2]. 底栖动物具有相对较长的生命周期、活动范围小,能够综合反映较长时间段内的环境质量[3];不同物种对环境变化的敏感程度不同,如蜉蝣目物种对水体污染较为敏感,而寡毛类则为中污染和重污染的指示物种[4-5];当水体受到污染时,底栖动物群落结构也随之发生变化,底栖动物的群落结构、优势种类和密度等生物指标可以反映不同类型环境压力的影响[6]. 因此,底栖动物常被用于生物监测和水质评价[7-8].
“生态区”的概念由Crowly于1967年首次提出[9]. 生态区是指在地理空间上具有独特的自然群落、物种和环境条件组合特征,同时具有相对一致的水生态系统组成、结构、格局、过程和功能的水体或具有“水陆一致性”的陆水联合体[10-11]. “十一五”以来,我国系统地开展了流域水生态功能分区理论与方法研究,建立水生态功能分区指标体系,建立全国水生态功能分区技术框架[12],主要以流域为单元根据环境要素、水生态系统特征及生态服务功能在不同地域的差异性和相似性开展了水生态功能区划分工作[13]. 流域水生态功能分区不仅反映了水生态空间分布格局的差异,也在一定程度上考虑了人类活动对水生态系统的影响,实现了自然要素与功能要素的结合,提出了面向水生态保护的管理区域,推动了从水环境管理向水生态管理理念的转变[14]. 生态区自然环境特征(如海拔、水文特征等)的变化,是决定区域内生物群落特征的关键因素. 因此,以生态区为空间单元来研究河流底栖动物群落结构的分布特征,可以有效协助淡水生态系统的管理工作[15-16]. 欧盟在生态区的基础上建立了以水生生物指标为核心的生态质量评价方法,通过“压力-状态-响应”关系分析,判断水体受损状况,分别针对受损水体和未受损水体明确保护目标[17]. 我国以生态区为空间单元来分析底栖动物群落的研究较少. 孟云飞等[18]以水生态功能区为单元研究了浑太河流域大型底栖动物群落特征,发现不同水生态功能区影响大型底栖动物群落结构的环境因子存在差异性. 张又等[19]对太湖流域的研究发现,西部丘陵区和东部平原区的底栖动物优势种差异明显,且两个生态区受污染程度差异较大.
中国经济与社会的发展迅速,正处于全面建成小康社会决胜阶段,农业稳定增长,农业现代化深入发展,工业化水平进一步提高,经济的地域性差异,给不同地区的生态环境造成了不同类型和不同程度的影响. 对于不同的地域,工农业发展的侧重和发展程度的差异都在很大程度上决定了人为干扰对该地区环境的影响方式和程度. 太湖流域和巢湖流域是较早完成水生态功能区划分的流域[20-21],且两个流域的土地利用类型差异较大,分别以工业发展和农业发展为主,水生态系统造成的环境压力不同. 因此,本研究以太湖流域和巢湖流域为研究区,对比分析两流域丘陵区和平原区的水环境与栖息地质量差异、底栖动物群落差异及其主要驱动因素;综合考虑两个流域的理化环境和底栖动物群落组成,分析底栖动物特征种对关键环境胁迫因子的响应关系,以期为流域水生态保护目标的确定提供科学依据.
1 材料与方法
1.1 研究区域与采样点布设
太湖流域(30°7′19″~32°14′56″N,119°3′1″~121°54′26″E)位于长江三角洲地区,北抵长江,东临东海,南滨钱塘江,西以天目山、茅山为界,面积3.69×104km2,地势中部低洼,总体呈现西部高东部低的趋势. 流域内大小湖泊星罗棋布,以太湖为中心,河流与湖泊相互交错,呈水网状. 流域河道总长12万km,河网密度达3.25 km/km2,是我国河网密度最大的地区[19,22].
巢湖流域(31°25′28″~31°43′28″N,117°16′54″~117°51′46″E)属长江水系,流域面积约1.3×104km2. 巢湖流域地势的总轮廓是东西向较长,地形总体北高南低,西高东低,中间低洼平坦. 西南为山区,东北为丘陵及浅山区,东南及沿湖为平原. 流域内河网密布、纵横交错,主要支流发源于大别山区,共33条出入湖河流,呈放射状直接或间接汇入巢湖[23].
本研究在太湖流域和巢湖流域分别布设93和147个采样点,其中太湖流域西部丘陵区(E1)和东部平原区(E2)布设了22和71个采样点,巢湖流域西部丘陵区(e1)和东部平原区(e2)分别布设了47和100个采样点,涵盖了生态区内的主要溪流和河流(图1).
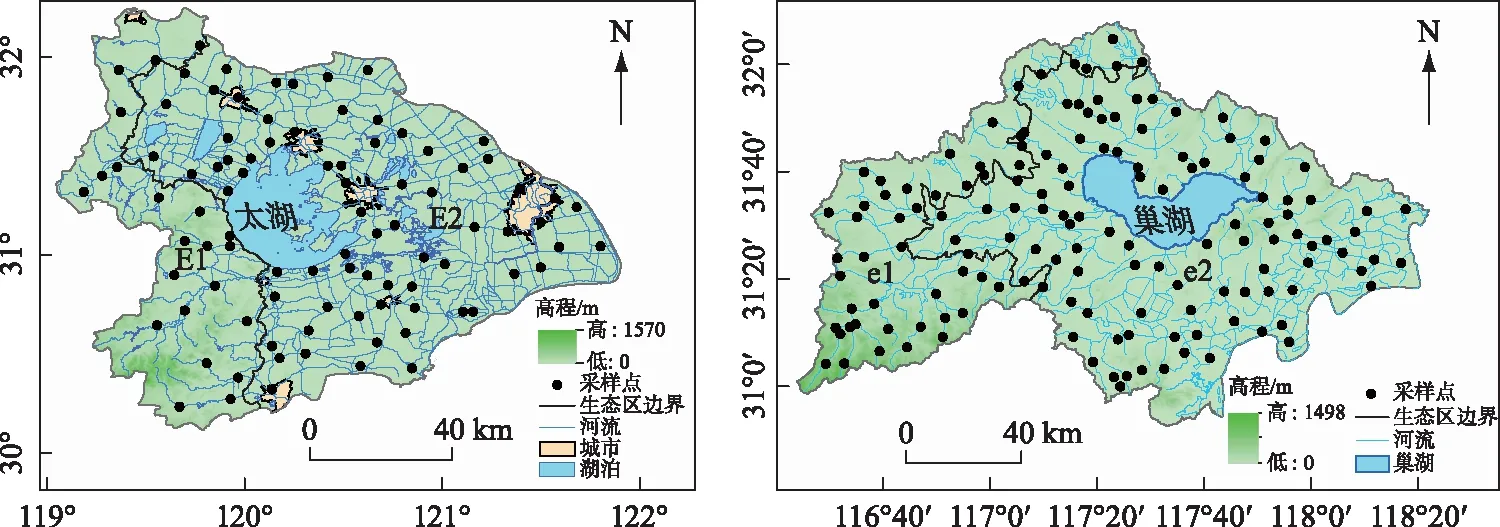
图1 太湖流域和巢湖流域采样点分布Fig.1 Sampling sites of Taihu Basin and Chaohu Basin
1.2 底栖动物样品采集和鉴定
太湖流域底栖动物样品采集时间为2012年10月,巢湖流域底栖动物样品采集时间为2013年4月. 大型底栖动物样品采用D型抄网或彼得逊采泥器采集. 对于可以涉水的溪流及小型河流,采用D形抄网进行底栖动物样品采集,每个采样点采集10个样方,包含100 m河段内的主要生境,每个样方的大小为30 cm × 50 cm. 每个样点的10个样方混合成一个样品. 对于无法涉水的大型河流,采用1/16 m2的彼得逊采泥器进行底栖动物样品采集,每个样点采集6次,并混合成一个样品. 每个样点所采集的样品,需经过60目尼龙筛网进行现场筛洗,剩余物带回实验室,置于白色瓷盘中,将所有底栖动物逐一挑出,并用7%福尔马林溶液固定保存.
底栖动物的鉴定需在解剖镜和显微镜下进行,所有样品都鉴定至尽可能低的分类单元[24-27]. 每个采样点所采到的底栖动物按不同种类准确地统计数量并用电子天平称重,称重前需用滤纸吸去表面固定液. 软体动物带壳称重. 根据每个样点的采样面积换算出每个样点的每种底栖动物的密度(ind/m2)和生物量(g/m2).
1.3 水环境与栖息地调查
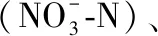
通过现场调查,根据质量状况优劣程度,将栖境多样性(Habitat)、河道变化(Channel)、交通干扰程度(Transportation)、河岸带土地利用(Land)、底质异质性(Substrate)5个反映生境质量的参数分为:好(16~20)、较好(11~15)、一般(6~10)和差(0~5)4个等级[19,29-30],具体的打分标准见表1.
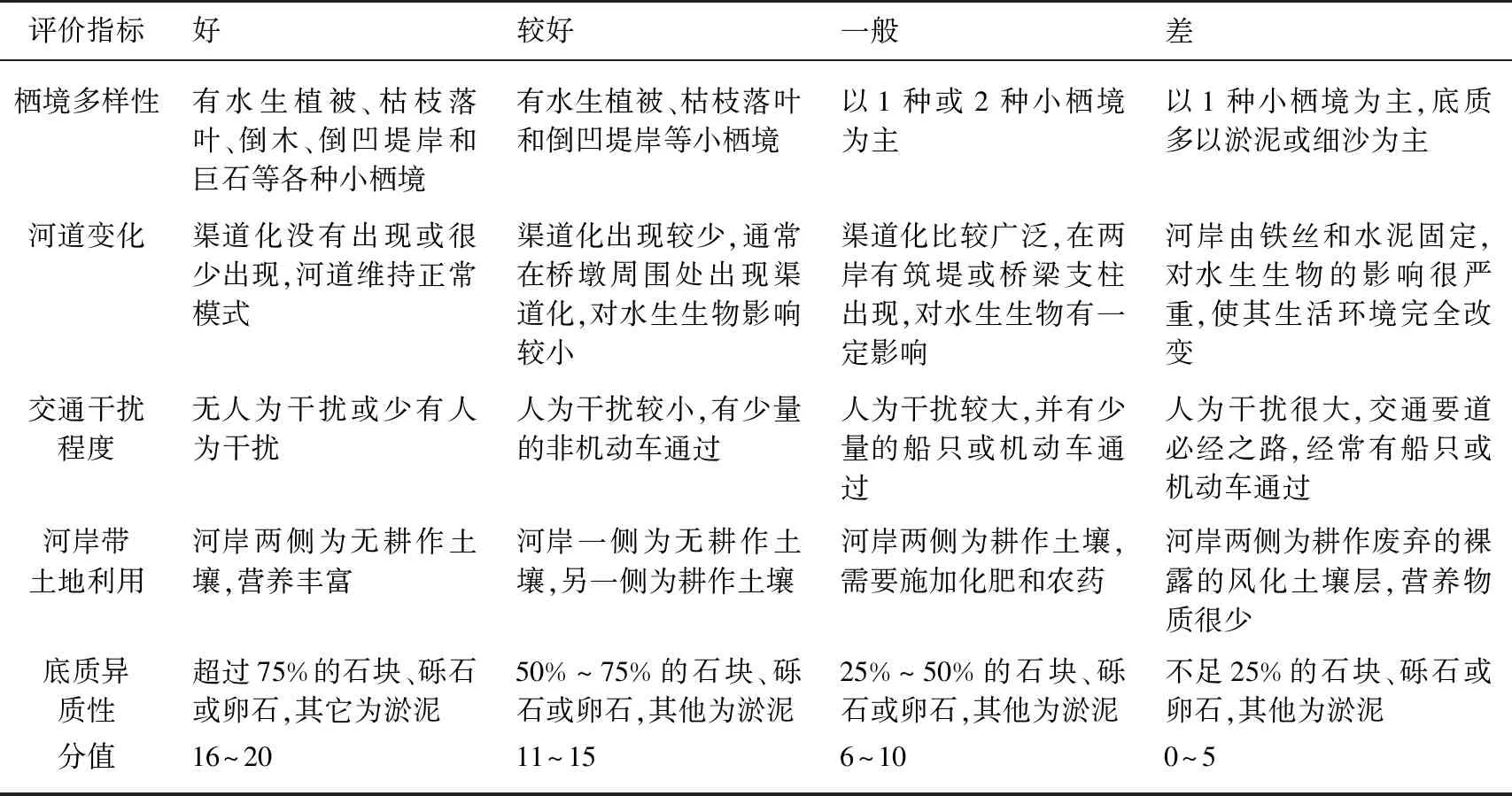
表1 生境质量评价指标及标准
1.4 数据分析处理
采用主成分分析(PCA)分别对两个流域的丘陵区和平原区的环境因子排序,从而提取出能够尽可能多地反映原来变量信息的综合变量,寻找研究区环境梯度. 采用相似性分析(analysis of similarities,ANOSIM)检验两个流域丘陵区和平原区的底栖动物群落组成的差异显著性,并利用相似百分比分析(similarity percentage,SIMPER)找出对两流域丘陵区和平原区底栖动物群落差异贡献最大的种类及其对差异的贡献率[31]. 同时采用ANOSIM和SIMPER分析两个流域丘陵区和平原区的环境差异,找出对差异贡献最大的水环境与栖息地指标,及其对差异的贡献率.
采用典范对应分析(canonical correspondence analysis,CCA)分别分析两流域丘陵区和平原区的底栖动物群落与环境因子之间的关系,用CCA排序图将物种、样点和环境因子绘出,直观地呈现出种类组成及群落分布与环境因子之间的关系[32]. 分析时,物种数据进行平方根转换,环境因子进行lg(x+1)转换(pH值除外). 环境因子的筛选采用向前引入法,保留能通过蒙特卡罗置换检验的显著因子(P<0.1).
广义加性模型(generalized additive models,GAM)是适用于响应变量与解释变量呈非线性关系的分析方法[33]. GAM模型对数据的分布限制较少,可以是高斯分布、二项式分布、泊松分布及其他复杂的分布等,尤其适用于某些离散数据,因此GAM模型可以较好地分析因变量与多自变量之间的非线性关系[34]. 因CCA只能总体确定影响底栖动物群落结构的环境因子,但不同物种对环境因子的响应模式是不同的,为明确特征种和多样性对环境因子的响应关系,本研究采用GAM模型,以泊松分布类型及AIC(Akaike information criterion)最小为最优的原则筛选并拟合底栖动物特征种与环境因素的响应关系,确定底栖动物特征种响应关系显著(P< 0.05)的环境因子,定量地解析底栖动物对环境压力的响应模式.
本研究的PCA、CCA和GAM分析在CANOCO 4.5软件中完成,ANOSIM和SIMPER分析在PRIMER 5.0软件中完成.
2 结果与分析
2.1 水环境与栖息地质量
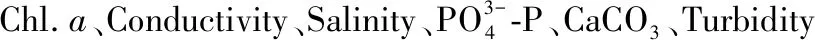
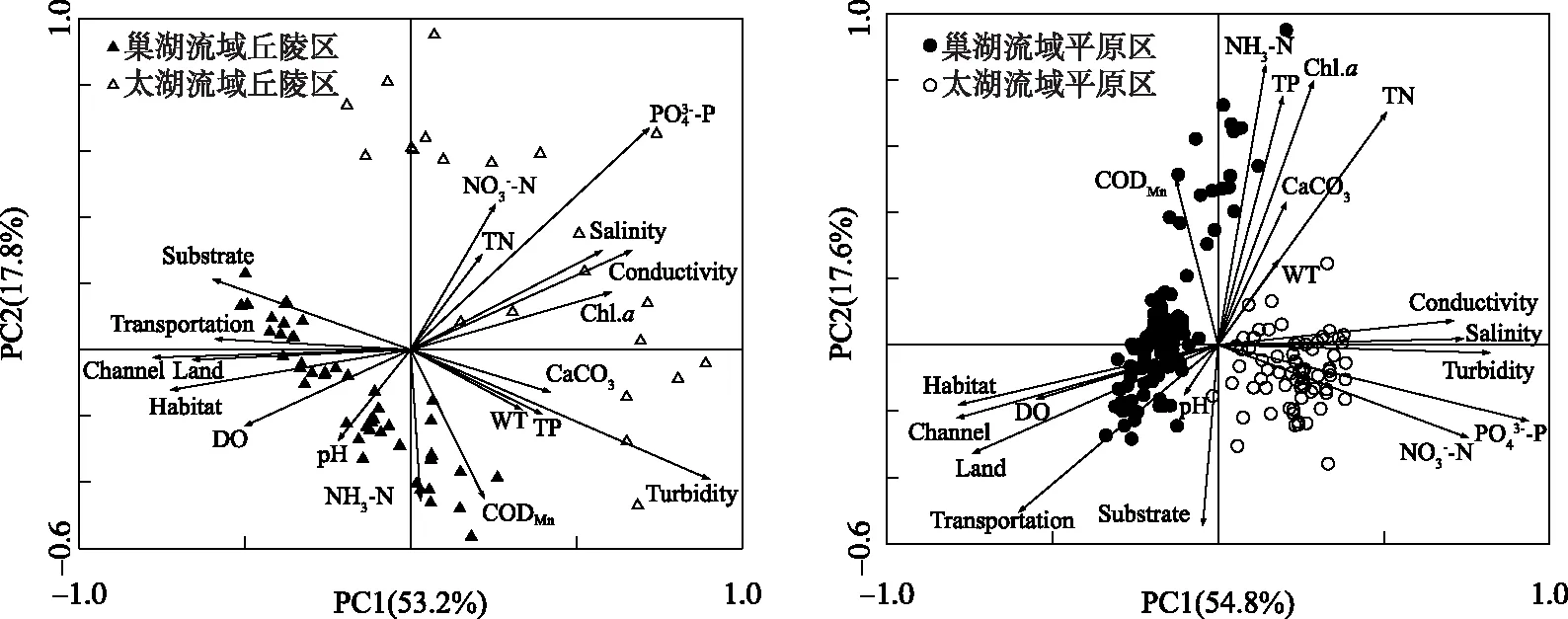
图2 水环境与栖息地质量的主成分分析排序图Fig.2 Principal component analysis of water quality and habitat quality factors
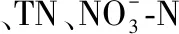
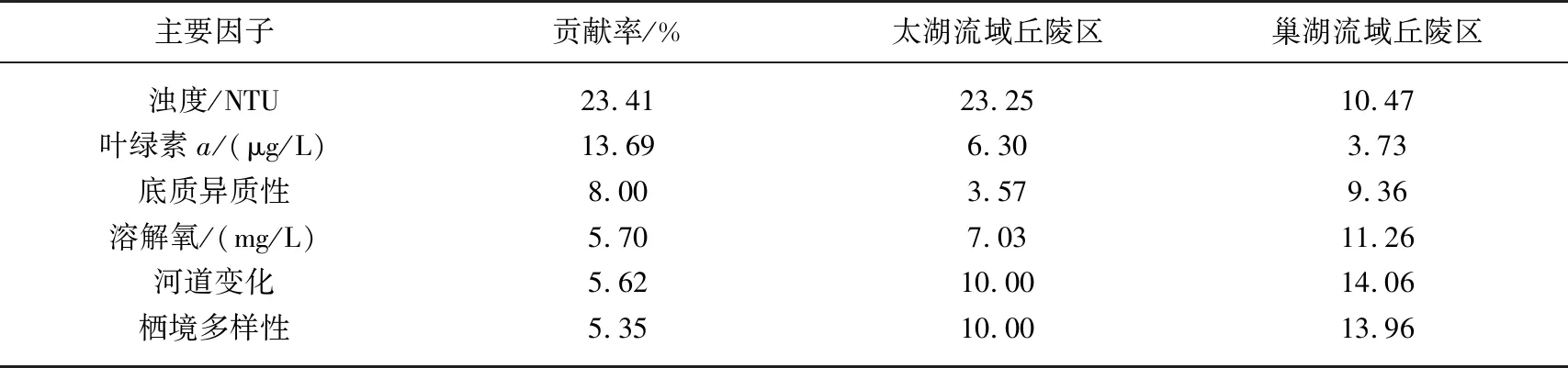
表2 两个流域丘陵区的环境特征差异的主要因子及其均值*
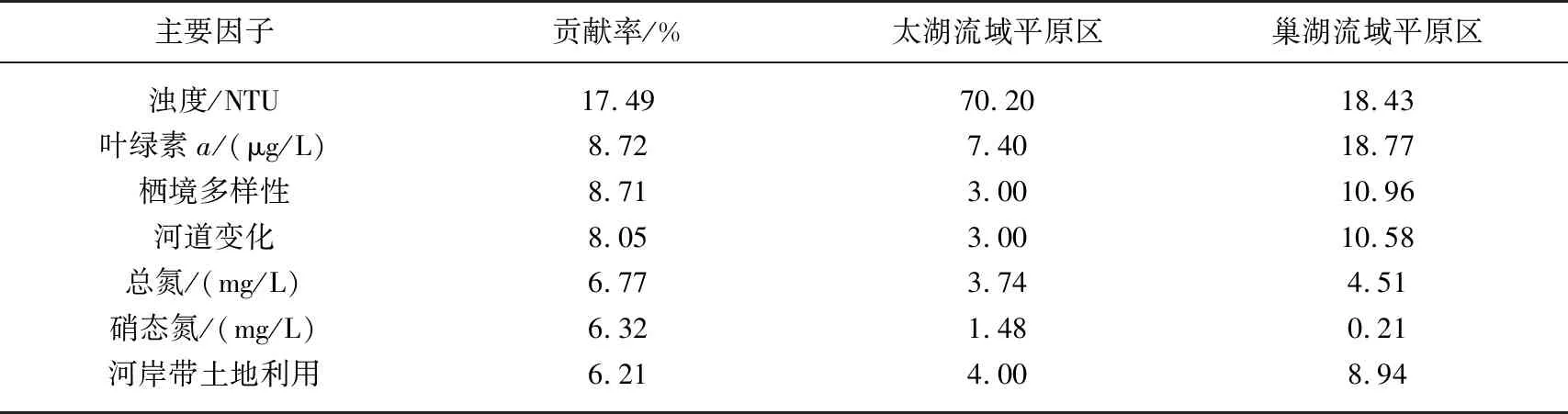
表3 两个流域平原区的环境特征差异的主要因子及其均值*
2.2 大型底栖动物群落结构
One-way ANOSIM结果显示,巢湖流域丘陵区与太湖流域丘陵区的底栖动物群落结构差异显著(P< 0.01,R=0.133),巢湖流域平原区与太湖流域平原区的底栖动物群落结构差异显著(P<0.01,R=0.724).
两个丘陵区底栖动物群落结构的差异物种主要为腹足纲(8种)、水生昆虫(5种)、软甲纲(3种)、双壳纲(3种)、寡毛纲(2种)及摇蚊幼虫(3种)(表4). 从平均密度来看,巢湖流域丘陵区的水生昆虫密度远高于太湖流域丘陵区,太湖流域丘陵区的寡毛纲和软甲纲平均密度稍高于巢湖流域丘陵区,而摇蚊幼虫和双壳纲在两个流域丘陵区的平均密度相差不大. 虽然铜锈环棱螺在两个流域丘陵区的平均密度接近,但总体来讲,巢湖流域丘陵区的腹足纲密度高于太湖流域丘陵区.
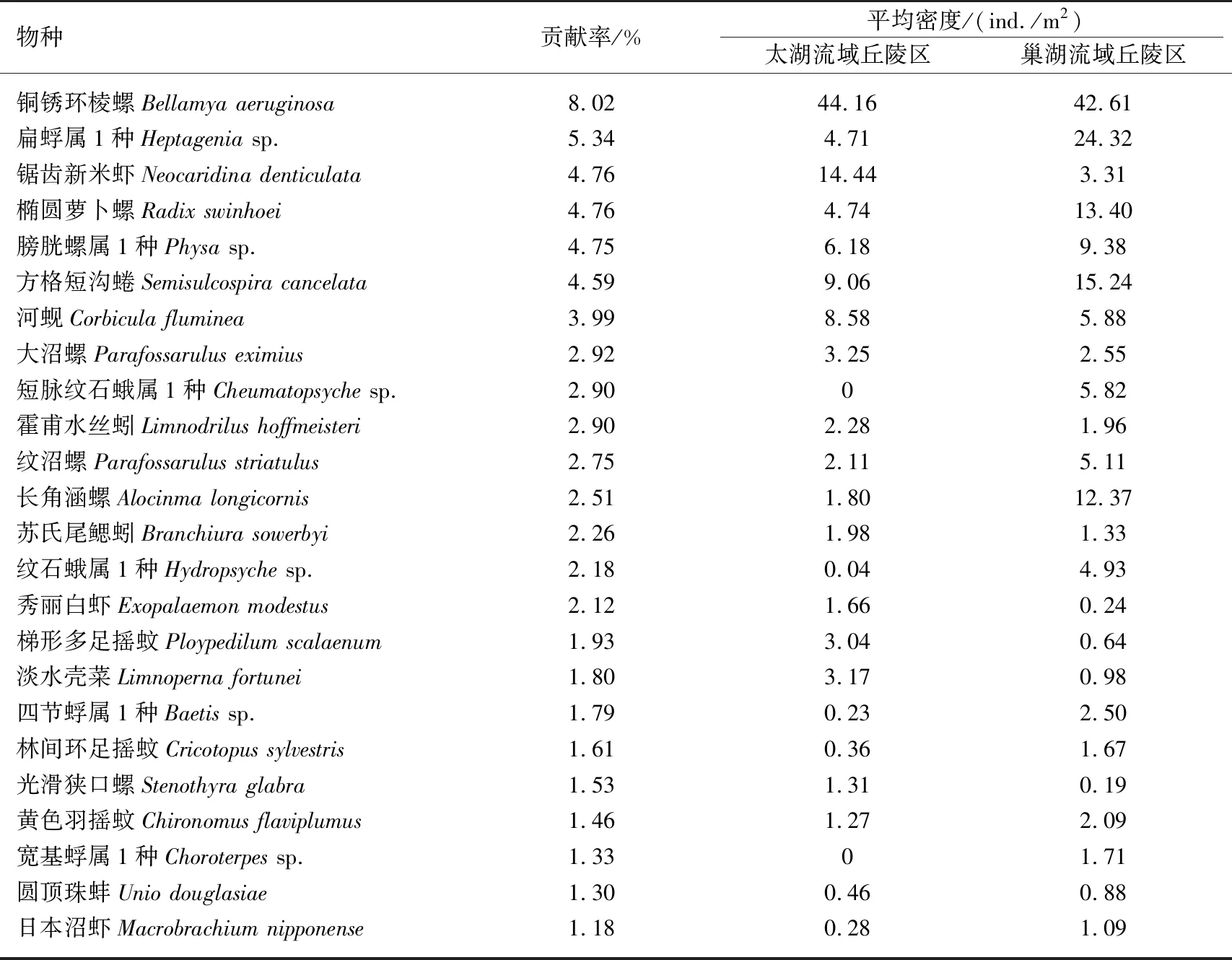
表4 两个流域丘陵区底栖动物群落差异的主要物种对组间差异的贡献率及其密度均值*
两个平原区底栖动物群落结构的差异物种主要为腹足纲(7种)、软甲纲(2种)、双壳纲(2种)、寡毛纲(2种)及摇蚊幼虫(2种)(表5). 从平均密度来看,巢湖流域平原区的寡毛纲(霍甫水丝蚓和苏氏尾鳃蚓)平均密度远高于太湖流域平原区. 腹足纲的铜锈环棱螺、大沼螺、纹沼螺、方格短沟蜷在太湖流域平原区的平均密度较高,而长角涵螺、椭圆萝卜螺、尖口圆扁螺在巢湖流域平原区的平均密度较高. 巢湖流域平原区双壳纲和摇蚊幼虫的平均密度稍高于太湖流域平原区. 而软甲纲在两个流域平原区的平均密度相差不大.
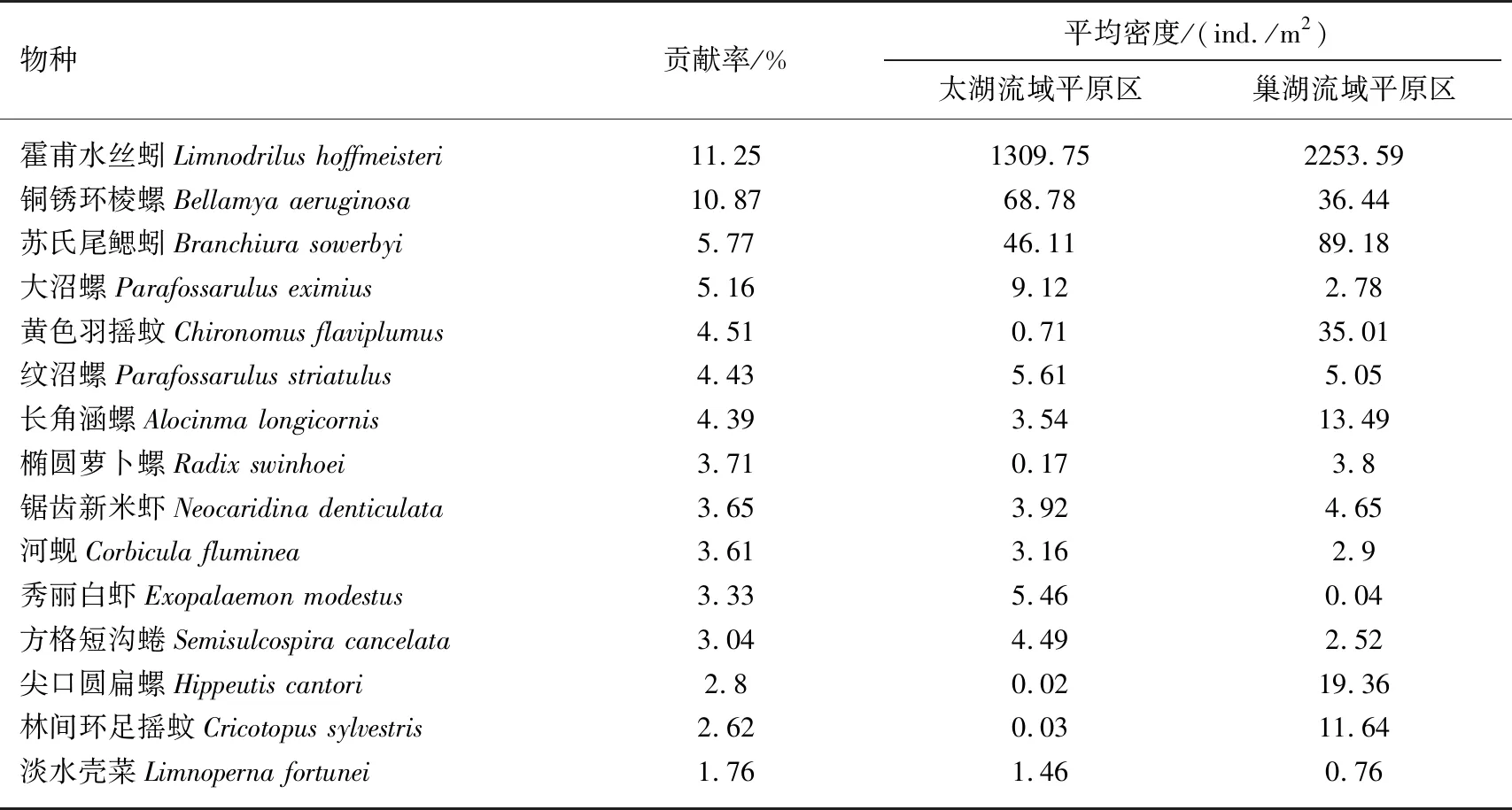
表5 两个流域平原区底栖动物群落差异的主要物种对组间差异的贡献率及其密度均值*
2.3 底栖动物群落结构的驱动因素
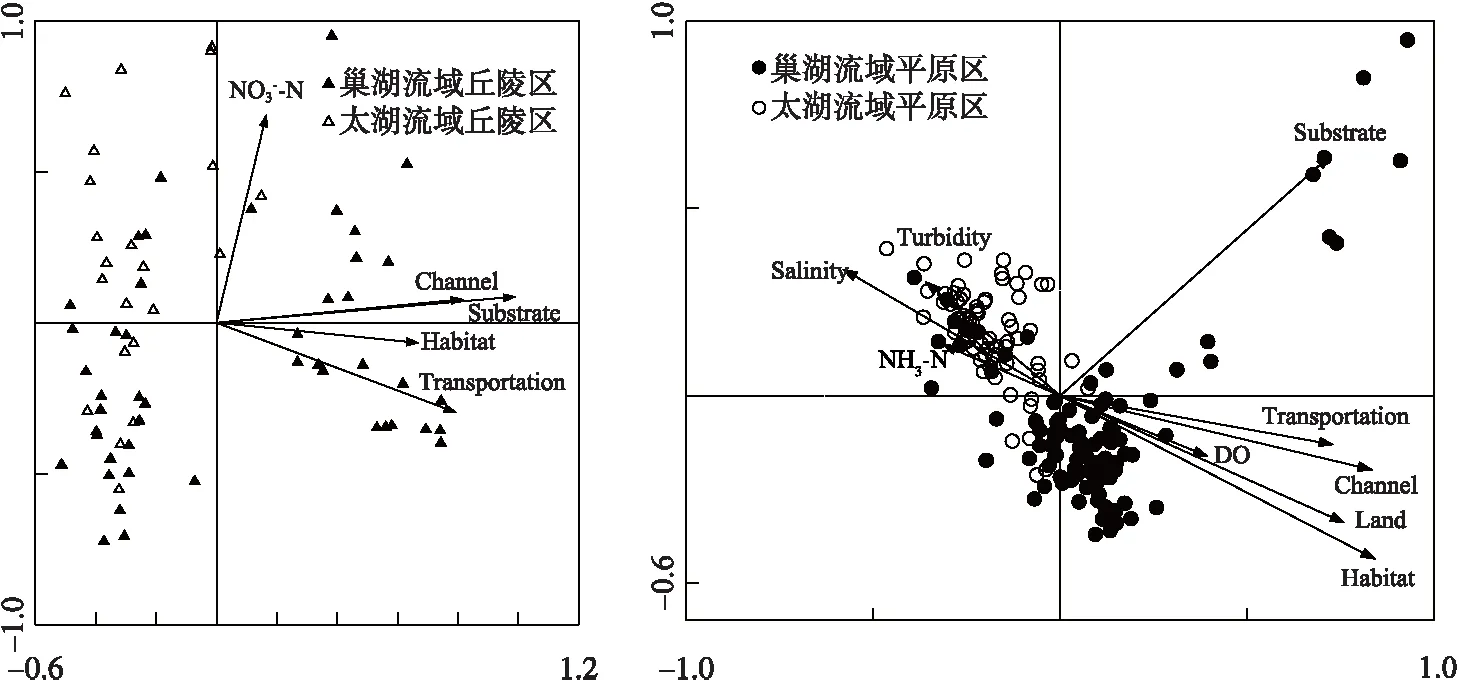
图3 底栖动物群落结构与环境因子的典范对应分析排序图Fig.3 Canonical correspondence analysis of macroinvertebrate community and environmental variables
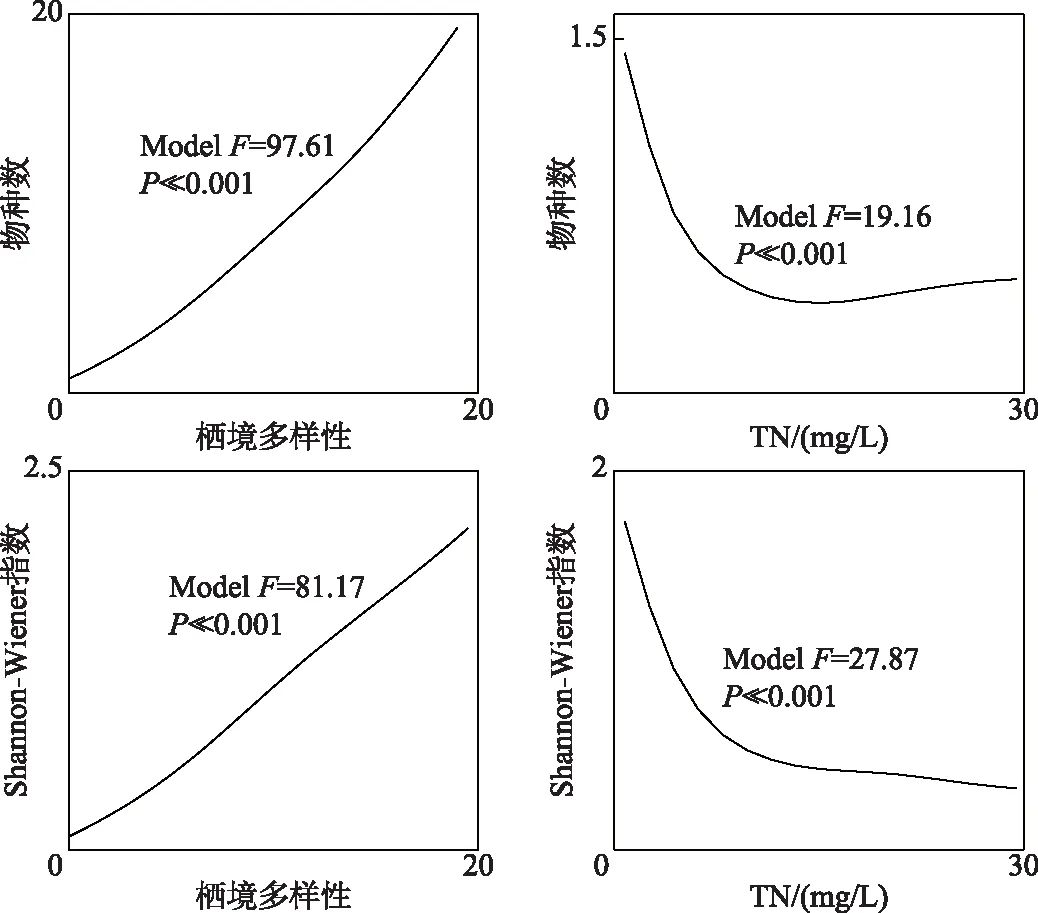
图4 底栖动物物种数及Shannon-Wiener多样性指数对关键环境因素的响应曲线Fig.4 Response curves of richness and Shannon-Wiener index to the key environmental factors
2.4 大型底栖动物对关键环境因素的响应
2.4.1 底栖动物多样性对关键环境因素的响应 底栖动物物种数和Shannon-Wiener指数与水体TN浓度和栖境多样性的响应相关性显著(P≪0.001),两者随着栖境多样性的升高而稳步上升(图4). Shannon-Wiener指数随着水体TN浓度的升高呈下降趋势,当TN浓度处于5~10 mg/L时,达到阈值浓度,指数处于相对较低的水平(< 0.5). 类似地,物种数随着水体TN浓度的升高急速下降;当TN浓度大于5 mg/L时,物种数的下降速度趋缓并处于较低水平.
2.4.2 底栖动物特征种对关键环境因素的响应 铜锈环棱螺和椭圆萝卜螺作为腹足纲的特征种,对栖境多样性、底质异质性和电导率的响应关系显著(P< 0.05),但两个特征种对上述3个环境因子的响应关系相差较大(图5). 铜锈环棱螺密度随着栖境多样性的提高呈先上升后下降的趋势,而椭圆萝卜螺密度随着栖境多样性的提高呈上升趋势. 铜锈环棱螺密度随着底质异质性的升高逐渐降低,而椭圆萝卜螺对底质异质性的响应呈单峰关系. 铜锈环棱螺对电导率的响应呈现单峰关系,在电导率为0.6 mS/cm时达到最大. 椭圆萝卜螺密度随着电导率的升高而降低,但在电导率大于0.3 mS/cm以后下降速度趋缓.
霍甫水丝蚓和苏氏尾鳃蚓作为寡毛纲的特征种,对电导率、CODMn和TP浓度的响应关系显著(P≪0.001),且响应关系类似(图6),其对电导率的响应均为典型的单峰关系,均在电导率约为0.7 mS/cm时达到峰值,电导率继续增加时,苏氏尾鳃蚓密度下降更快. 两特征种对CODMn和TP浓度的响应关系相似,密度均随着指标值的增大而增大. 此外,响应曲线的结果还显示:相同环境条件下,霍甫水丝蚓的密度远高于苏氏尾鳃蚓.
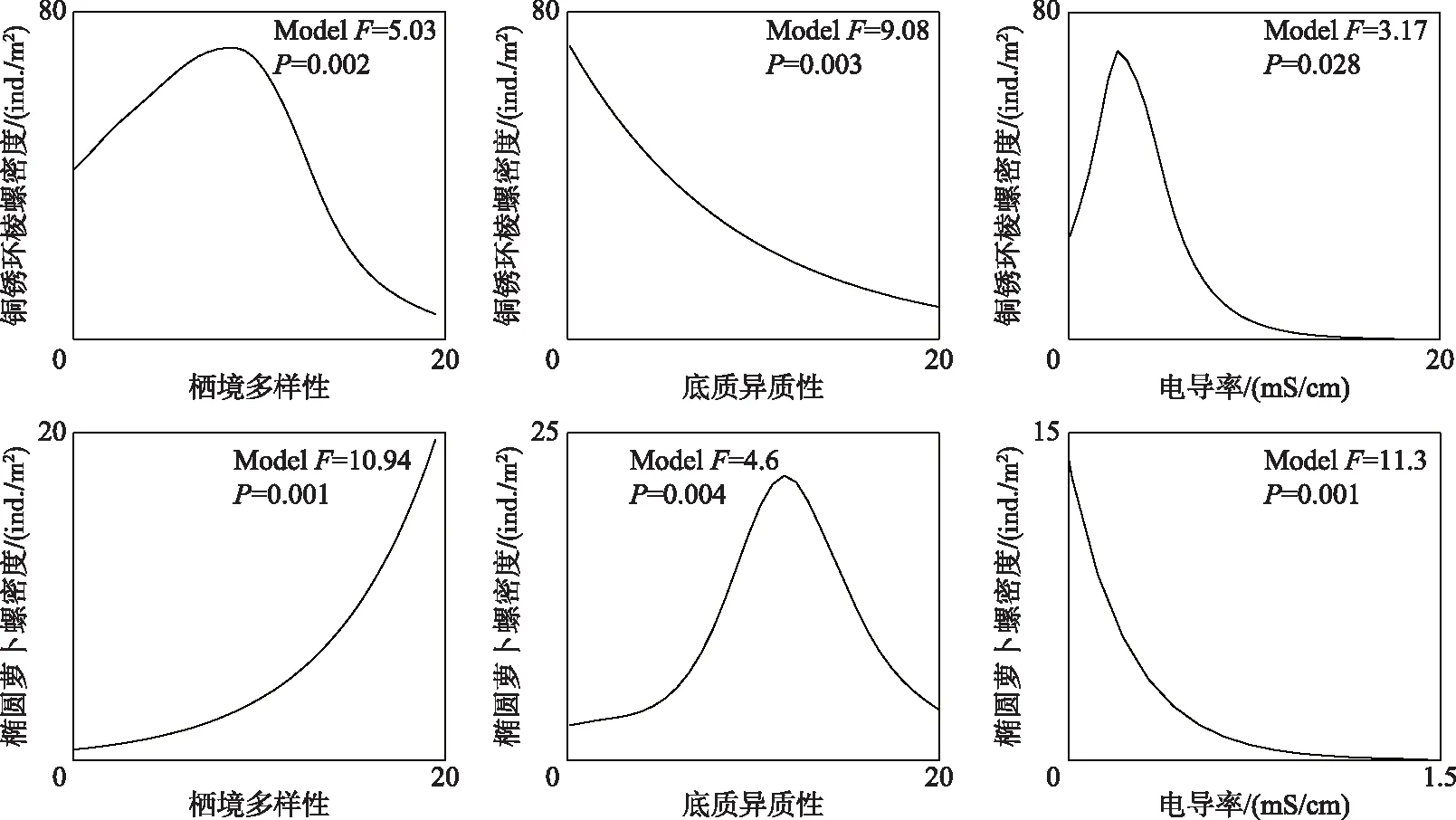
图5 腹足纲特征种对关键环境因素的响应曲线Fig.5 Response curves of characteristic species of Gastropod to the key environmental factors
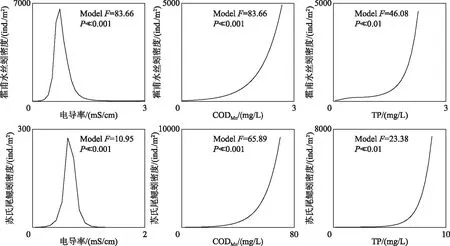
图6 寡毛纲特征种对关键环境因素的响应曲线Fig.6 Response curves of characteristic species of Oligochaeta to the key environmental factors
双壳纲特征种河蚬对底质异质性(P<0.01)和TN浓度(P<0.05)的响应关系显著(图7). 河蚬对底质异质性的响应大致呈单峰关系,在底质异质性约为8时达到峰值. 河蚬密度随着TN浓度的升高而急速降低,但当TN浓度大于3 mg/L时,其下降速度趋缓.
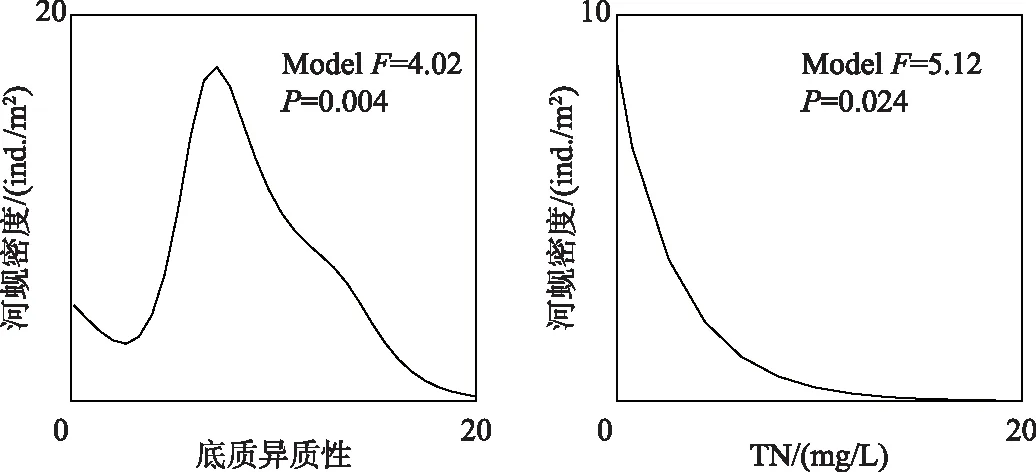
图7 双壳纲特征种河蚬对关键环境因素的响应曲线Fig.7 Response curves of Corbicula fluminea (Bivalvia) to the key environmental factors
摇蚊幼虫特征种黄色羽摇蚊对栖境多样性、CODMn和TN的响应关系显著(P<0.001)(图8). 黄色羽摇蚊的密度会随着CODMn的升高而增加,而对栖境多样性和TN浓度的响应均为典型的单峰关系,在栖境多样性约为6或TN浓度约为42 mg/L时达到峰值.
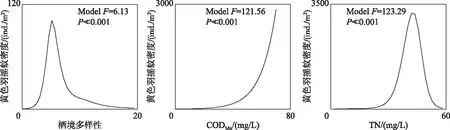
图8 摇蚊幼虫特征种黄色羽摇蚊对关键环境因素的响应曲线Fig.8 Response curves of Chironomus flaviplumus (Chironomidae) to the key environmental factors
研究区的两个水生昆虫类特征种对不同的环境因素响应关系较高:扁蜉属对DO(P≪0.001)和CODMn(P≪0.001)的响应关系显著,而四节蜉属对电导率(P≪0.001)和TN(P< 0.05)的响应关系明显(图9). 扁蜉属在DO浓度低于6 mg/L的环境中基本无法生存;在DO浓度高于6 mg/L的情况下,扁蜉属密度随着DO的升高而上升. 扁蜉属密度随着CODMn的升高而下降,CODMn超过2.1 mg/L时,其下降速度趋缓,而当CODMn大于4.2 mg/L时,扁蜉属几乎不出现. 四节蜉属密度随着电导率和TN浓度的升高而下降:在电导率大于0.3 mS/cm的情况下,四节蜉属的密度趋于0;在TN浓度大于6 mg/L的环境中,四节蜉属已经很难生存.
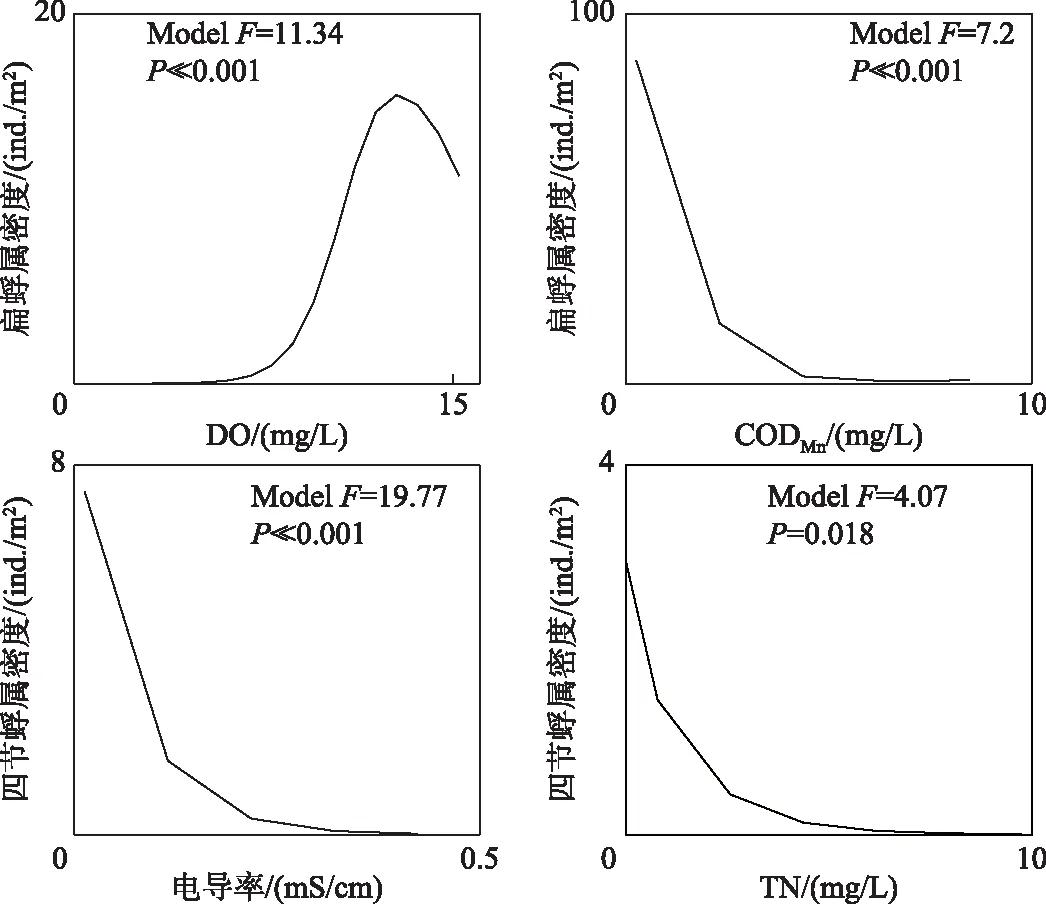
图9 水生昆虫特征种对关键环境因素的响应曲线Fig.9 Response curves of characteristic species of aquatic insects to the key environmental factors
3 讨论
3.1 底栖动物群落结构特征差异分析
巢湖流域大型底栖动物群落结构与太湖流域差异较大,巢湖流域丘陵区生境质量(栖境多样性、河道变化、交通干扰程度和底质异质性)优于太湖流域丘陵区. 与人类干扰强度更大的太湖流域相比,巢湖流域大型底栖动物的物种丰富度更高,各采样点底栖动物的密度、物种数及多样性指数也更高. 巢湖流域是一个农业主导型的湖泊型流域,其河道以自然土质护坡为主. 而太湖流域的很多河流为硬质护坡,流域内河流渠道化程度较高,生境退化严重,这进一步证明了城市化和河道渠道化会严重影响大型底栖动物的结构特征[4]. 巢湖流域大型底栖动物的物种丰富度水平和生物多样性水平与土地利用同样以农业为主的辽河流域[35-36]相似,但辽河流域源头溪流中的清洁种,特别是四节蜉属昆虫(Baetissp.)的数量较多,而在第4级河流的点位中摇蚊幼虫、软甲纲等耐污性物种更占优势[35]. 此外,辽河流域以水生昆虫为主的清洁种主要分布在以林地为主的区域中,在以耕地和居民住宅为主的区域摇蚊幼虫占据优势,而以城市为主的区域寡毛纲的优势地位明显[36],说明人类活动干扰程度和土地利用类型对底栖动物群落结构空间分异影响最显著.
我国不同管理部门根据其管理需求采用的分区方式不同,如水功能区、生态功能区和主体功能区等,这些分区方式与流域水生态功能区的分区目的、分区体系、分区方法等均不相同[13]. 水生态功能分区主要是面向水生态系统,以小流域为基本单元,考虑水生态系统的完整性和生物区系,揭示不同区域水生生物空间分布格局,而水功能区则注重水体使用功能的划分,难以反映水生态特征及水生态功能的差异. 生态功能区是以陆地生态系统为基础的,也不适用水生态系统的管理需求. 本研究中发现,两流域的丘陵区和平原区的特征种组成也有很大差异. 丘陵区排在前5位的特征种为铜锈环棱螺、扁蜉属、锯齿新米虾、椭圆萝卜螺和膀胱螺属,均为耐污和清洁的种类,而平原区排在前5位的特征种为霍甫水丝蚓、铜锈环棱螺、苏氏尾鳃蚓、大沼螺和黄色羽摇蚊,均为耐污和中等耐污的种类,说明水生态功能分区可以很好地区分底栖动物群落特征.
3.2 底栖动物-环境压力响应曲线的应用
水质指标是水环境质量评价的最常用方法,但只能代表采集时的水质状况,并不能反映人类活动对水环境的长期效应[37]. 此外,人为干扰不仅仅体现在水质状况上,生境质量下降也是人类活动压力的重要体现,在研究水生生物群落响应时也应予以充分考虑. 底栖动物群落特征对水质状况及生境质量的响应具有时间累积效应,其监测可以真实地反映过去一段时间内的环境状况变化. 因此,建立底栖动物群落结构特征与环境梯度响应关系,对环境监测和淡水生物多样性保护均具有积极作用.
本研究基于GAM筛选并拟合了大型底栖动物对关键环境梯度的响应曲线,结果表明底栖动物物种丰富度、生物多样性及特征种与不同的环境因子具有显著响应关系. 栖境多样性和TN浓度可以作为底栖动物生物多样性和物种丰富度的指示因子. 铜锈环棱螺和椭圆萝卜螺对电导率、底质和栖境多样性较为敏感,但两者的响应关系曲线不同. 这主要是因为两者生活习性不同,椭圆萝卜螺属肺螺亚纲,适合有水生植物分布的水域;而铜锈环棱螺属前鳃亚纲,有厣和鳃,以底栖着生藻和有机碎屑等为食,适应性较强,生态位较宽[38]. 研究还表明,河蚬对底质异质性的响应关系呈单峰曲线,在底质异质性为8左右达到最大,说明河蚬偏好的沙泥混合底质;河蚬的密度随着TN浓度的升高而极速降低,这可能是因为该物种对环境变化较敏感,可以作为水质清洁的指示物种. 霍甫水丝蚓和苏氏尾鳃蚓可以在TP浓度较高且DO浓度极低的严重污染环境中生存. 黄色羽摇蚊可以在低DO的严重污染水体大量富集,但在TN浓度大到一定程度时,其密度也会急速下降. 特别地,霍甫水丝蚓、苏氏尾鳃蚓和黄色羽摇蚊对CODMn的响应曲线均呈指数增长模式,说明这3个特征种的耐污能力极强,可以在有机污染严重的环境中生存并大量富集[39]. 水生昆虫的各特征种丰度随着DO的升高而升高,且随着CODMn、TN和电导率的升高而下降,说明该物种类群对生境质量下降和水质降低特别敏感,较常出现在清洁的水体中. 这些结果都表明,以上这些特征物种可以作为环境监测的指示物种. 底栖动物-环境梯度的响应曲线能够更加清晰地描述底栖动物群落对环境因子的响应关系,有利于深入地了解水质、营养状态及生境质量与大型底栖动物群落结构的相关关系,并进一步分析大型底栖动物群落结构的变化趋势和演替过程.
4 结论
不同人类干扰强度河流的底栖动物群落结构差异显著,且底栖动物物种丰富度、生物多样性和特征种对不同环境因子的响应不同. 巢湖流域丘陵区和平原区的生境质量均优于太湖流域,但巢湖流域平原区部分点位的水体营养盐极高于太湖流域平原区. 两个流域的大型底栖动物群落组成差异较大,两个丘陵区的差异物种较两个平原区更多,丘陵区敏感型物种(水生昆虫)的种类更多. 巢湖流域丘陵区的水生昆虫密度远高于太湖流域丘陵区,太湖流域丘陵区的寡毛纲平均密度稍高于巢湖流域丘陵区,而巢湖流域平原区的寡毛纲平均密度远高于太湖流域平原区,这些特征与理化结果是一致的. 基于广义加性模型拟合的响应曲线表明,大型底栖动物物种丰富度和生物多样性对栖境多样性和TN浓度的响应关系显著,不同物种类群对不同的环境因子响应关系显著. 霍甫水丝蚓、苏氏尾鳃蚓、黄色羽摇蚊可作为水质重污染的指示种,椭圆萝卜螺可作为水质中污染的指示种,而河蚬和扁蜉属、四节蜉属等水生昆虫可作为水质清洁的指示种,可为生物监测提供参考.
猜你喜欢
现代经济信息(2022年25期)2022-11-09
资源节约与环保(2022年8期)2022-09-20
河北地质(2022年2期)2022-08-22
环境工程技术学报(2022年3期)2022-06-05
恋爱婚姻家庭(2020年25期)2020-09-08
恋爱婚姻家庭(2020年9期)2020-09-07
安徽园林(2018年3期)2018-10-09
科技资讯(2018年30期)2018-03-11
江苏农业科学(2015年11期)2016-01-27
湖北农业科学(2014年6期)2014-07-02
