
沾化冬枣生长季碳、氮、磷化学计量特征研究
2021-01-04 07:41:00刘京涛宋爱云赵西梅
中国土壤与肥料 2020年5期
刘京涛,宋爱云,赵西梅,彭 玲,周 峰
(1.滨州学院/山东省黄河三角洲生态环境重点实验室,山东 滨州 256600;2.南京晓庄学院,江苏 南京 211171)
碳(C)、氮(N)、磷(P)是植物生长必需的大量元素,是碳水化合物、蛋白质、磷酸合成等代谢过程的物质基础。植物体内的C、N、P 组成和分配及其与外界环境因子的关系共同决定着植物的营养水平和生长发育过程[1]。生态化学计量学则是研究生态过程中多重化学元素(特别是C、N、P)的平衡和能量平衡,揭示有机体对养分的利用和适应策略[2-3]。
营养元素的可利用性及供应量显著影响有机体生长、种群结构、物种相互作用[4]。因此,判断供应量不足的营养元素类型对调控有机体生长、调节产量、维持生态系统稳定等至关重要[3,5-7]。通过C∶N∶P 化学计量比值的变化则可以判断限制有机体生长、发育或繁殖的营养元素类型[8-11]。
在农业生产上,施肥是实现作物稳产、增产的重要措施。然而,施肥显著改变土壤营养元素的化学计量特征,影响作物对营养元素的吸收机制,促使作物调整化学计量特征,改变生物量分配,进而影响作物产量[12-13]。通过作物和土壤的C、N、P化学计量特征可以确定限制作物生长的营养元素,采取合理的施肥措施,对稳定作物产量,保持作物生态系统平衡有着重要的意义[14]。
冬枣由山东省沾化县开发,在山东省滨州、东营、德州和河北省黄骅、沧州等地广泛种植[15]。沾化冬枣(Zizyphus jujubaMill var.inermisDongzao)已列入中华人民共和国国家标准,受到原产地地域产品的保护[16]。沾化冬枣具有抗盐碱、耐瘠薄的特点,是山东栽种的重要经济林树种之一,但不合理的施肥可造成冬枣品质下降[17]。
国内外已有较多关于经济林木叶片或林下土壤化学计量学特征的研究[18-24],而对冬枣叶片或土壤化学计量学特征的研究相对较少。前期对冬枣的研究主要涉及内源激素[25]、根际微生物[26]、生物肥[27]、土壤养分状况[28-29]及冬枣对养分的吸收分配[30-31]等方面,初步开展了冬枣不同器官生态化学计量特征[32-33]、叶片矿质元素动态变化[34]、枣芽微量元素[35]等方面研究。然而在冬枣叶片及土壤C、N、P生态化学计量特征的时间动态,以及土壤养分条件与冬枣生长状况之间的关联性方面仍需要深入研究。本文以沾化冬枣为研究对象,研究冬枣叶片及土壤C、N、P 化学计量特征及C、N、P 之间的关联性,判定不同生长阶段冬枣生长的限制性营养元素,为冬枣合理施肥,提高产量及改善品质提供理论指导。
1 材料与方法
1.1 研究区概况
本研究所涉及沾化冬枣园位于山东省滨州市沾化区(北纬37°44′54.29″、东经117°55′18.08″),海拔高度4 m,面积为3 300 m2(长66 m,宽50 m)。滨州市沾化区地处黄河三角洲腹地,属北温带大陆性季风气候;年均日照时数为2 627.3 h,年均生理辐射总量2.65×105J/cm2;年均气温12.5℃;年均降水量为544.3 mm,降水多集中于夏季,生长季平均降水量488.2 mm[16]。
1.2 样品采集
研究样地内为均匀种植的沾化冬枣,树龄为14年,行间距为4 m,株间距为2.5 m。采用“五点取样法”设置5 个采样点。每一个样点内选取生长良好、长势一致的枣树2 株,在树冠东、西、南、北4 个方向分别采集生长良好叶片约50 g,同一样株4 个方向的叶片混合均匀为1 个样品。每个样点内,挖80 cm 深土壤剖面1 个,采集0 ~20、20 ~40 、40 ~60、60 ~80 cm 土壤,每层采集土壤约1 kg。于2016 年5 ~10 月,每月中旬每棵样株采集叶片样品约200 g,土壤样品每2 个月采集1 次,每个样品约1 kg。叶片样品于实验室内用去离子水清洗,避免叶面施肥的影响,105℃杀青30 min,85℃烘干至恒重;土壤样品于阴凉通风处自然晾干。所有样品干燥后,研磨粉碎过0.2 mm 筛,储存于自封袋待测。
1.3 样品测定
所有叶片、土壤样品测定C、N、P 含量。其中,采用重铬酸钾外加热法测定土壤样品有机碳(SOC)含量,元素分析仪(Vario EL Ⅲ,德国)测定所有样品全氮(TN)含量,采用钼锑抗比色法测定所有样品全磷(TP)含量。
1.4 数据分析
利用SPSS 13.0 对数据进行统计分析,Origin 8.5 进行制图。
2 结果与分析
2.1 沾化冬枣叶片C、N、P 含量生长季动态
生长季内,沾化冬枣叶片C 总体呈逐渐上升趋势(图1A)。叶片C 含量6 月份最低,为(443.01±1.53)mg/g;10 月份含量达(477.89±1.0)mg/g,叶片C 含量变异系数0.7%~1.4%。沾化冬枣叶片N、P 含量5 月份最高,分别为(42.93±0.38)mg/g 和(3.37±0.16)mg/g,然后显著下降,6 月份后不同月份间差异不显著(图1B、C)。叶片N 含量6 ~10月份变异系数为3.9%~9.8%,小于P 的变异系数(4.9%~21.8%)。
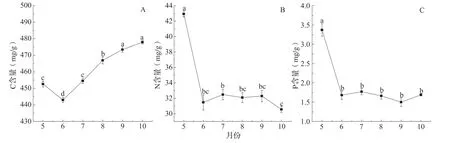
图1 冬枣叶片C、N、P 含量生长季动态
叶片C 含量与N、P 含量之间呈负相关,但其决定系数较小且相关性不显著,说明叶片C 含量与N、P 含量没有显著关联性(图2A、B);N、P 含量呈显著正相关,决定系数达到0.566,说明两者之间密切的关联性(图2C)。
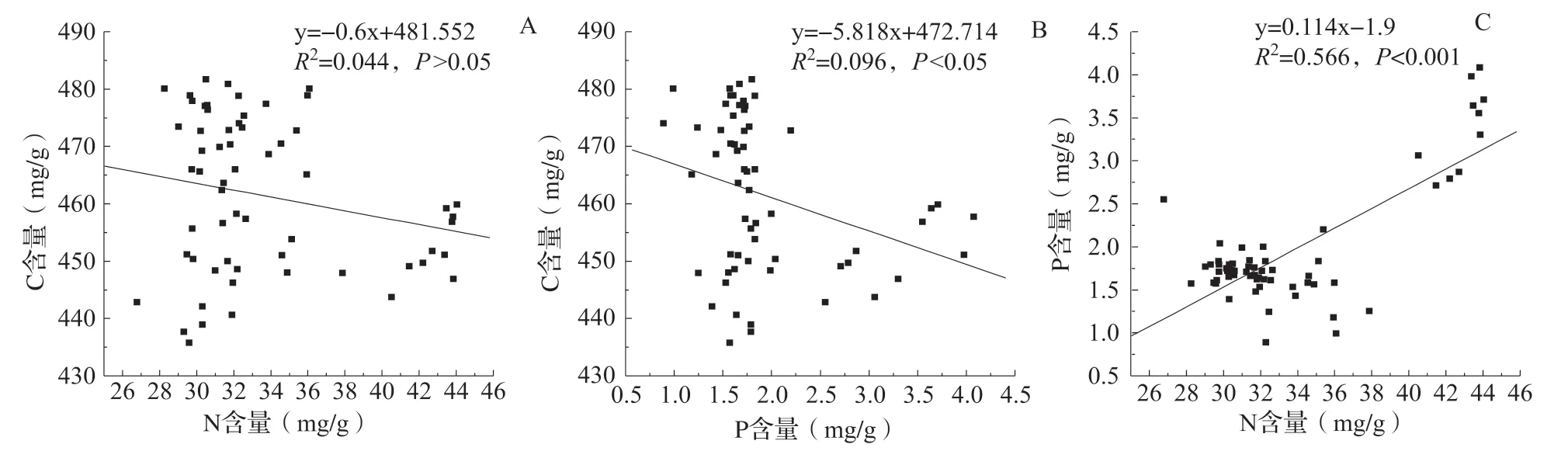
图2 冬枣叶片C、N、P 含量相关性
2.2 林下土壤C、N、P 化学计量特征
冬枣林下土壤C、N、P 含量都随深度增加而降低(表1)。0 ~20 cm 土壤C、N、P 含量分别为22.27 ~35.97、0.94 ~2.6、1.18 ~2.67 mg/g,均值分别为(29.19±1.09)mg/g、(1.73±0.11)mg/g、(1.88±0.1)mg/g,显著高于其他土壤层;20 cm 以下土壤层C、N、P 含量虽然逐渐降低,但各层之间多数差异不显著;同一土层的C、N、P 含量不同月份之间没有显著差异。
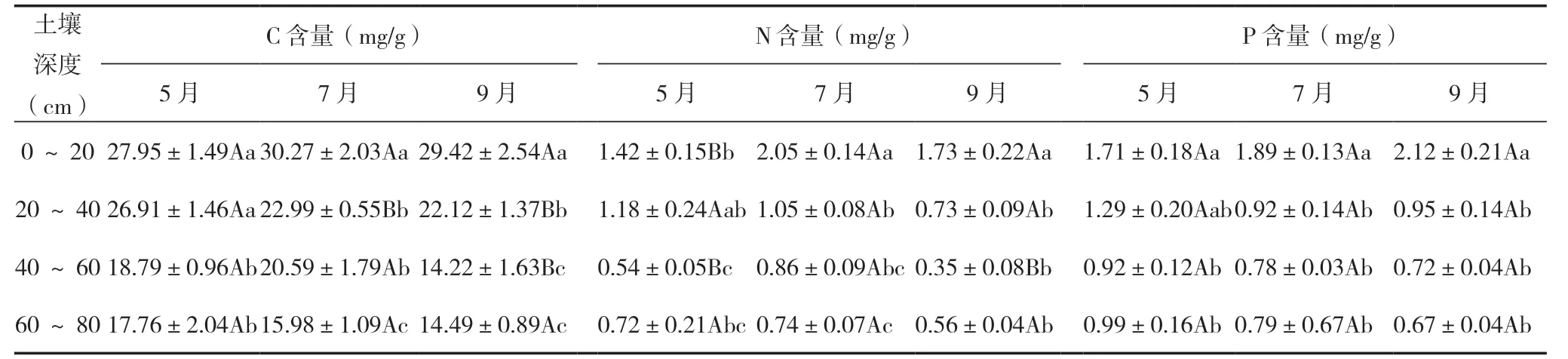
表1 冬枣林下土壤C、N、P 化学计量特征
土壤C∶P、N∶P 质量比随土层加深呈现先增加后下降的趋势,20 ~40 cm 土壤C∶P、N∶P 质量比大于其他土层,而不同月份之间则没有显著差异(表2)。土壤C、N、P 含量之间表现了显著的正相关关系(图3),但冬枣叶片C、N、P 含量与土壤各层C、N、P 含量之间相关性不显著(表3)。
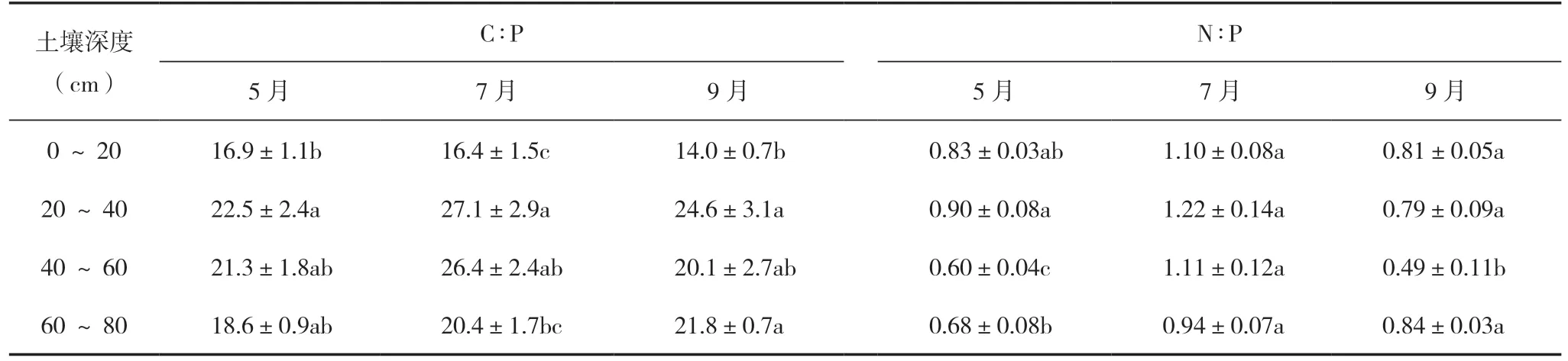
表2 冬枣林下土壤C∶P、N∶P 特征
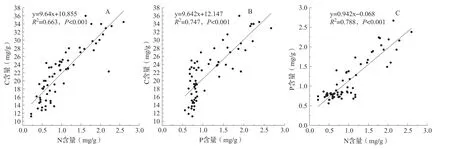
图3 冬枣林下土壤C、N、P 含量相关性

表3 沾化冬枣叶片与不同深度土壤C、N、P含量皮尔森相关性
2.3 沾化冬枣叶片C、N、P 化学计量特征
冬枣叶片的C∶P、N∶P 质量比都呈现增加后趋于稳定的变化趋势(图4)。生长季初期的5 月份最低,叶片C∶P、N∶P 质量比分别为136.9±6.2、12.7±0.5。6 月 份 以 后,C∶P、N∶P质量比显著上升后趋于稳定,各月份之间差异不显著。
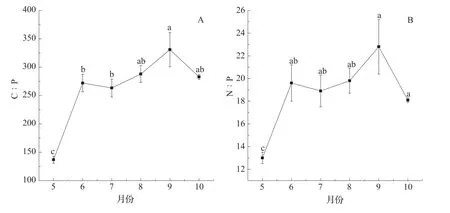
图4 冬枣叶片C、N、P 化学计量比
冬枣叶片的C∶P、N∶P 质量比与叶片C、N、P 含量存在显著相关性,但与不同养分之间的相关程度及相关性质不同(图5)。C∶P质量比与C 含量呈正相关,但决定系数较小,而与P 含量呈显著负相关(图5A、B)。叶片的N∶P 质量比与N 含量呈负相关,但决定系数仅为0.141,与P 含量则呈显著负相关(图C、D)。
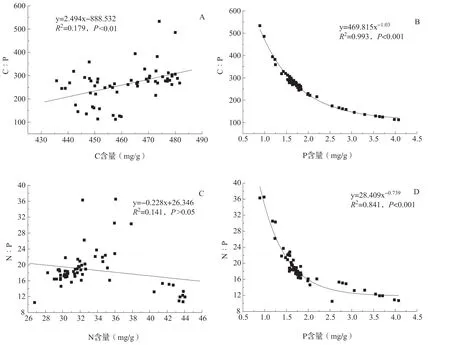
图5 冬枣叶片C∶P、N∶P 与C、N、P 含量相关性
3 讨论
3.1 沾化冬枣叶片C、N、P 含量及其化学计量特征
植物叶片C、N、P 含量与自身结构和生长节律密切相关[36],本研究表明沾化冬枣叶片C、N、P 含量随生长季呈现规律性变化。叶片C 含量随着生长逐渐累积,这与有些学者研究的规律一致[37-38]。沾化冬枣叶片的C 含量10 月份达到最大值(477.89±1.0)mg/g,大于全球陆地植物叶片C含量[(464±32.1)mg/g][39]和全国森林叶片C 含量(455.1 mg/g)[40],与中国东部南北样带森林生态系统102 个优势种叶片C 含量的480.1 mg/g 接近[41]。C 元素在植物体叶片内主要合成有机质,随着植物的不断生长,叶片维管组织不断增加,光合作用的糖类积累,因此冬枣叶片C 含量不断升高。叶片C含量6 月份较低,主要是由于冬枣在这个月份进行树皮环剥,影响了根部吸收的矿质元素的运输,进而影响叶片糖类合成代谢,导致叶片C 含量有所下降。
叶片N、P 含量的变化趋势与植物的生长特征密切相关。植物在生长初期生长缓慢,生物量小,但细胞分裂能力强,需要大量蛋白质和核酸[42],因此,植物叶片N、P 含量相对较高。生长季初期的5 月份,沾化冬枣叶片N、P 含量最高,显著高于其他研究同期冬枣叶片N(23.38 ~33.63 mg/g)含量,但低于P(3.81 ~4.02 mg/g)含量[27,43],显著高于全国陆地植物叶片N(20.2 mg/g)、P(1.46 mg/g)的含量[44]和全球植物叶片N(19.3 ~20.1 mg/g)、P(1.11 ~1.42 mg/g)的 含 量[45-46]。6 月份以后,叶片N、P 含量显著下降,但其他月份叶片的N、P 含量仍高于全国陆地植物及全球植物叶片的N、P 含量。这与有些学者研究的植物叶片N、P 含量在生长季初期显著高于其他阶段的规律一致[38,47-48]。但冬枣叶片N、P 含量在生长旺季逐渐降低并保持一定水平,这与其他研究的规律并不一致。这是由于本研究中的冬枣土壤分别在每年3月和11 月施用腐熟粪肥,7 月和9 月施用叶面肥和复合化肥,导致冬枣叶片N、P 含量在后期没有显著下降。
沾化冬枣叶片5 月份的C∶N∶P 质量比为137∶13∶1,6 ~10 月 份,质量比平均值为287∶20∶1,都显著低于中国东部南北样带森林生态系统102 个优势种叶片的C∶N∶P质量比313.9∶11.5∶1[41]以及全球森林叶片C∶N∶P 质量比469∶13∶1[49]。叶片5 月份C∶N质量比为12.3∶1,6 ~10 月份质量比平均值为17.1∶1,都显著低于中国东部南北样带森林生态系统102 个优势种叶片C∶N 质量比29.1∶1[41]以及全球森林叶片C∶N 质量比37.4∶1[49]。叶片5月份N∶P 质量比为13.0∶1,高于有些学者研究同期冬枣叶片N∶P 质量比8.4∶1[43],低于全国N∶P 质量比16.3∶1[44],接近全球森林叶片N∶P质量比12.6∶1[49];6 ~10 月份,质量比平均值为19.8∶1,高于其他研究冬枣叶片N∶P 质量比8.3∶1 ~15.3∶1[27,50],接近全国N∶P 质量比[44],高于全球森林叶片N∶P 质量比[49]。由于沾化冬枣属于人工经济林,经常进行人工施肥,因此叶片N、P 含量较高,这导致其叶片C、N、P 化学计量比与自然生态系统相比差异显著。
3.2 沾化冬枣林下土壤C、N、P 及其化学计量特征
沾化冬枣林下表层土壤(0 ~20 cm)C、N、P 平均含量分别为(29.19±1.09)mg/g、(1.73±0.11)mg/g、(1.88±0.1)mg/g。N、P 含量高于同区域有些学者研究的冬枣表层土壤(0~20 cm) N (0.63~1.4 mg/g)、P(1.02 ~1.72 mg/g)含量[51];C、P 含量显著高于全国表层土壤(0 ~10 cm)C(24.56 mg/g)、P(0.78 mg/g)含量,N 含量则与全国表层土壤氮含量(1.88 mg/g)接近[52]。土壤剖面(0 ~80 cm)C、N、P平均含量分别为(21.91±0.81)mg/g、(1.01±0.07)mg/g、(1.15±0.07)mg/g,C、P 含量也显著高于全国土壤C(11.12 mg/g)、P(0.65 mg/g)含量总体均值,N 含量则与全国土壤N 含量总体均值(1.06 mg/g)接近[52]。沾化冬枣表层土壤C∶P、N∶P 质量比均值分别为16.0∶1、0.9∶1,土壤剖面C∶P、N∶P 质量比均值分别为20.8∶1、0.9 ∶1,显著低于全国表层土壤C∶P 和N∶P 质量比(52.6∶1、4.2∶1)和土壤总体C∶P 和N∶P质量比(23. 6∶1、2 . 3∶1)[52],但表层土壤N∶P 质量比高于同区域其他研究的冬枣表层土壤N∶P 质量比(0.75∶1)[51]。表层土壤C∶N∶P质量比为15.5∶0.9∶1,显著低于全国土壤表层C∶N∶P 质量比(52∶4∶1);土壤剖面C∶N∶P 质量比为19.1∶0.9 ∶1,与全国土壤总体C∶N∶P 质量比22∶3∶1 接近[52],但显著低于全球土壤C∶N∶P 质量比(72∶6∶1)[53]。
沾化冬枣林下土壤C、N、P 化学计量特征表明土壤C、P 相对丰富而N 缺乏,主要是由于冬枣种植管理过程中,多采用低氮、高磷、高钾的土壤施肥方式所致[54]。土壤中较高的C∶N,将使土壤微生物生长受到N 元素限制,土壤有机物分解减慢,可能导致土壤中C 含量进一步增加。而不同月份之间冬枣土壤养分之间有一定差异,但不同月份的C∶N∶P 相对稳定,说明人为施肥虽然影响沾化冬枣土壤养分,但C∶N∶P 仍存在Redfield比值。
植物叶片养分含量通常与土壤养分含量具有一定相关性[55-56]。自然生态系统中,土壤C 主要取决于土壤的有机质数量及其分解程度,土壤N 主要来源于生物固氮和氮沉降,土壤P 元素则主要是岩石风化。在全球尺度研究表明,土壤TP 可解释48%的植物叶片P 变异[57]。然而,本研究表明植物叶片N、P 含量与土壤TN、TP 之间没有显著相关性。这一方面是因为植物对土壤养分的吸收及其在体内的运输、利用非常复杂,其不仅受到土壤养分含量、水分、温度、pH 值、微生物活性等影响,而且种内、种间竞争等也调控植物对土壤养分的吸收[58-59]。另一方面,沾化冬枣在管理过程中经常进行叶面施肥,养分经过叶片直接进入植物体内而不是来自土壤,也是导致叶片养分含量与土壤养分含量相关性不显著的原因。
3.3 C、N、P 化学计量特征对冬枣施肥的指示意义
“生长速率假说”认为生物体必须改变C∶N∶P 以适应自身生长速率的变化。生物有机体的C∶N∶P 变化主要决定于生物体内P 含量的变化[60]。rRNA 是生物有机体的主要P 库,生长快速的生物有机体需要增加蛋白质合成量以支持快速生长。因此,rRNA 的增加将引起P 浓度上升,C∶P、N∶P 下降[3,7]。本研究中,沾化冬枣叶片P 与C∶P、N∶P 呈显著负相关,而C、N 与C∶P、N∶P 之间相关性不显著,表明冬枣叶片P 对C∶N∶P 具有主导作用(图5)。生长季初期的5 月份,冬枣叶片迅速生长,因此叶片内P含量最高,叶片N∶P 最低,这与“生长速率假说”相吻合。6 月份以后,叶片P 含量下降并保持在一定水平,其N∶P 变化与“生长速率假说”不吻合。本研究的冬枣林下土壤P 含量显著高于全国土壤P含量平均水平,加上叶面施肥及土壤施肥,导致沾化冬枣处于富P 的环境条件,P 不再是限制性营养元素。环境中某一营养元素过剩时,植物存在过量或奢侈吸收现象[61],而P 为非限制性因素时,N∶P与生长速率之间的负相关关系就不一定存在[62-63]。这是导致沾化冬枣叶片N∶P 与“生长速率假说”不一致的原因。
N、P 是植物生长所需的大量营养元素,对植物的生长、发育有着重要的作用,植物叶片N∶P常被用来诊断植物个体、群落以及生态系统N、P的养分限制性[2,9-10]。对湿地植物的研究表明,N∶P>16 时,植物生长受P 限制;N∶P<14 时,受N 限制;14<N∶P<16 时,受到N 或P 一种元素限制,或两者共同限制[2]。而针对陆生植物的施肥试验表明,N∶P>20 时,植物生长受P 限制;N∶P<10时,受N 限制[1]。5 ~10 月份,沾化冬枣叶片N∶P 分 别 为12.7、18.7、18.4、19.4、21.4、18.1。综合前期研究结果的N∶P 临界值,沾化冬枣在5月份生长受到N 限制;在6 ~10 月份,P 的供给量略显不足。本研究中,沾化冬枣土壤N 含量略低于全国平均水平,而N∶P 表明在5 月份N 元素可能限制冬枣生长,因此在生长季初期应该适当增施氮肥。沾化冬枣土壤TP 含量虽然高于全国平均水平,但由于土壤中P 大部分为迟效性状态,土壤TP 作为土壤磷素供应水平指标具有一定局限性[60]。通过测定表明,本研究中的冬枣林下表层土壤有效磷(AP)含量仅为(31.01±3.51)mg/kg,20 ~80 cm 土层AP 含量为(5.37±1.29)mg/kg,显著偏低。本研究中,冬枣林下土壤pH 值为8.61±0.03,而pH>7.5 时,由于大量钙(镁)离子的存在,土壤中的P 元素容易形成难溶性的磷酸钙盐,降低了P 的有效性[64]。6 ~10 月份,冬枣相继进入开花期、坐果期、果实膨大期、果实成熟期,对P 元素的需求量较大,因此P 元素将有可能成为限制性元素。此阶段应该结合氮肥,施用过磷酸钙,增加P元素的供应量和肥效,以提高冬枣产量。
植物生长不仅受到N、P 等营养元素的影响,光照、水分、温度等非生物因素以及生物间相互作用也都影响植物生长。而N∶P 在一定程度上能反映植物N、P 状态,但植物N∶P 受到温度、水分、土壤养分、植物生长阶段等影响[65],不同生活型、不同物种对养分需求的临界值也有差异[3,44]。因此,N∶P 不能作为植物养分限制判定的唯一标准,但植物较低N∶P 时,生长受到N 限制;较高N∶P 时,生长受到P 元素限制,是许多学者普遍认可的理论[1]。结合土壤养分、植物生长阶段等相关因素,在综合分析植物养分状况基础上,N∶P 对沾化冬枣施肥管理仍具有一定的指导意义。
4 结论
生长季内,沾化冬枣叶片C、N、P 含量总体平均值分别为(463.15±1.57)mg/g、(31.78±0.68)mg/g 和(1.66±0.08)mg/g。沾化冬枣叶片C 含量总体呈逐渐上升趋势,10 月份达到最大值;叶片N、P 含量随生育期呈逐渐下降后趋于稳定的趋势,N、P 在5 月份含量最高,6 月份后显著下降且逐渐趋于稳定。冬枣林下土壤C、N、P 在0 ~40 cm的耕作层含量最高,且C∶N∶P 在不同月份之间相对稳定,存在Redfield 比值。冬枣叶片C∶P、N∶P质量比5 月份最低,6 月份以后显著上升后趋于稳定,各月份之间差异不显著,6 ~10 月份平均值为287.4±8.4、19.8±0.7。综合沾化冬枣叶片及土壤C、N、P 化学计量特征,沾化冬枣生长在5 月份受到N 元素限制,在6 ~10 月份受到P 元素限制。
猜你喜欢
落叶果树(2021年6期)2021-02-12 01:28:54
文苑(2020年12期)2020-04-13 00:54:10
农产品市场周刊(2020年23期)2020-01-25 16:17:09
中国新闻周刊(2019年46期)2019-12-20 08:42:17
今日农业(2019年12期)2019-08-13 00:50:16
电子制作(2019年10期)2019-06-17 11:45:22
海峡姐妹(2019年1期)2019-03-23 02:42:46
营销界(2015年25期)2015-08-21 07:24:52
书香两岸(2014年4期)2014-05-30 08:11:12
书香两岸(2014年4期)2014-05-30 08:11:12
