
玉米幼苗响应低铁胁迫的根系形态与干物质积累特征
2020-12-31 05:58:20李哲馨
作物杂志 2020年6期
任 云 刘 静 李哲馨 李 强
(重庆文理学院园林与生命科学学院(特色植物研究院),402160,重庆)
铁(Fe)是植物生长发育所需的关键微量营养元素,可参与一系列重要的生理生化过程[1]。Fe参与叶绿素合成,约80%的Fe集中在植物叶绿体中,Fe除了参与光合作用外,也参与细胞解毒作用,是过氧化氢酶和过氧化物酶活性中心的组成成分[2]。植物能够通过2种方式从根际中获取Fe,即基于螯合和还原的双重机制[3]。尽管Fe在地壳中含量丰富,但由于Fe在氧化环境、中性和碱性土壤条件下趋向形成氧化态和氢氧化态,土壤中可被植物直接利用的离子态Fe含量很低[4],从而导致作物缺Fe黄化,影响产量[5]。
不同植物或同一植物不同品种对Fe胁迫的适应能力存在显著差异[6-7]。Dellÿ Orto等[8]研究发现,大豆在缺Fe时,叶绿素含量明显减少。不同Fe效率大豆品种的划分是相对的,不仅由自身特性所决定,也与胁迫程度紧密相关[9]。Ciaf fi等[10]和Zamboni等[11]指出低Fe胁迫对小麦生长有显著影响,小麦幼苗表现出植物缺Fe的典型症状(如叶绿素降低)以及其他矿物营养的不均衡积累,地上部分和根的生长均减少30%。Jia等[12]发现3种苹果砧木对低Fe胁迫存在明显的抗性差异,且苹果砧木地上部生物量、株高、叶绿素含量、总根长和侧根数与耐低Fe能力呈显著正相关。章爱群等[13]和龙文靖等[14]发现低Fe胁迫下玉米幼苗地上部与根系Fe含量均明显下降,且存在显著的基因型差异,且Fe处理和基因型之间存在显著互作效应。
1 材料和方法
1.1 试验材料
供试材料为西南地区主要玉米杂交种32个,品种编号及名称见表1。
表1 供试玉米品种编号及名称Table 1 Number and name of maize varieties in test
1.2 培养方法
于2017年10月在重庆市永川区重庆文理学院温室大棚内进行水培试验,采用两因素完全随机设计。用清水漂洗玉米种子并除去包衣,用70%酒精消毒30s,蒸馏水清洗并浸泡10h,待种子露白转移至粒径为1~2mm的湿润石英砂盒发芽,早晚浇蒸馏水1次,放置于光强12 000lx、昼夜长短及温度14h(26℃)/10h(22℃)、相对湿度60%的人工培养箱内催芽。设置100μmol/L Fe(Ⅲ)-EDTA(正常,NFe)和10μmol/L Fe(Ⅲ)-EDTA(低铁,LFe)2个Fe水平处理,营养液pH值5.8±0.2。幼苗2叶1心时除去胚乳后移至改良Hoagland营养液中培养,待幼苗长至3叶1心时进行低Fe胁迫处理(LFe营养液),以Hoagland营养液(NFe)作为对照。水培容器为45cm×33cm×20cm的黑色塑料盆,覆2cm厚含20个孔的泡沫板,每孔定植1株玉米苗,用海绵包茎以固定植株。幼苗在14h/10h(28℃/22℃)的光温周期的生长室中培养,每隔3d更换1次营养液,用NaOH调节营养液pH值至5.8,并使用通气泵通气。
1.3 指标测定
所有指标均在处理后7d测定,形态指标、根系性状和生物量的测定均为3次重复,每重复10株幼苗。
1.3.1 形态指标 株高为胚芽鞘节至最长叶叶尖的长度,用直尺测量。茎粗为玉米幼苗基部离根1cm处的最大直径和最小直径,用游标卡尺测量,取平均值。可见叶数为包括心叶在内的所有可见叶片数。展叶数为叶枕已伸出下一叶叶鞘的叶片数。测量每片绿叶的最大叶长和最大叶宽。若在一片叶中有部分是黄色的,则同样测量绿叶部分的最大叶长、最大叶宽,采用长宽系数法计算绿叶面积。
1.3.2 根系性状 用根系扫描仪(Expression 10000XL,Epson)及图像分析软件(WinRhizo Pro 2009)测定分析根表面积、根体积、总根长、根平均直径和根数。
1.3.3 生物量 将幼苗分为地上部分和根系,于120℃杀青30min,80℃烘至恒重,称量干重。分别计算根冠比和单株干重,根冠比=根系干重/地上部干重,单株干重=地上部干重+根系干重。
1.3.4 Fe含量 采用原子吸收光谱法测定幼苗地上部和根系Fe含量[17]。活性Fe含量:叶片鲜样切碎后用1mol/L HCl按1∶10的比例浸提(连续振荡5h),过滤后,用原子吸收分光光度计测定浸提液中Fe的含量。含Fe量:称取1.00g经烘干粉碎的玉米植株样品于塑料瓶中,加入25.0mL Vc-HCl(0.40g VC溶解后,加入166.0mL浓HCl定容至1L),置于80℃恒温振荡器上振荡2h,用蒸馏水补至原重后过滤,用原子吸收分光光度计测定浸提液中Fe的含量。Fe积累量=含Fe量×单株干重;根系Fe吸收效率=Fe积累量/根系干重;Fe生理效率=单株干重/Fe积累量。
60例患者,随机分为中药治疗组和西药治疗组。患者均有3次或3次以上流产史,年龄24~36岁,夫妻双方染色体检查正常,无感染性疾病,无先天性生殖系统畸形。两组患者在年龄构成、观察指标等方面比较,无显著性差异(P>0.05),具有可比性。
变异系数=标准偏差/均值×100%;低Fe效应=(LFe条件下性状值-NFe条件下性状值)/NFe条件下性状值×100%。
1.4 数据整理与统计分析
利用Excel 2010整理测定的数据。利用SPSS 21.0软件对整理好的数据进行统计分析,利用“相关”和“双变量”函数对16个性状进行相关性分析,利用“降维”和“因子分析”函数对16个性状进行主成分分析,利用“比较均值”和“ANOVE”方差分析函数进行差异显著性分析。采用Min-Max方法对各性状统计数据进行标准化处理,利用变换公式:(n-最小值)/(最大值-最小值),使所有性状值在0~1的特定区间,其中n是自变量,然后使用TBtools软件基于最长距离法和欧氏距离法进行层次聚类,并绘制聚类热图[18]。
2 结果与分析
2.1 2个Fe水平下玉米苗期性状及相关性分析
方差分析结果(表2)显示,除了品种与Fe处理的互作对茎粗的影响不显著外,其余性状在品种、Fe处理、品种与Fe处理的互作上均存在极显著差异。进一步分析发现,在NFe条件下,16个性状中以根体积、总根长和根表面积的变异系数较大,分别达到30.46%、27.34%和27.33%,叶绿素含量的变异系数最小,仅有6.34%,其余12个性状的变异范围为7.71%~27.27%。而在LFe条件下,根尖数的变异系数最大,为43.62%,可见叶数的变异系数最小,仅为8.32%,其余14个性状的变异系数范围为11.85%~39.73%;在LFe效应上,仅有根直径和根冠比表现出正效应,其余14个性状均为负效应,其中以根尖数负效应值最大(–42.04%),说明低Fe胁迫对玉米幼苗的生长影响较大,对根尖数的影响最大。
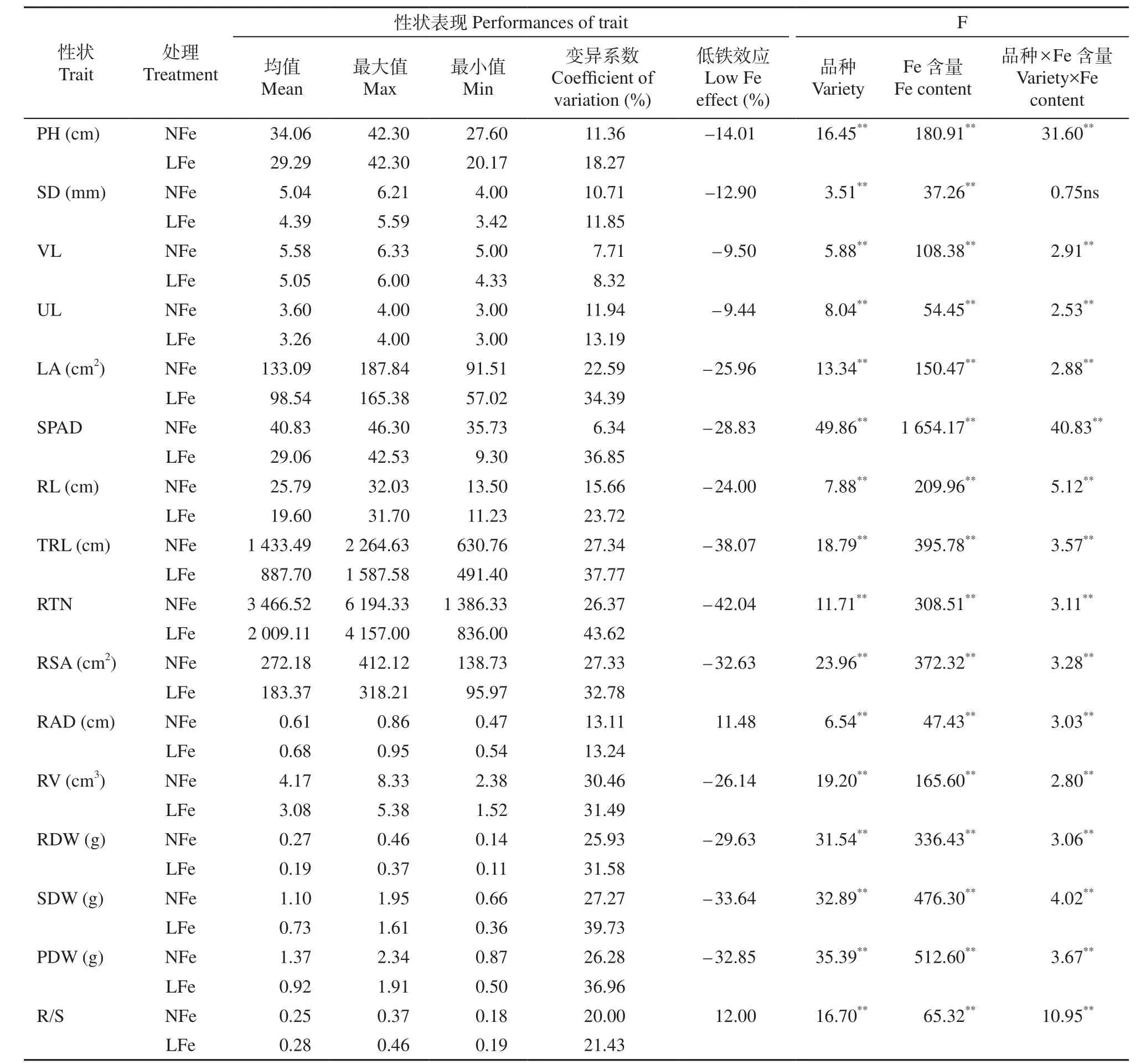
表2 2个Fe水平下不同玉米杂交种性状Table 2 Seedling traits of different maize inbred lines at two Fe application levels
相关性分析结果(表3)表明,在NFe处理下,地上部和单株干重与叶绿素含量、根直径和根冠比相关性不显著,根干重与叶绿素含量、根长和根直径相关性不显著,分别与其余12个性状呈显著或极显著相关,而在LFe处理下,地上部干重、根干重和单株干重除与根直径相关性不显著外,与其余14个性状均极显著相关。在NFe处理下,与株高、茎粗、可见叶、展叶、叶面积、叶绿素含量、根长、总根长、根尖数、根表面积、根直径、根体积、茎叶干重、总干重和根冠比相关性显著或极显著的性状分别有12、13、9、7、11、1、9、11、11、12、3、11、12、12 和 2 个;而在LFe处理下,与上述性状相关性显著或极显著的性状分别有 14、14、14、13、14、12、14、14、13、13、2、13、14、14和 7个( 表 3)。 由此可见,玉米幼苗各性状在2种Fe处理下的变异来源具有明显差异。

表3 2个Fe水平下不同玉米品种苗期16个性状间相关性分析Table 3 Correlative analysis of sixteen traits at seedling stage among different maize varieties under two Fe levels
2.2 2个Fe水平下玉米幼苗性状的主成分分析和聚类分析
主成分分析结果表明,4个主成分的累积贡献率在NFe和LFe处理下分别达到88.53%和84.26%,说明这4个成分在2个Fe水平下均具有较强的代表性(图1)。NFe处理主成分1的方差贡献率为52.40%,在主成分1中,除了根平均直径和根冠比为负向载荷外,其余性状均为正向载荷,且相关系数均大于0.3,主要反映“幼苗长势因子”;主成分2和主成分3分别反映所有变异的15.07%和10.47%,相关性状包括根直径、根冠比、总根长、根尖数和根表面积,可定义为“根系生长因子”;主成分4可以解释全部变异的6.31%,相关性状有可见叶数、展叶数和叶绿素含量,主要体现“叶片生长因子”;在LFe条件下,主成分1的方差贡献率为61.79%,同样除了根直径和根冠比为负向载荷外,其余性状均为正向载荷,且相关系数均大于0.6,主要反映“幼苗长势因子”;主成分2和主成分3分别解释全部变异的11.19%和9.48%,相关性状有总根长、根尖数、根表面积、根直径、根冠比、根体积和根干重,也可定义为“根系生长因子”;主成分4反映了全部变异的6.07%,较大相关性的性状只有可见叶数和展叶数,主要体现“叶片生长因子”。2个Fe水平处理下主成分的主要因子相对稳定,但性状的相关程度存在变化。

图1 2个Fe水平下玉米幼苗性状的主成分分析Fig.1 Principal components analysis of maize seedling traits at two Fe application levels
对32个玉米品种的上述16个性状进行聚类分析,发现在NFe和LFe处理下16个性状均可以聚为7组(图2)。NFe处理下,1组为可见叶数和展叶数(叶片生长因子),2组为根直径和根冠比(根系生长因子),3组为地上部干重和单株干重(幼苗长势因子),4组为根体积、根表面积和根干重(根系生长因子),5组为总根长和根尖数(根系生长因子),6组为株高、茎粗和叶面积(幼苗长势因子),7组为根长和叶绿素含量。LFe条件下,NFe处理的1、2、3和6组的性状仍聚为一组,相对稳定,其余组的性状在2个Fe水平下存在明显的组别变化。综上可以看出,各个性状在2个Fe水平下的不同主成分间和聚类间的主要因子相对稳定,聚类和主成分间的变化可能与其在2个Fe水平下相关性的差异存在一定关系。
2.3 2个Fe水平下不同玉米品种的聚类分析
对32个玉米品种上述16个性状进行聚类分析(图2),发现在NFe处理下,32个玉米品种聚为6组,分别包含6、5、5、5、7和4个品种;而在LFe处理下,聚为7组,分别包含5、5、5、3、5、3和6个品种,玉米品种在2个Fe水平下存在明显的组别变化。
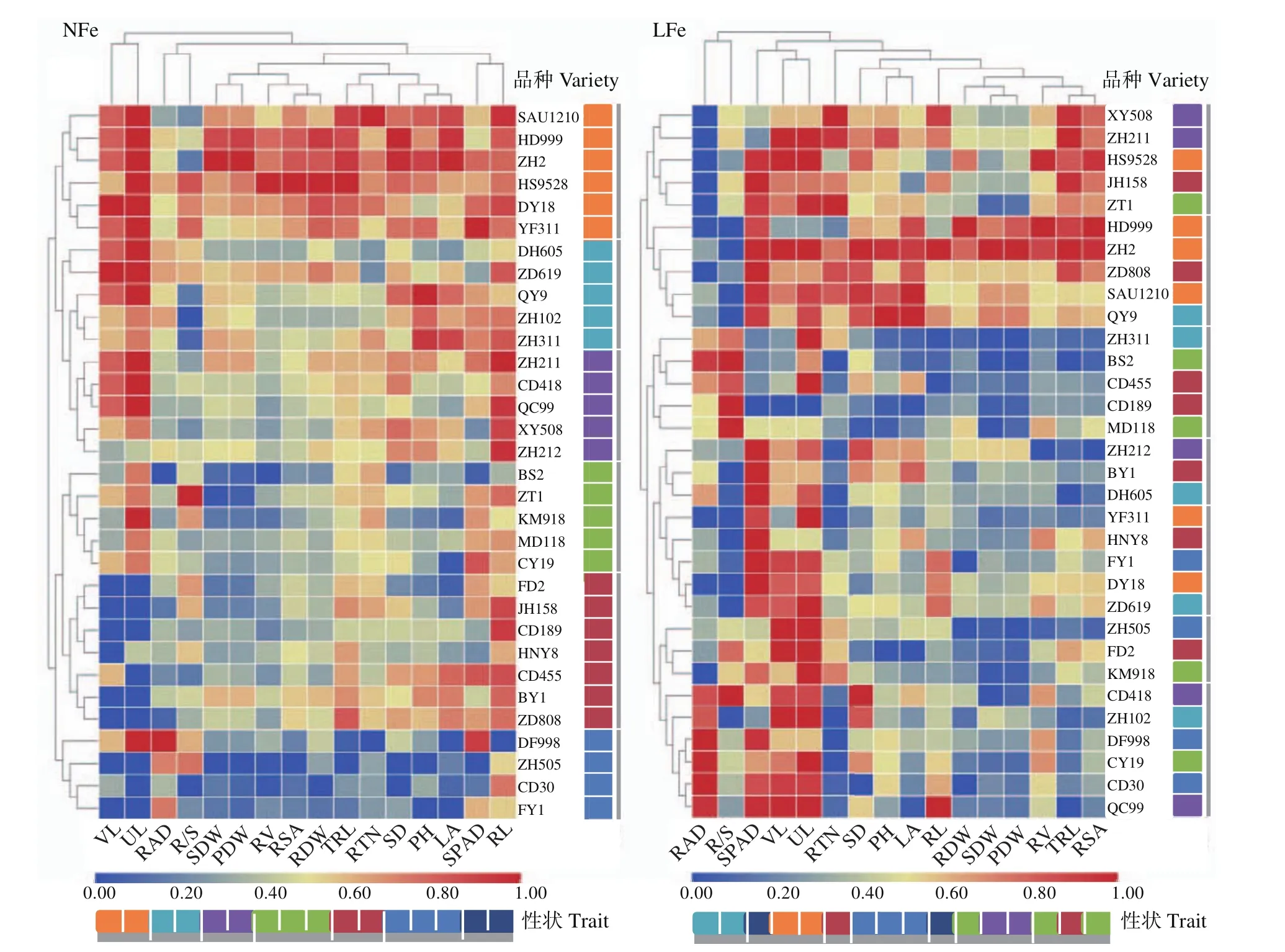
图2 2种Fe素水平下32个玉米品种基于不同性状的聚类分析Fig.2 Clustering analysis of 32 maize varieties based on different traits at two Fe application levels
对在2个Fe水平下聚类结果中各组在不同性状上的差异进行进一步分析,发现在NFe处理下,第1组中的6个品种除根直径和根冠比低于其余品种外,其余14个性状均有更好的表现;第2组中的5个品种根冠比低于其余品种,其余性状表现相对较好;第3组和第4组除可见叶数、展叶数和根长相对较好,其余性状表现相对较差;而第5组和第6组中除展叶和根直径相对较好外,其余性状上均劣于其余组中的品种(图2)。在LFe处理下,第1组和第2组的10个品种的根直径和根冠比相对较差,而其余性状表现一般或较好;第3组的5个品种在根直径和根冠比上表现相对较好,而在其余性状上表现一般或较差;第4、5和6组的11个品种的叶绿素含量、可见叶数和展叶数表现相对较好,而其余性状表现较差;第7组的6个品种除叶绿素含量、可见叶数、展叶数和根直径表现相对较好,其他性状表现均较差(图2)。综上可以看出,不同玉米品种的同一性状对NFe和LFe处理的响应存在明显差异。
2.4 玉米苗期性状对低Fe胁迫的响应差异
进一步对32个玉米品种各性状的低Fe效应值进行聚类分析(图3),结果显示32个玉米品种可以聚为5组,1~5组中分别包含8、7、5、6和6个品种,且每一组各性状对低Fe胁迫的响应存在明显差异(图3A)。在LFe处理下,根直径在5组中均显著增加,而根冠比在第1、2组中显著降低,第4组差异不显著,第3、5组中显著增加(图3B)。据此推测,不同组别的品种对低Fe胁迫的响应可能受品种与Fe处理互作的影响。
低Fe效应值聚类分析(图3)显示,第1组和第2组的品种适应低Fe能力较强,包含15个品种,结合2个Fe水平下基于不同性状的聚类(图2),正红 2号、汉单999和荃玉9号在NFe(第1组和第2组)和LFe(第1组和第2组)条件下性状都表现较好,为Fe高效品种;第3组和第5组对低Fe表现最敏感,包含11个品种,结合2个Fe水平下基于不同性状32个玉米品种的聚类(图2),川单 455、川单 189和成单30在NFe(第5组和第6组)和LFe(第3组和第7组)条件下性状都表现较差,为Fe低效品种。

图3 基于LFe 效应值的32个玉米品种的聚类分析及性状变异Fig.3 Clusters of 32 maize varieties and the variations analysis between different traits based on LFe condition
2.5 不同玉米品种Fe吸收和利用差异
对以上低Fe效应聚类成的5组玉米品种的Fe吸收及利用效率进行分析,结果(表4)表明,低Fe胁迫下,5组玉米品种的Fe积累量均显著降低,其中对低Fe表现敏感的第3组降幅最大,达到72.88%。根冠比相对降幅较小或增加,由此造成5组玉米品种的根系Fe吸收效率在LFe处理下较NFe处理均显著降低,其中也以第3组降幅最大,降低了61.48%。5组32个玉米品种在LFe处理下的Fe生理效率均显著提高,其中适应低Fe能力较强的第1组增加幅度最大,达到160.62%;其余依次为第2组、第4组、第3组和第5组。
对上述Fe高效(正红2号、汉单999与荃玉9号)/Fe低效(川单455、川单189与成单30)品种的Fe吸收及利用效率单独进行分析,结果(表5)显示,在LFe胁迫下,所有Fe高效/Fe低效品种的单株Fe积累量和根系Fe吸收效率较NFe处理下均显著降低而Fe生理效率均显著提高。Fe高效品种中单株Fe积累量、根系Fe吸收效率的降幅与根冠比的增幅均低于Fe低效品种,而Fe生理效率的增幅高于Fe低效品种。
3 讨论
Fe高效玉米品种的评价和选择是缓解玉米低Fe胁迫,保障石灰性土壤玉米高产稳产最简单有效的措施之一。徐健钦等[21]指出,地上部与根系生物量、叶片SPAD值是表征玉米幼苗Fe利用效率的最佳指标,根冠比及叶片活性Fe含量是良好指标。龙文靖等[23]将展叶数、叶面积、根长、根干重、Fe积累量作为玉米苗期Fe高效筛选的重要指标。本研究分析了32个玉米品种各性状的变异系数,发现总根长、根表面积、地上部和单株干重的变异系数在2个Fe水平下均较大,且这些性状

表4 玉米品种物质积累与Fe吸收利用差异Table 4 Differences of matter accumulation and Fe absorption and utilization in maize varieties

表5 不同耐低Fe玉米品种性状Table 5 Traits of different lowiron-tolerant maize varieties
优先分配给根系,利于根系维持生长[21]。低Fe胁迫下玉米的苗干重、根干重和总根长均减少、根体积和根表面积也有不同程度的降低,而根质量比与平均根直径明显提高[22]。本研究表明,低Fe胁迫导致玉米幼苗的总根长减小,根体积、根系干重均显著降低,而根直径和根冠比显著增加(表2),这与前人研究结果基本一致[22]。对32个玉米品种的16个性状进行聚类分析,发现在NFe和LFe处理下主要代表“根系生长因子”的第4和5组(根体积、根表面积、根干重、总根长和根尖数)的性状存在明显的组别差异,表明低Fe胁迫对玉米幼苗的影响主要表现在根系间存在极显著的相关性。进一步分析发现,品种总根长、根表面积、地上部干重和单株干重在Fe高效和Fe低效品种间的差异也十分显著(表5),表明总根长、根表面积、地上部干重和单株干重可以作为玉米Fe效应能力的参考评价指标。
Fe是影响植物生长发育的重要营养元素之一[19]。低Fe胁迫可以诱导地上部同化产物向根系转运,促进根系代偿性伸长[13]。低Fe胁迫会导致玉米幼苗的根长、根体积、根系活力、干物质量、Fe含量、Fe积累量、相对吸Fe能力降低[20]。低Fe胁迫下,玉米幼苗地上部将光合作用产物性状上。
植株对Fe的吸收利用是一个复杂的过程,不同植株在低Fe条件下表现不同[24-25]。徐健钦等[21]研究表明,低Fe胁迫下Fe高效品种Ye478根冠比较Fe低效品种根冠比更高,认为提高根冠比是玉米提高低Fe胁迫适应性的原因。陈旭蕾等[26]发现,低Fe胁迫下,玉米品种的株高、SPAD值、地上部生物量、根部生物量以及总生物量均显著降低,而根冠比显著增加,且根冠比与Fe吸收效率呈显著的负相关。本研究也得出类似的结果,低Fe胁迫下,玉米幼苗单株干重、单株Fe积累量、根系Fe吸收效率和叶片活性Fe含量均显著下降,而根冠比和Fe生理效率显著增加,与前人研究结果一致[14]。但进一步对比发现:Fe高效品种(正红2号、汉单999和荃玉9号)单株干重、单株Fe积累量和Fe吸收效率的降幅与根冠比的增幅均低于Fe低效品种(川单455、川单189和成单30),而Fe生理效率的增幅高于Fe低效品种,因此推测提高Fe生理效率是玉米幼苗适应低Fe胁迫的重要生理机制。这与徐健钦等[21]的研究结果不一致,主要是因为本研究在品种筛选过程中涉及较多根系性状,所筛选出的高效品种为Fe高效吸收型,而Ye478的Fe高效是根向地上部转运效率以及Fe在体内的再分配引起的。
4 结论
低Fe胁迫对玉米幼苗影响较大,根干重、地上部干重、单株干重、总根长、叶绿素含量、叶面积、根表面积、根体积、株高、根尖数、可见叶数、展叶数和茎粗均显著降低,根平均直径和根冠比显著增加,单株Fe积累量和Fe吸收效率显著下降,而Fe生理效率显著增加。32个玉米品种对低Fe胁迫的敏感程度存在较大差异,其中正红2号、川单999与荃玉9号耐低Fe能力相对较强,可以作为玉米Fe高效育种的种质资源。
猜你喜欢
安徽农学通报(2022年6期)2022-04-07 21:30:29
小小说月刊·下半月(2021年8期)2021-09-05 05:49:31
中国科技纵横(2018年14期)2018-08-30 09:15:34
浙江农林大学学报(2018年4期)2018-08-22 01:54:12
动漫界·幼教365(大班)(2018年3期)2018-05-14 11:50:46
畜牧与饲料科学(2018年3期)2018-05-08 02:48:35
现代农业科技(2017年16期)2017-09-22 14:47:46
山东农业科学(2017年4期)2017-05-18 17:06:13
湖南林业科技(2017年1期)2017-02-06 05:28:55
湖南农业科学(2014年22期)2014-09-28 03:23:28
