
梵净山保护区主要雉类的繁殖期栖息地选择与空间分布*
2021-01-05 02:47杨朝辉粟海军胡灿实张明明
林业科学 2020年11期
王 丞 冉 伟 杨朝辉 毕 兴 粟海军 胡灿实 石 磊 张明明
(1. 贵州大学林学院 贵州大学生物多样性与自然保护研究中心 贵阳 550025; 2. 贵州梵净山国家级自然保护区管理局 江口 554400; 3. 生态环境部南京环境科学研究所 南京 210042)
目前, 中国有63种鸡形目(Galliformes)鸟类正面临着栖息地丧失、捕杀等威胁, 其中,绝大多数物种(49种)已被列入《中国脊椎动物红色名录》受威胁等级(蒋志刚等, 2016)。自然保护区作为维持生物多样性保护的重要屏障, 为野生动物提供了重要栖息地(Geldmanetal., 2013; Pouzolsetal., 2014)。对物种的栖息地选择和空间分布研究可了解野生动物种群现状及其栖息地特征(Drouillyetal., 2018), 是生物多样性保护的重要内容, 对确定物种保护的优先级和保护规划具有重要价值(Wintleetal., 2005; Hirzeletal., 2006)。同时, 有助于了解不同物种在空间上的共存模式(Morris, 2003)。
过去野生动物的生态调查, 主要依据动物痕迹、野外直接观察和铗日法等传统方法(唐继荣等, 2001; 刘少英等, 2005), 对警觉、行踪隐蔽、夜间活动、种群数量稀少的物种和珍稀濒危物种而言, 通常难以记录, 得到全面真实的监测结果颇有难度(Gardenetal., 2007; O′connelletal., 2011)。Mackenzie等(2002)开发的占域模型(Site occupancy modelling)可作为一种资源选择函数, 基于出现与不出现调查地点随机样本的探测概率(p), 以估计其各抽样地点的占用率(ψ), 并可引入解释变量利用Logistic回归确定其资源选择, 很好地解决了在抽样地点调查过程中物种缺失的问题(Mackenzieetal., 2006)。同时, 随着红外相机的普及使用, 物种信息可被真实记录, 且因其干扰小、多点位、连续工作等优势, 在不干扰动物正常行为的前提下, 还极大地增加了动物的捕获率, 十分适合应用于占域模型分析之中(O′connelletal., 2011)。其中, 两者结合常应用于动物的行为生态学调查, 确定物种的栖息地选择及其空间分布状况(Penjoretal., 2018), 也可利用多个物种的栖息地占用变化了解同域生存物种的空间关系(Lietal., 2010), 还可延伸到动物群落与所处环境的生态关系(Richetal., 2016)。
贵州梵净山国家级自然保护区(以下简称梵净山保护区)作为国际人与生物圈保护区(MAB)网络成员, 也是长江上游森林生态区生物多样性保护优先区中具有最高保护优先性的区域之一(吴波等, 2006)。独特的自然生态系统, 为多种雉类提供了优良的栖息地, 目前已记录到7种, 物种丰富度高(张明明等, 2019), 但对其分布特征和环境需求了解较少。为了解梵净山保护区雉类的栖息地选择及其空间分布, 本研究基于红外相机的抽样监测方法, 使用占域模型对梵净山保护区所有雉类进行占用率建模, 以了解各物种的栖息地选择和空间分布, 为保护区雉类的种群现状评估和管理提供科学数据。
1 研究地概况
梵净山保护区地处云贵高原向湘西丘陵过渡的斜坡地带, 是武陵山脉的主峰, 地理位置为27°49′50″—28°1′30″N, 108°45′55″—108°48′30″E, 总面积419.00 km2, 海拔500~2 570 m, 较大的高差构造了复杂的高山峡谷地形。同时, 常年受东南亚海洋季风和明显的气候垂直带谱的影响, 形成了中亚热带湿润区山地森林生态系统(周政贤, 1990), 植被类型呈现垂直地带性特点, 并交错分布, 主要植被类型有: 1) 中山常绿、落叶阔叶混交林, 分布于海拔800~1 800 m范围, 主要群落类型有光叶水青冈(Faguslucida)林、贵州青冈(Cyclobalanopsisargyrotricha)林等; 2) 低山常绿阔叶林, 主要分布于海拔1 400 m以下范围, 主要群落类型有栲树(Castanopsisfargesii)林、米槠(Castanopsiscarlesii)林等; 3) 低山针阔混交林, 受人为干扰较重, 有明显次生特点, 多镶嵌于海拔1 300 m以下的低山常绿阔叶林中, 主要群落类型有杉木(Cunninghamialanceolata)林、马尾松(Pinusmassoniana)林等; 4) 低山落叶阔叶林, 分布于海拔1 000~1 800 m范围, 受人类活动影响, 在原有地带性植被消失后而出现次生的植物群落, 主要群落类型包括枫杨(Pterocaryastenoptera)林、赤杨叶(Alniphyllumfortunei)林和枫香(Liquidambarformosana)林等; 5) 高山灌丛和针叶林, 分布于海拔1 800 m以上, 主要群落类型包括铁杉(Tsugachinensis)林、黄山松(Pinustaiwanensis)林、高山柏(Sabinasquamata)灌丛、大花黄杨(Buxushenryi)灌丛等(周政贤, 1990; 杨海龙等, 2010)。保护区内的年平均气温变化在5~25 ℃, 年降水量变化在1 100 ~2 600 mm。
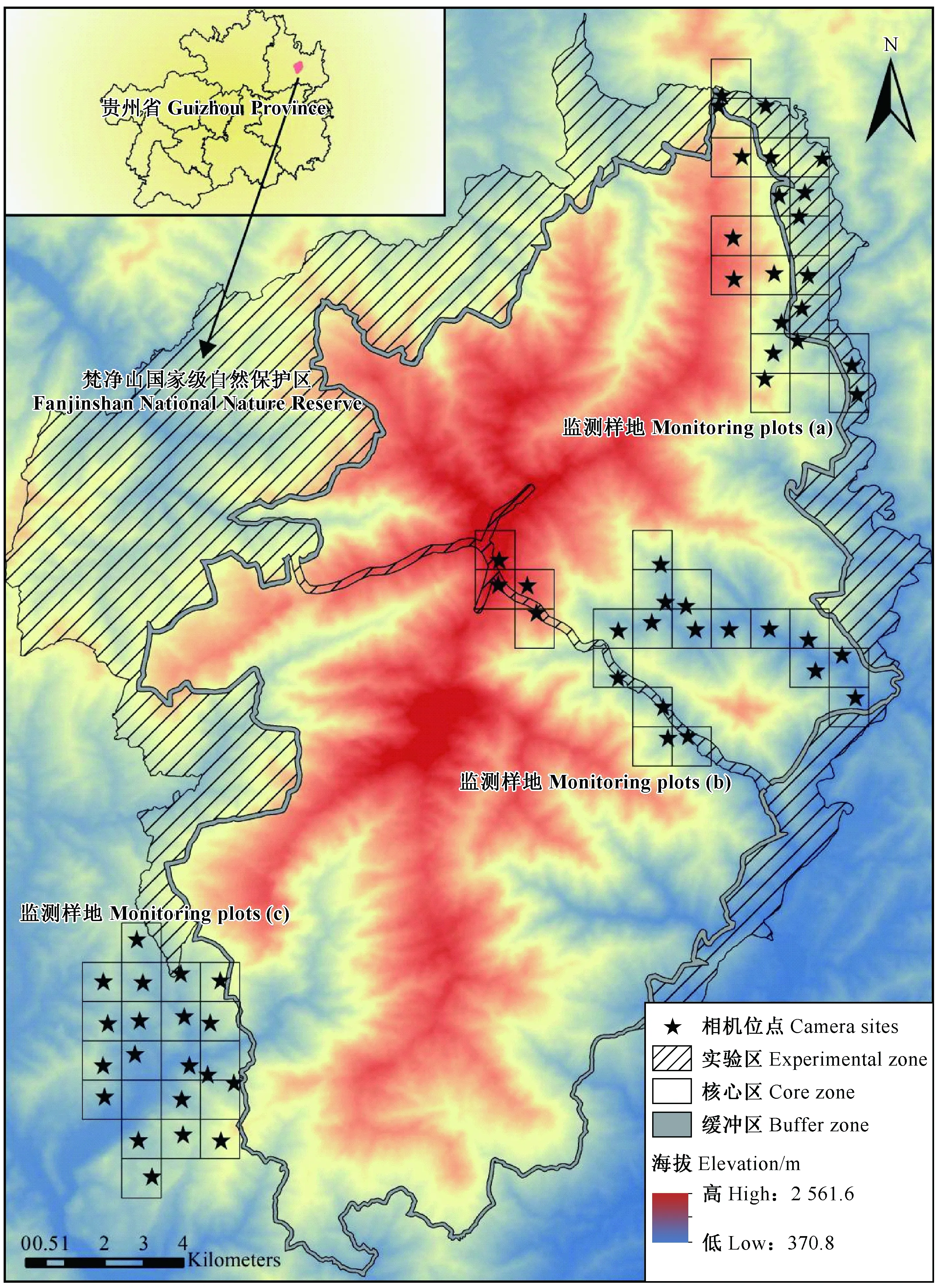
图1 梵净山国家级自然保护区红外相机布设方案Fig.1 The infrared camera locations in Fanjingshan National Nature Reserve
2 研究方法
2.1 相机布设方案
基于梵净山保护区的矢量化边界, 利用ArcGIS 10.2平台, 将梵净山保护区及其周边地区划分为1 km×1 km的网格, 结合梵净山保护区主要植被类型的分布特点(杨海龙等, 2010), 选择3个样区(a、b、c)(图1)。其中, a样区存在连片中山常绿、落叶阔叶混交林, b样区存在连片低山常绿阔叶林, c样区以次生的低山落叶阔叶林和针阔混交林为主。样区内以每千米网格布设1台相机, 每个样区布设20台红外相机, 整个样区覆盖了梵净山保护区的4种主要植被类型(表1)。
相机理论位点为ArcGIS10.2提取的抽样网格中心点坐标, 实际安装过程考虑到理论点的可达性和生境适宜性, 相机实际安装在理论点附近有野生动物活动痕迹的林间开阔地, 并记录位点周边的生境信息。红外相机型号为猎科LTL-6210MC, 拍摄模式设置为拍照, 连拍3张, 时间间隔10 s, 灵敏度为中; 相机固定在距离地面0.5~1.0 m的树干上, 镜头与地面保持平行、避开阳光直射。连续监测期间, 每4个月更换一次, 期间共收集照片7次。
2.2 占域模型
根据研究目的, 本研究选择在雉类的繁殖期内(4—7月), 使用单季单物种占域模型(Single-season occupancy model), 以了解梵净山主要雉类的繁殖期栖息地选择和空间分布关系。基于此, 本研究的占用率应解释为栖息地选择概率, 而不是样点或样方真实的存在比例(Mackenzieetal., 2006)。
为了拟合模型, 使模型收敛,在Presence软件(www.usgs.gov/software/presence)对数据进行测试, 最终拟合以15天为一个采样周期, 期间共采样8次。根据梵净山保护区的主要植被类型及其他生态影响因素, 考虑了6个影响雉类占用率的解释变量(表1)。其中, 植被类型作为相互独立的分类变量, 其不能有效地解释与雉类占用率的线性关系, 但可对不同植被类型的选择偏好提供判别方向。
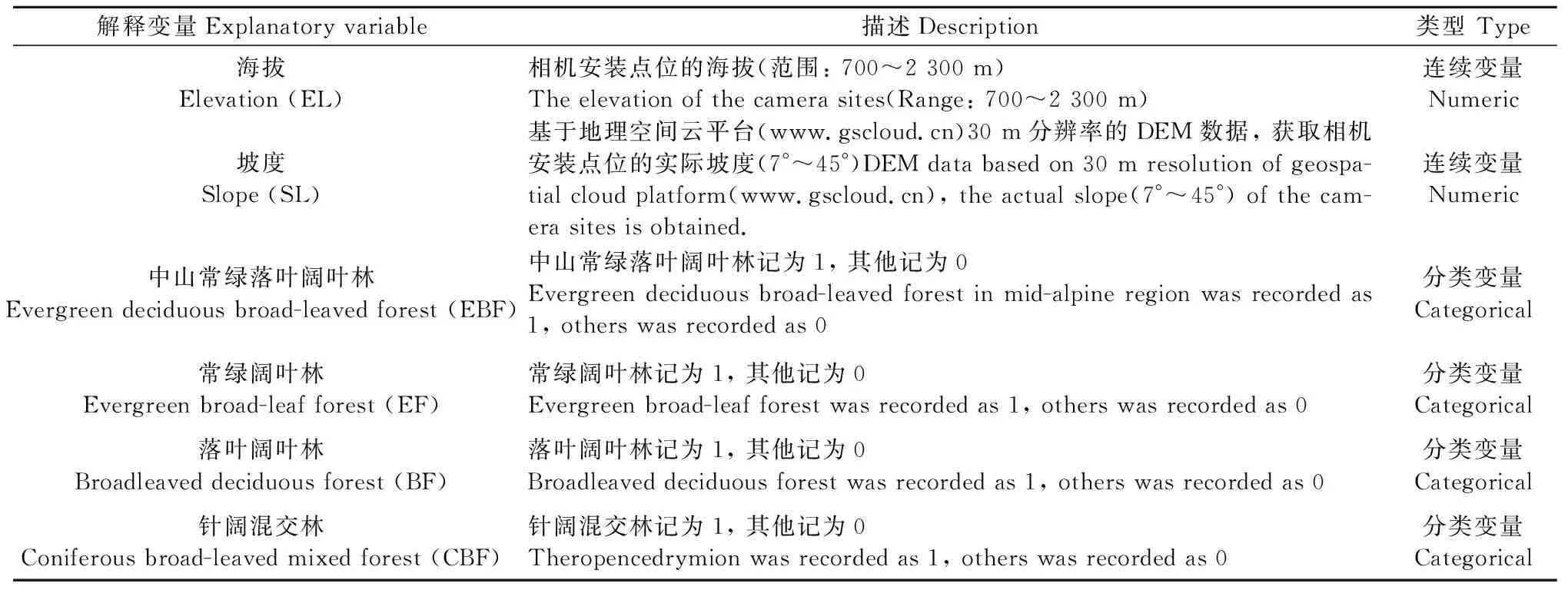
表1 雉类占用率的潜在影响因素①Tab.1 Potential factors influencing the occupancy of pheasant
将采样矩阵与解释变量导入Presence软件, 每个雉类输出64个模型, 所有模型权重合计为1, psi()为占用率,p()探测概率。为评价模型优劣, 所有模型根据Akaike信息准则(AIC)进行排序, ΔAIC≤2是等价最优模型, 将其保留, 合计权重较高的解释变量对雉类的栖息选择具有重要影响(Burnhametal., 2002; Barbierietal., 2004)。其中, AIC值最低的为顶级模型。
另外, 研究考虑了物种间空间共存关系, 基于单季单物种占域模型的探测数据和采样期间的共现数据, 计算2个物种独自的占用率以及当存在到另一个物种时占用率的变化, 以获得物种互作因子(Species interaction factor, SIF):
SIF=ψAB/(ψA×ψB)。
式中,ψAB为2物种共同存在时, 2物种的共有占用率,ψA为物种B不存在时, A物种的占用率;ψB为物种A不存在时, B物种的占用率。SIF=1表明物种间的空间分布是相互独立; SIF > 1表明2物种存在同一地点的概率更高, 空间分布上存在重叠竞争; SIF < 1表明2物种存在同一地点的概率更低, 空间分布上呈现空间回避(Mackenzieetal., 2006)。
3 结果
在2017—2019年的雉类繁殖期的4—7月, 累计连续监测32 972个相机工作日, 共计回收有效位点58个,获雉类可识别照片2 876张, 独立有效照片882张。 记录到7种雉类, 为白颈长尾雉(Syrmaticusellioti)、灰胸竹鸡(Bambusicolathoracicus)、红腹角雉(Tragopantemminckii)、红腹锦鸡(Chrysolophuspictus)、勺鸡(Pucrasiamacrolopha)、环颈雉(Phasianuscolchicus)、白冠长尾雉(Syrmaticusreevesii)(表2)。其中, 白颈长尾雉在31个位点被探测到, 占有效监测网格的53.45%; 灰胸竹鸡在16个位点被探测到, 占有效监测网格的27.59%; 红腹角雉在25个点位被探测到, 占有效监测网格的43.10%; 红腹锦鸡在28个位点被探测到, 占有效监测网格的48.28%; 勺鸡在4个位点被探测到, 占有效监测网格的6.90%, 环颈雉和白冠长尾雉仅在1个点位被探测到, 占有效监测网格的1.72%。因此, 本研究删除了位点和探测次数极少的3种雉类, 以达模型可运行检测范围, 并计算了4种雉类占域模型结果(表3)。

表2 红外相机监测到的梵净山保护区雉类①Tab.2 Pheasant recorded by camera trapping in Fanjingshan National Nature Reserve
模型表明, 海拔是影响4种雉类的栖息地选择的主要因素(表3), 合计权重最高, 并呈现不同海拔分布格局(图2a)。灰胸竹鸡、白颈长尾雉、红腹锦鸡的占用率与海拔呈负相关。其中, 灰胸竹鸡栖息海拔最低,主要栖息于海拔1 100 m以下; 白颈长尾雉主要栖息于1 500 m以下; 红腹锦鸡栖息海拔适应较宽, 高海拔区域也有活动。另外, 红腹角雉的占用率与海拔呈正相关, 占用率随海拔上升而增高。
4种雉类的栖息地选择在植被类型和坡度上表现出相异的模式(图2b、3)。灰胸竹鸡和红腹锦鸡,占用率与坡度呈负相关, 倾向于选择具有次生特点明显, 人为干扰相对较大的针阔混交林栖息,并在空间上存在竞争重叠(表4); 白颈长尾雉和红腹角雉, 占用率与坡度呈正相关; 倾向于选择具有原生性质的常绿落叶阔叶混交林栖息, 并在空间呈现回避特征。此外, 前一组(灰胸竹鸡、红腹锦鸡)与白颈长尾雉空间上存在竞争重叠, 而与红腹角雉呈现空间回避。其中, 灰胸竹鸡表现尤为明显, 未探测到与红腹角雉共现(表4)。
4 讨论
在山地景观中, 海拔变化对野生动植物分布具有重要影响(Torreetal., 2009; Matsuuraetal., 2013), 常沿海拔梯度形成多种植被类型, 导致野生动物的食物资源及其可利用的栖息地类型的多样化, 直接影响到鸟类的栖息地选择(Macfadenetal., 2002)。在梵净山保护区, 海拔同样对4种雉类的栖息地选择具重要影响, 呈现不同的海拔分布差异以及对不同海拔带的适应性(图2a)。
物种共存主要取决于对资源的竞争分化, 使其获得足够的资源以保障生存和繁殖(Schoener, 1974)。其中, 栖息地分化是同域分布近缘物种生态位分化最为普遍和重要的形式, 能够降低物种对空间和其他环境资源的种间竞争, 从而促进近缘物种的相互共存(Parra, 2006)。在梵净山, 红腹角雉的栖息选择与史海涛等(1999)、王德良等(2009)研究结果相似, 呈现适应高海拔的特征, 更喜欢栖息于坡度较陡的原生性阔叶林, 可有效地减少与适应低海拔栖息的另外3种雉类的空间竞争。虽然, 红腹角雉与白颈长尾雉表现相似的植被、坡度选择, 但两者主要海拔栖息段明显不同(图2a), 呈现空间回避状态(表4), 可有效避免竞争。而对于海拔选择相似的雉类而言, 合理的分配栖息海拔段和选择相异生境资源尤为重要(Sachotetal., 2003; 刘鹏等, 2012)。由于长期的人为活动干扰影响, 梵净山的低海拔地段交错分布着多种植被类型(杨海龙等, 2010), 丰富的食物资源, 多样的隐蔽条件, 满足了多种雉类的栖息条件。植被类型选择上, 白颈长尾雉与邻近的贵州雷公山种群表现相似(丁平等, 1996), 更喜欢栖息于原生阔叶林中, 这有别于红腹锦鸡和灰胸竹鸡(图3), 从而减少了对资源的竞争。对于存在空间重叠度高的红腹锦鸡和灰胸竹鸡而言, 红腹锦鸡有较宽的海拔适应性, 同样缓解了资源的争夺, 以维护种群正常生存繁衍。另外, 相关微环境选择研究表明,红腹锦鸡更倾向于乔木层发达的林间开阔地栖息(梁伟等, 2003; 李宏群等, 2011), 而灰胸竹鸡体型小、喜隐伏, 更倾向于灌草密度较高的林下栖息(刘鹏等, 2012)。
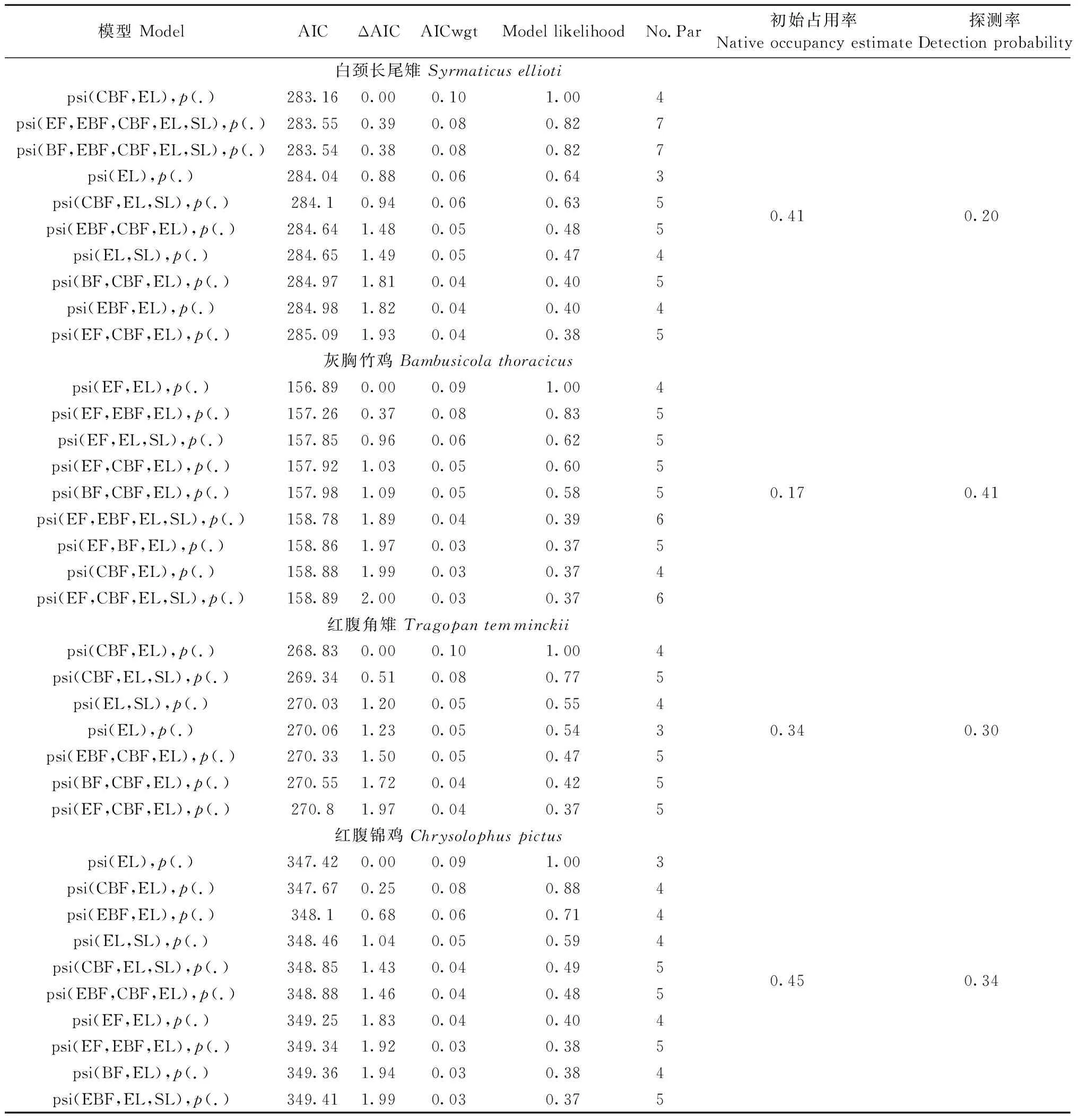
表3 梵净山保护区雉类的最优模型①Tab.3 Optimal model of pheasant in Fanjingshan National Nature Reserve
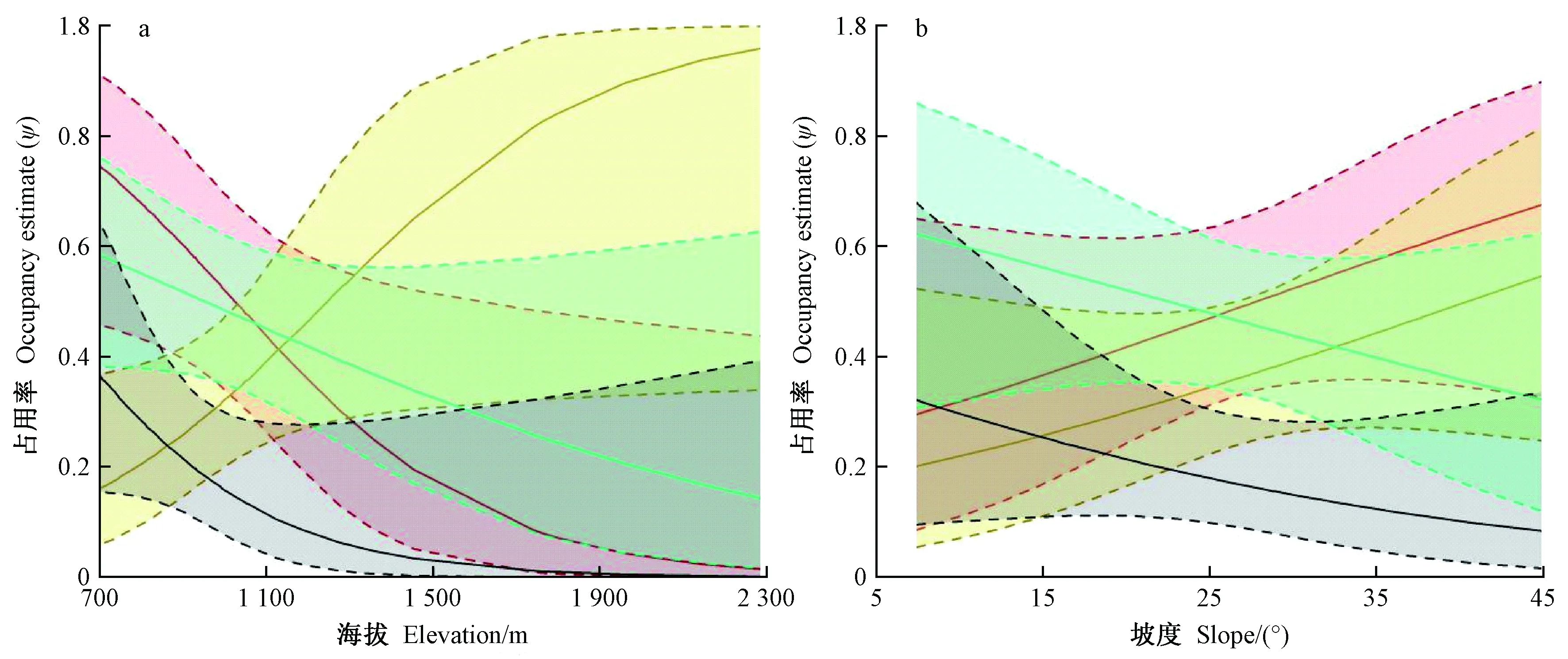
图2 4种雉类的栖息地选择与海拔(a)、坡度(b)的关系Fig.2 Correlations between four species of pheasant estimated habitat use probability and covariates of elevation(a), slope(b)多边形区域为95%置信区间(红线: 白颈长尾雉; 黄线: 红腹角雉; 蓝线: 红腹锦鸡; 黑线: 灰胸竹鸡)。Polygons represent 95% CI(Redlines: Syrmaticus ellioti; yellow lines: Tragopan temminckii; blue lines:Chrysolophus pictus; black lines:Bambusicola thoracicus).
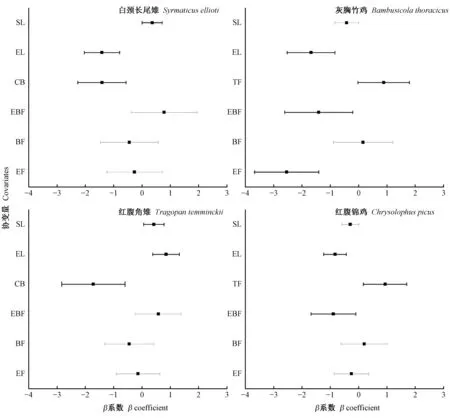
图3 4种雉类ΔAIC ≤ 2的占域模型的变量β系数Fig.3 Variable β coefficient of occupancy model with ΔAIC < 2 for four species of pheasantSE没越过0表示相关显著(黑色误差线),SE越过0表示相关不显著(灰色误差线)。If SE do not overlap zero,it means significant effect (black line),if do overlop zero,it means no significant effect (grey line).
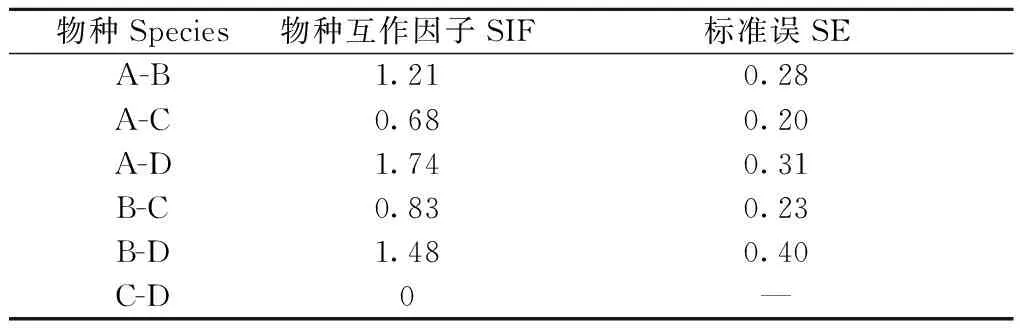
表4 4种雉类的物种互作因子①Tab.4 Species interaction factor between four species of pheasant
雉类在维持生态系统的稳定方面发挥着重要的作用, 其生存状况在一定程度上可作为反映当地森林类型、质量和保护状况的有效指标(张正旺等, 2003)。本次红外相机共监测到7种雉类, 其中具有重要保护价值的白颈长尾雉、红腹角雉和红腹锦鸡在梵净山较为常见(表2), 在保护区周边也频繁活动。Herremans(1998)认为, 在保护区周边增设特定的管理区, 可为多种鸟类提供更大适宜栖息地面积, 增加缓冲屏障, 提高保护效果。同时, 对减少人类活动与野生动物的冲突十分重要, 尤其是活动范围较大的中大型兽类(Keeping, 2014; Weydeetal., 2018)。因此, 保护区管理局应加强保护区周边野生动物重要栖息地的评估, 加大对周边社区的宣传教育, 普及法律法规, 降低盗猎危害, 更有效地保护好重要的濒危野生动物。
占域模型基于调查物种的出现概率可很好的获得物种在一个抽样点的存在比例(Mackenzieetal., 2006), 但对于其所获比例极少或不常见的物种而言, 如本研究中的勺鸡、环颈雉和白冠长尾雉, 仍然不能有效拟合其真实分布情况, 全面了解梵净山保护区7种雉类的种间空间分化模式仍有困难, 但通过2年的连续监测可以确定, 这3种雉类在研究区出现频次较少。另一方面, 雉类栖息地选择通常存在季节性变化, 并表现出动态的空间分布及其种间关系(韩婉诗等, 2017), 本研究了解到梵净山保护区4种主要雉类繁殖期的栖息地选择与空间分布, 但还有必要进一步增加红外相机覆盖度, 摸清梵净山雉类全年的主要栖息地利用特征。
5 结论
本研究利用红外相机监测数据和占域模型, 探讨了梵净山保护区4种主要雉类在繁殖期的栖息地选择差异及其同域共存模式。结果表明, 植被类型的相异选择模式可有效减少4种雉类的空间重叠, 同时4种雉类不同的海拔分布格局, 进一步分离了植被类型选择相似的竞争物种, 增加了物种所在栖息地的活动自由度, 使其获得足够的资源以保障生存和繁殖。另外, 研究结果有助于深入了解这4种雉类的分布情况、栖息地选择和种间关系, 为有针对性地制定保护策略和管理措施提供科学数据。
猜你喜欢
计算机工程与设计(2022年3期)2022-03-22
企业文化(2020年8期)2020-06-03
魅力中国(2019年6期)2019-07-21
乡村地理(2018年3期)2018-11-06
汉语世界(The World of Chinese)(2018年3期)2018-10-22
铜仁学院学报(2018年7期)2018-09-08
贵茶(2018年6期)2018-05-30
厦门航空(2018年4期)2018-04-25
乡村地理(2017年4期)2017-09-18
安阳工学院学报(2015年2期)2015-09-26
