
病原真菌对琥珀酸脱氢酶抑制剂抗性研究进展
2021-01-04 08:09胡伟群
世界农药 2020年12期
胡伟群
(浙江省化工研究院,杭州 310000)
农作物病虫害是影响农业持续和健康发展的重要制约因素。农药作为控制农林作物病虫草鼠等有害生物危害的特殊商品,在保护农业生产、提高农业综合生产能力、促进粮油安全等方面发挥重要作用。近年来,SDHI类杀菌剂因结构新颖、活性高以及杀菌谱广等优势,受到各大农药公司的高度关注,她们投入大量精力和财力开发该类药剂。目前,有23个SDHI类杀菌剂品种已经上市,此类产品已成为市场销售额增长最为迅速的一类药剂,在防治各种重要植物病害中发挥重要作用。2016 年,SDHI类杀菌剂的全球销售额为 16.91亿美元,占全球杀菌剂市场销售额的11.%[1]。但是,随着SDHI类杀菌剂的大量使用,多种病原真菌已经对该类药剂产生了抗性,且抗性频率不断发展,这在一定程度上影响了该类药剂的防治效果。本文首先介绍 SDHI类药剂的作用机制,在此基础上,重点综述病菌对该类药剂抗性发生和发展情况,阐述抗性机制,进而讨论抗性治理策略。
1 SDHI类杀菌剂的作用机制
生物通过呼吸链进行呼吸产生生物能。通常情况下,真菌体内一个分子葡萄糖完全氧化为CO2和H2O时,在细胞内可产生36个ATP分子,其中32个是在呼吸链中通过氧化磷酸化形成的。因此,呼吸链是研发杀菌剂的重要靶标,抑制或干扰呼吸链的杀菌剂常常有很高的杀菌活性。
在真菌线粒体呼吸链上有 6个关键酶复合物(I到 VI)参与电子传递,并通过电子传递产生 ATP。在复合物 I中,电子从还原态烟酰胺腺嘌呤二核苷酸(NADH)传递到辅酶Q,该过程由NADH-Q还原酶催化。在复合物Ⅱ中,琥珀酸脱氢酶(SDH)催化琥珀酸形成延胡索酸以及从泛醌辅酶Q还原到还原型辅酶Q的偶联反应。然后,在辅酶Q细胞色素c氧化还原酶的催化下,辅酶Q或还原型辅酶Q将电子传递到细胞色素 bc1酶复合物(复合物III)。最后,细胞色素c将电子经过细胞色素a和a3氧化酶(复合物Ⅳ)传递到最终的受体O2。在特殊环境下,在真菌体内的电子能够绕过正常的呼吸路径,从辅酶Q传递到O2,这一途径对氰化物不敏感,由旁路氧化酶(the alternative oxidase,也称复合物V)催化,这种呼吸作用也称为旁路呼吸(alternative respiration)。在呼吸电子传递过程中,所释放的质子在几个不同的位点由ATP合成酶(复合物VI)催化,经氧化磷酸化产生ATP。
琥铂酸脱氢酶由黄素蛋白(SdhA)、铁硫蛋白(sdhB)和另外2种嵌膜蛋白(SdhC和SdhD) 4个亚单位组成。SdhA和SdhB组成该复合体的可溶性部分,具有琥珀酸脱氢酶活性;嵌膜蛋白SdhC和SdhD将SdhA和SdhB固定在内膜上,且具有泛醌还原酶活性[2]。SDHI类杀菌剂作用于琥铂酸脱氢酶,通过完全或者部分占据底物泛醌的位点,抑制了电子从琥珀酸到泛醌的传递,干扰呼吸链上复合体II电子传递,阻止其产生能量,进而抑制病原菌生长,最终导致病菌死亡[3]。药剂与药靶精细互作试验表明,SDHI的酸性部分插入琥铂酸脱氢酶的泛醌结合位点(Q位点),与SdhC的R46和S42以及SdhB的I218和P169位氨基酸通过范德华(vdW)力相互作用;而药剂的胺基延伸到Q位的开口部,与SdHC的W35、I43以及I30氨基酸通过vdW相互作用;药剂的羰基氧原子与SdhB的W173和SdhD的Y91氨基酸互作;精细的药剂-药靶互作试验有助于设计更加高效的SDHIs新药剂[4-5]。
2 病菌对SDHIs抗性发生情况
尽管 SDHIs对多种病原菌有很高的抑菌活性,但由于药剂作用于琥铂酸脱氢酶这个单一的位点,而且该靶标蛋白存在较高的变异性,因此,国际杀菌剂抗性行动委员会[Fungicides Resistance Action Committee,FRAC(https://www.frac.info/)]将SDHI类杀菌剂归为中等至高抗性风险药剂。从1970年报道了第一个对SDHI类药剂抗性的真菌突变体以来[5],随着SDHI类药剂的广泛应用,目前已经有20多种病菌对该类药剂产生抗性,其中16种病菌在田间已经对它们产生抗性(表1)。抗药水平及抗性频率因不同病菌-药剂组合而异。例如,SDHI类的啶酰菌胺 2005年在我国登记防治多种作物的灰霉病。2012-2013年测定发现,湖北不同地区草莓的灰霉病菌对啶酰菌胺抗性频率在0~4.76%[6-7]。Zheng等人报道从山东葡萄上分离的灰霉病菌对啶酰菌胺产生了严重抗性,抗性菌株比例达 100%[8]。因此,在今后一段时间,随着SDHI类药剂的广泛使用,我国灰霉病菌对该类药剂的抗性问题还会加重。
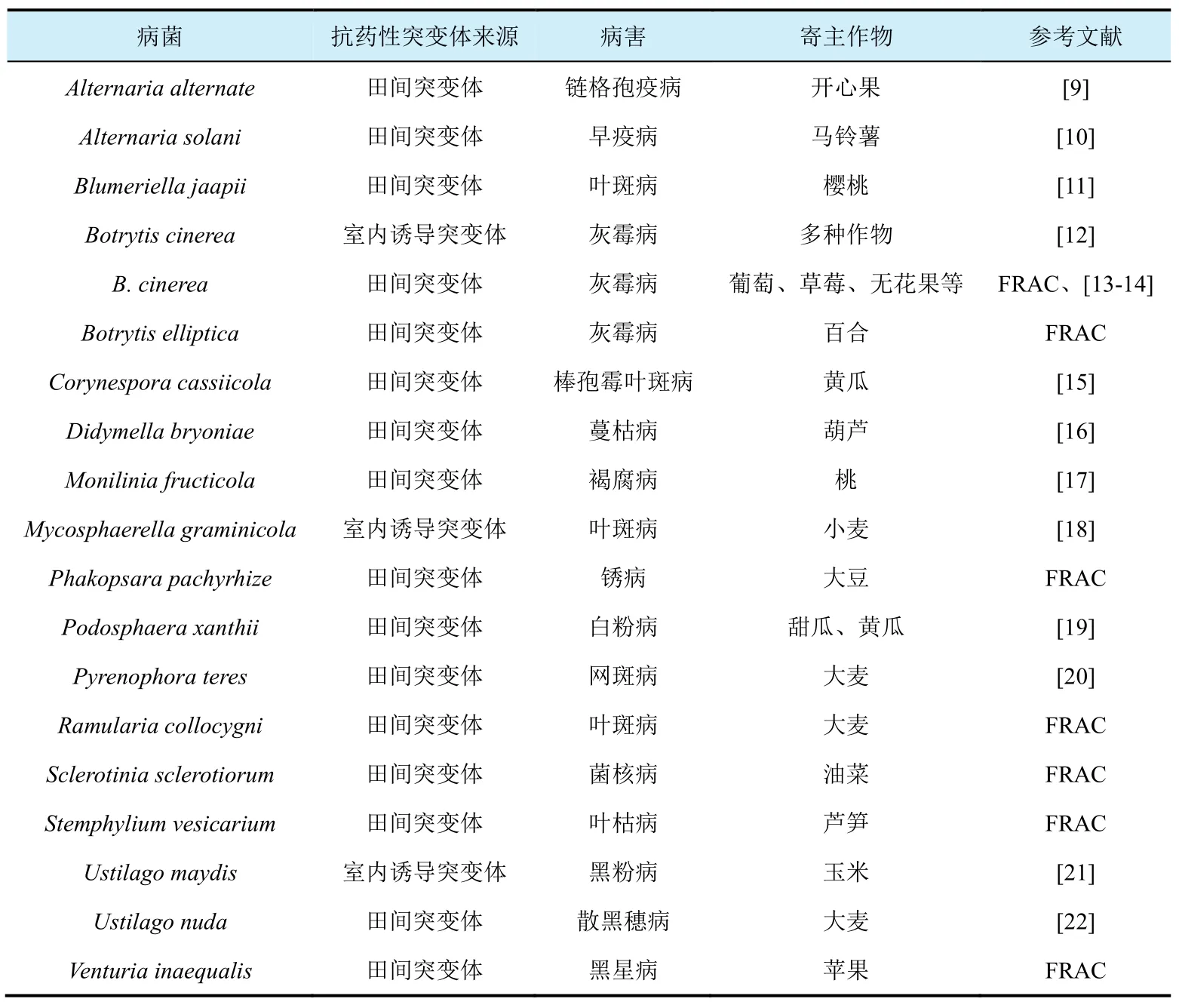
表1 目前已报道的对SDHI类杀菌剂产生抗性的病原真菌
3 病原真菌对SDHIs的抗性机制
研究发现,病原菌对SDHIs的抗性主要由琥铂酸脱氢酶复合体亚基上氨基酸突变所致。琥铂酸脱氢酶复合体亚基上的氨基酸突变,改变了药物与复合体的相互作用,降低药剂与药靶的亲和力,进而引起病菌的抗药性。目前,田间菌株对SDHIs的抗性主要由SdhB亚基突变所致。例如,灰霉病菌SdhB亚基上第 225、230或 272氨基酸位点分别发生P225L/F/T (第225位的脯氨酸突变成亮氨酸、苯丙氨酸或苏氨酸)、N230I (第230位的天冬酰胺突变成异亮氨酸)或 H272L/R/Y(第 272位的组氨酸突变成亮氨酸、精氨酸或酪氨酸)突变,导致灰霉病菌对啶酰菌胺产生不同水平的抗性[23-24]。例如,H272L点突变的灰霉病菌对啶酰菌胺表现高水平抗性,但对其他SDHI类药剂表现低到中等抗性;N230I点突变的灰霉病菌对啶酰菌胺、氟吡菌酰胺和氟唑菌酰胺表现中等抗性,对吡唑萘菌胺、联苯吡菌胺和萎锈灵等仅表现低水平抗性[25]。有趣的是,同一位点突变成不同的氨基酸,可以引起不同的抗性水平和抗性谱。例如,SdhB亚基上H272L的突变,使得灰霉病菌对所有 SDHI类药剂均产生抗药性;但H272Y的突变使得灰霉病菌对啶酰菌胺产生抗性,但对氟吡菌酰胺反而更加敏感[24]。目前,SdhB亚基突变引起多种病菌,包括Alternaria alternata、Alternaria solani、Corynespora cassiicola、Didymellabryoniae、Podosphaera xanthii、Sclerotinia sclerotiorum以及Stemphylium vesicarium等对SDHIs产生抗药性。
除了SdhB亚基发生突变之外,SdhC和SdhD亚基上突变同样引起SDHIs抗性问题。苹果黑星病菌(Venturia inaequalis) SdhC亚基第 151位发生H151R突变,导致病菌产生抗药性。番茄早疫病菌(Alternaria solani) SdhD亚基的第133位发生H133R突变,引起病菌对SDHIs的抗性。目前,尚未有SdhA亚基发生突变引起抗药性的报道。
最新研究还发现,小麦叶枯病菌群体中存在一类对SDHIs天然抗性的菌株,这些菌株在SDHI类药剂使用之前已经在自然界存在。对这类菌株的抗性机制研究发现,这类菌株中除了SdhC基因之外,还有一个SdhC的同源基因(命名为ZTDHC3),该基因同样能够编码琥珀酸脱氢酶 C亚基(命名为alt-SdhC)。反向遗传学试验表明,alt-SdhC与正常的SdhC亚基一样,能够与其他3个琥铂酸脱氢酶亚基结合,形成功能完整的琥珀酸脱氢酶酶,并且alt-SdhC蛋白中独特的Qp位点残基使得这类菌株对SDHIs产生抗性[26]。这些结果表明,自然界中病原真菌对SDHI类药剂存在多种抗药机制。
4 SDHIs抗性治理策略
由于 SDHI类杀菌剂属于中高抗性风险药剂,在使用一段时候后,病菌比较容易对这类药剂产生抗性。因此,在使用药剂过程中,必须要进行抗性治理。抗性治理的基本策略包括⑴ 实施病害综合治理,降低病害压力;⑵ 同一个生长季节,要严格限制 SDHI类药剂的使用次数,且药剂应该在发病初期使用,减轻药剂对病菌群体的选择压力。以葡萄灰霉病为例,FRAC建议一个生长季节,一般情况下SDHI类药剂的使用不超过3次;在一个生长季节使用 12次以上药剂的地区,SDHI类药剂最多只能使用4次;而且SDHI类药剂需要和其他作用机制的药剂轮换使用,避免连续使用。⑶ 由于SDHI类药剂之间存在交互抗性,因此,药剂混用时,必须将不同作用机制的药剂进行混用,不能将 SDHI类药剂进行混用。⑷ 要加强病菌抗药性监测,及时了解抗性发生情况,并针对性地制定实施病害防控方案。⑸ 强化药剂抗性机制及构效关系研究,针对抗性突变位点,设计和研发反抗性或难以产生抗药性的新药剂,从而克服抗性问题。
猜你喜欢
中国药学药品知识仓库(2022年10期)2022-05-29
麦类作物学报(2022年3期)2022-05-19
临床与实验病理学杂志(2022年3期)2022-04-06
家庭科学·新健康(2021年5期)2021-06-21
西北农业学报(2021年4期)2021-05-19
智慧健康(2021年33期)2021-03-16
岭南急诊医学杂志(2020年6期)2021-01-14
中华养生保健(2020年4期)2020-11-16
科学与财富(2020年1期)2020-03-02
烟台大学学报(自然科学与工程版)(2020年1期)2020-02-08
