
茶园周边景观格局对茶小绿叶蝉种群遗传结构的影响
2020-12-11 07:11李金玉牛东升尤士骏尤民生
昆虫学报 2020年10期
李金玉,牛东升,陈 杰,尤士骏,*,尤民生,*
(1.福建农林大学应用生态研究所,闽台作物有害生物生态防控国家重点实验室,福州 350002; 2.福建农林大学,教育部害虫生态防控国际合作联合实验室,福州 350002; 3.福建农林大学,农业部闽台作物有害生物综合治理重点实验室,福州 350002)
由于茶树为多年生常绿木本植物,传统茶园中的生物群落结构通常比其他作物生态系统的群落结构复杂,并且在相当长的时间内可以保持动态平衡(陈李林等,2019)。近年来,随着茶树种植面积的持续增加,茶树栽培制度也发生了变化,越来越多地采用集约化经营和单一化种植的模式,导致茶园及其周围的景观环境也逐渐趋于简单化,容易引发茶园病虫害的流行和扩散(李乐,2012;周宁宁,2014;陈李林等,2019)。与此同时,大量使用化肥和农药致使茶园生态平衡日益遭到破坏,引起有害生物特别是刺吸式害虫的发生危害不断加重。
茶小绿叶蝉Empoascaonukii是茶园中分布最广,为害持续时间最长,造成损失也最大的头号害虫,在我国可造成减产10%~15%,严重时减产50%以上(Chenetal.,2019)。有学者提出茶小绿叶蝉是在20世纪60年代侵入茶园后迅速上升为我国茶园害虫区系中的头号害虫(陈宗懋,1979),并普遍认为这可能与茶园景观环境的简单化过程以及化肥、农药施用水平的变化有关(李乐,2012;周宁宁,2014;Chenetal.,2019)。因此,在周边景观格局分布不同的茶园中对茶小绿叶蝉种群结构差异和扩散特点进行研究有助于理解其发生机制,进而为茶园景观合理布局和开展可持续害虫治理提供重要的信息。
近20年来,得益于分子技术和地理空间分析的快速发展,分子水平上的微观分析手段与景观生态学的宏观统计工具相结合,对于连接景观环境与生物种群结构、适应性和迁移扩散过程提供很大的帮助(Storferetal.,2007)。一些学者利用RAPD、线粒体基因序列和微卫星分子标记对茶小绿叶蝉不同地理种群进行了分析(李乐,2012;Fuetal.,2014;周宁宁,2014;Zhangetal.,2016,2019)。这些研究的重点大多用于解析不同种群的亲缘关系和确认是否属于相同的优势种,较少通过基因流的估算和基因流格局的分析以获得其种群迁移、扩散相关的信息,也没有将这些遗传结构、基因流和扩散信息与茶园景观环境相关联,并用于阐明茶小绿叶蝉在不同茶园景观环境中的种群分布和扩散特点。本研究以福建省安溪县为研究区域,选取了周边景观结构和组成不同的18个茶园采集茶小绿叶蝉,基于23个微卫星分子标记对这些茶小绿叶蝉种群进行了遗传结构和基因流格局的分析,探究在周边景观格局不同的茶园中茶小绿叶蝉的种群结构差异和扩散特点,为阐明其灾变机理、优化茶园景观合理布局方案和害虫可持续治理提供依据。
1 材料与方法
1.1 研究区域选择和样本采集
选择福建省安溪县作为研究区域,县域范围117°36′~118°17′E,24°50′~25°26′N,全县总面积3 057.28 km2(税伟等,2017)。区域内地势从西北向东南倾斜,西北部山峰林立,东南部地势相对平缓,以丘陵山地为主。茶园在全县区域内均有分布,但由于明显的地势地貌差异,不同乡镇的种植模式有明显差异:在位于南部的西坪等乡镇,主要采用单一化和集约化的种植模式,在茶园周围少见自然和半自然生境或其他作物田;在位于西北部的桃舟和福田等乡镇,茶园大都分布于高海拔的山坡上,周围经常会有森林等自然和半自然生境分布;而在位于东部的蓬莱和湖头等乡镇,种植模式则会更多样化,茶园周围经常会有其他多种作物田的分布(税伟等,2017)(表1)。
基于以上信息,在研究区域内选取了周边景观格局不同的18个茶园(其中茶树有多年树龄)进行研究(表1),避开选择新建茶园以避免近期茶苗移植对分析的影响。2018年8月,在选定的每个茶园中,随机选择位于其中央的5~10个点(彼此间隔20~100 m),用扫网法采集至少50头茶小绿叶蝉成虫。本研究将来自同一样点的个体作为一个种群用于后续的研究和分析。
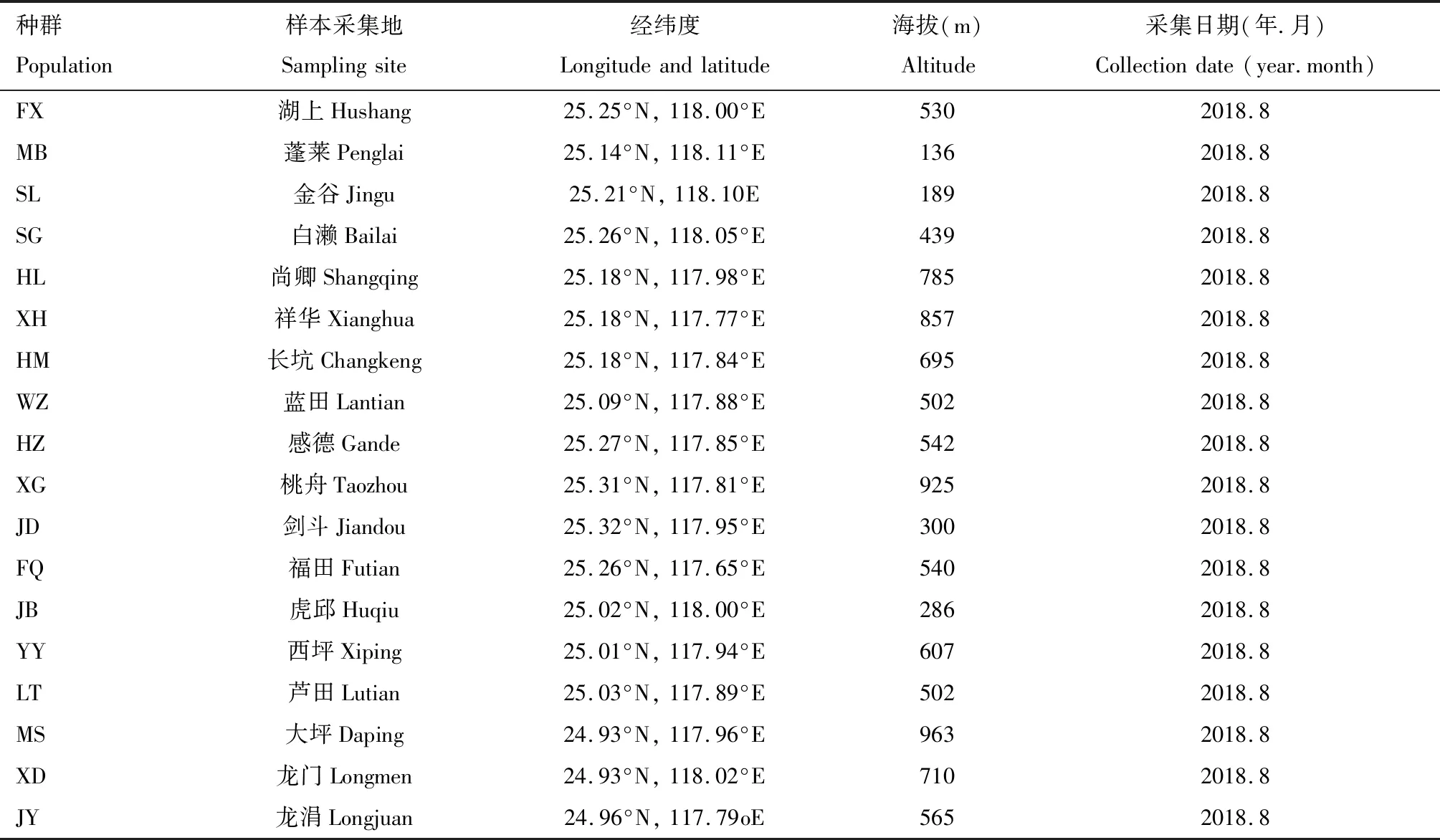
表1 福建省安溪县茶小绿叶蝉采样点信息Table 1 Information of sampling sites of Empoasca onukii in Anxi County,Fujian Province,southeasterm China
1.2 微卫星位点基因型检测
本研究使用天根生化科技(北京)有限公司的微量样品基因组提取试剂盒,对单头茶小绿叶蝉进行基因组DNA的提取。为了保证样品种类的一致性,分析个体在基因组提取之前首先进行初步形态学鉴定(Qinetal.,2015),并参照Demichelis等(2010)的COI基因引物以及PCR反应体系和条件对每份基因组样品进行扩增,将产物序列(经ABI 3730XL测序成功)与NCBI中茶小绿叶蝉Empoascaonukii的线粒体全长序列(Liuetal.,2017)进行比对,仅保留可以比对上的样品用于后续微卫星位点的扩增和分析。
本研究选取23个微卫星位点作为分子标记研究茶小绿叶蝉种群遗传变异,引物序列信息参照Zhang等(2016)和李倩等(2016)(表2)。PCR扩增采用天根生化科技(北京)有限公司的多重PCR扩增试剂盒(KT109),将4~6对引物组合在同一个体系中进行扩增,反应体系和条件参照其使用说明。PCR产物经电泳检测后,利用ABI 3730XL测序仪(Applied Biosystems)结合GENEMAPPER 3.2软件(Applied Biosystems)分析确定其中对应位点的片段长度和识别片段基因型,利用Micro-Checker(Van Oosterhoutetal.,2004)检测是否存在因为无效等位基因和滑移导致的位点读取错误的问题。最终,我们获得了来自安溪县18个样点的530个个体的微卫星基因型数据用于后续种群分析。

表2 本研究中所用的23个微卫星位点的引物信息Table 2 Primer information of 23 microsatellite loci in this study
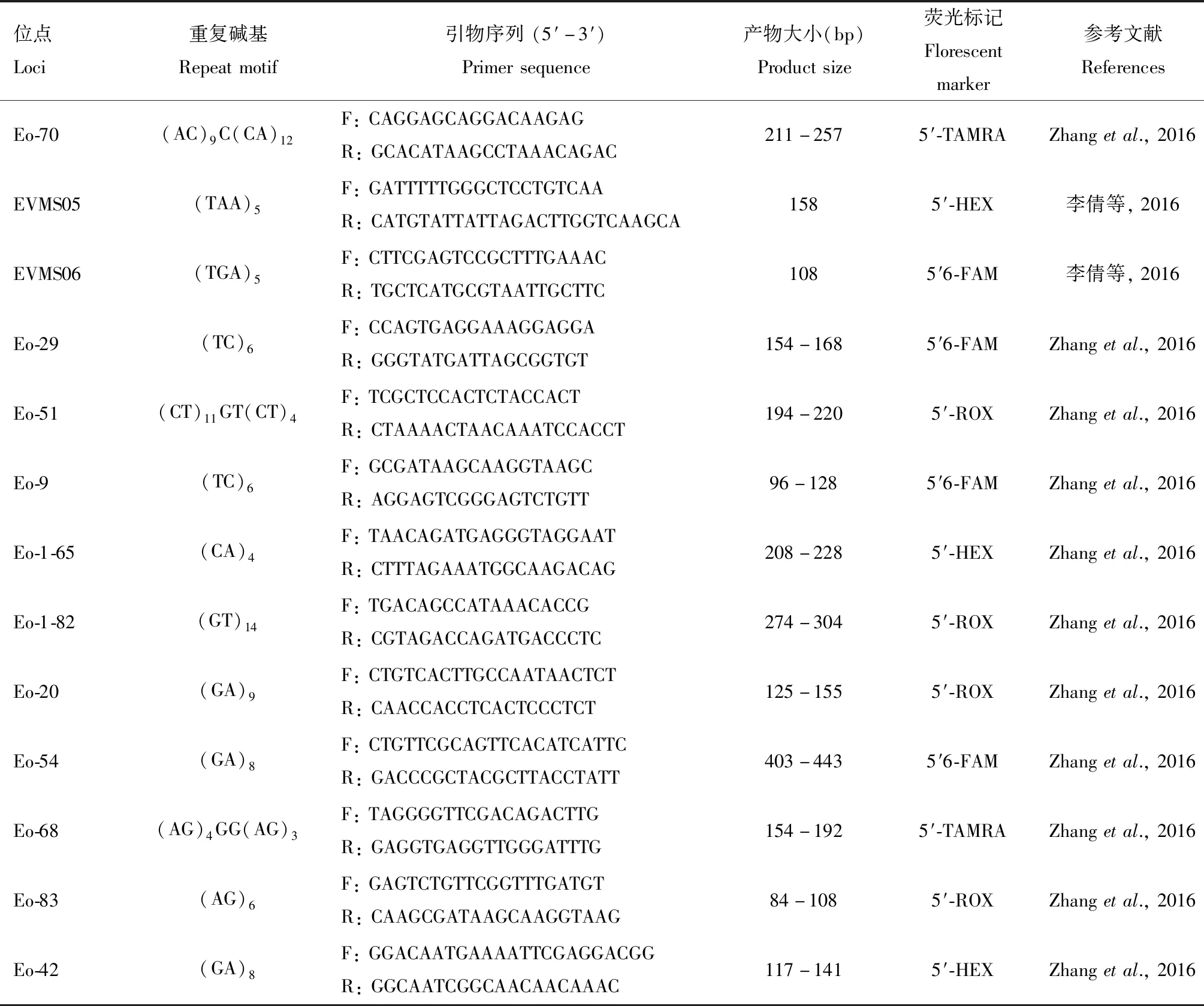
续表2 Table 2 continued
1.3 景观格局分析
为了定量化解析茶园周边景观格局特征与茶小绿叶蝉种群遗传信息之间的相关性,从地理数据云平台(http:∥www.gscloud.cn/)下载获取了2018年国家标准土地利用类型分类(land cover)原始影像图,其分辨率大约为30 m,包括了6大类国家标准的土地利用类型分类。在该影像图的基础上,结合谷歌地球(Google Earth)的投影影像和采样实地考察的照片资料,把研究区域内茶园单独勾绘出来,构建了包含茶园、林地、草地、耕地、水域、建筑和未利用(裸地、岩石等)7个土地利用类型的矢量影像资料。利用该矢量数据,我们对研究区域的景观组成和结构进行了分析。
使用ArcMap10.5分别生成统计了各采样点周围半径500,1 000,1 500,2 000和2 500 m内各类土地利用类型的面积占比(分析中将水域、建筑和未利用合并为一个无植被的土地利用类型统计),使用R(R Core Team,2018)中的cor函数对25个表示景观组成的变量进行了相关性检验,只保留没有明显共线性关系(r<0.8)的11个景观组成因子(表3)。同时,使用R软件包raster(Hijmans and van Etten,2016)将景观空间数据转化生成阻力距离曲面的栅格数据,以表示不同土地利用类型对茶小绿叶蝉扩散的阻力大小(Chenetal.,2019),分析获得代表景观结构的因子(表3)。此外,使用手持GPS(Garmin GPSMAP® 60CS)定位各采样点海拔高度和经纬度坐标(表1),并基于经纬度信息使用R软件包geosphere 1.5-10(Hijmansetal.,2017)计算采样点间地理距离,从而获得海拔和地理距离的因子(表3)。
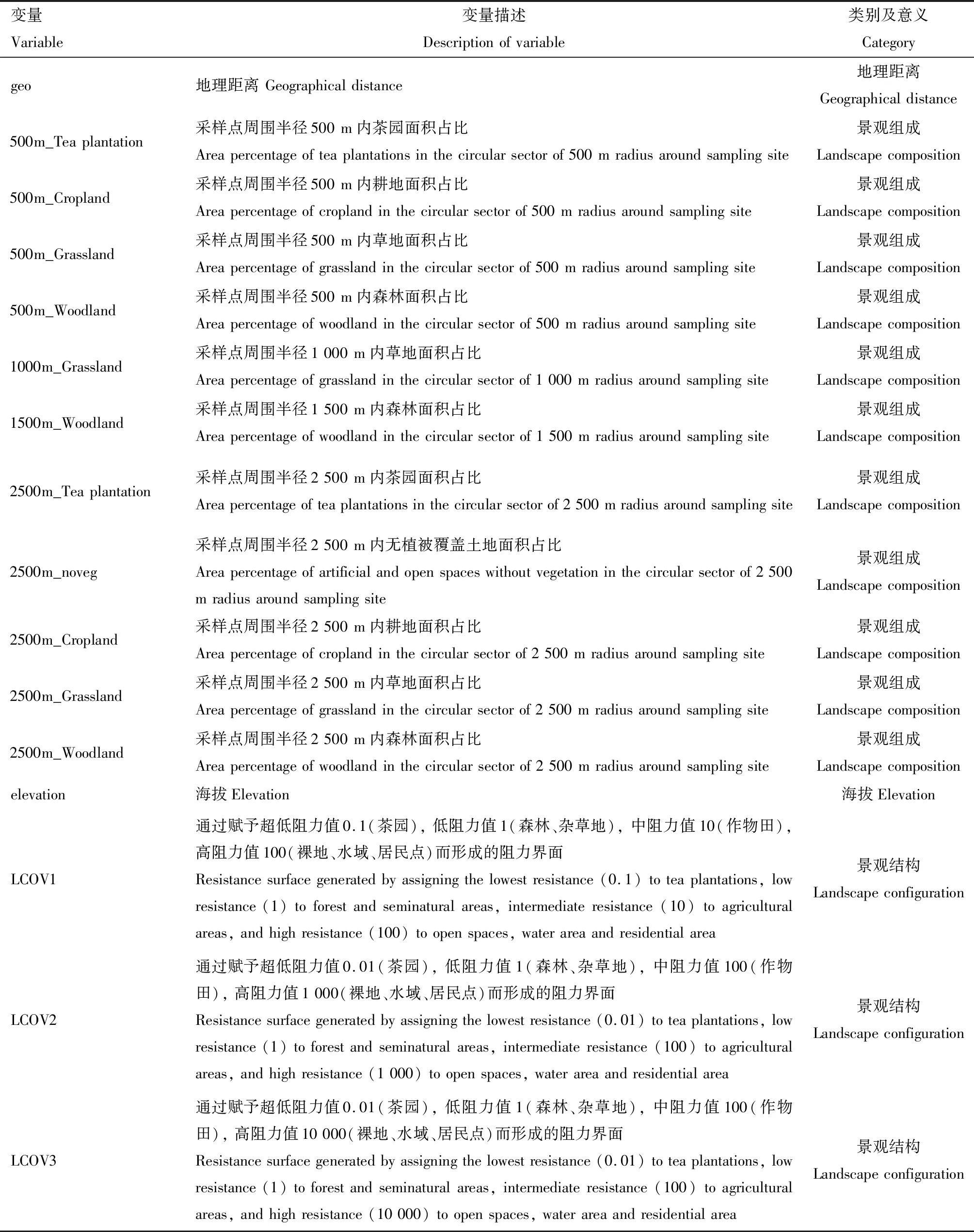
表3 本研究中所用的景观环境变量信息Table 3 Landscape and environmental variables used in this study
1.4 种群遗传多样性分析
利用ARLEQUIN 3.5.2(Excoffier and Lischer,2010)计算各种群期望杂合度(expected heterozygosity,He)和观察杂合度(observed heterozygosity,Ho),用FSTAT 2.9.3(Goudet,2001)计算各种群的基因多样性(gene diversity,DIV)和等位基因丰富度(allelic richness,AR)等遗传多样性参数。
1.5 种群遗传结构分析
使用STRUCTURE 2.3.4(Pritchardetal.,2000)对18个种群进行贝叶斯聚类分析。选择R软件包adegenet 2.0.0(Jombart,2008)中不基于模型假设的DAPC(discriminant analysis of principal components)(Jombartetal.,2010)程序解析个体之间的遗传关系。
1.6 种群遗传分化分析
利用R软件包adegenet 2.0.0中的dist.genpop函数计算种群间的遗传距离,包括Nei氏遗传距离(Nei’s genetic distance)(Nei,1972,1978)和Provesti氏遗传距离(Provesti’s genetic distance),并用ARLEQUIN 3.5.2(Excoffier and Lischer,2010)计算种群的遗传分化指数(pairwiseFST)和显著性检验P值。
1.7 种群基因流格局分析
使用BAYESASS 3.0.4(Wilson and Rannala,2003)估算样点之间茶小绿叶蝉种群最近几代的迁移率(m),即每个世代中迁移个体在种群中所占的比例。设置程序运行10次,每次进行20 000 000次重复的MCMC搜索(MCMC interaction),舍弃最初的2 000 000次重复的数据,每隔100次采样一次以确保m值的可靠、准确。
1.8 茶园周边景观格局与茶小绿叶蝉种群遗传的相关性分析
以11个景观组成因子和3个景观结构因子以及海拔(elevation)和地理距离(geo)(表3)为解释变量(explanatory variable),使用R软件包ecodist(Goslee and Urban,2007)分析其与茶小绿叶蝉种群的Provesti氏遗传距离、Nei氏遗传距离和遗传分化指数之间的Mantel相关性。
以16个景观环境因子(表3)为解释变量,以Provesti氏遗传距离、Nei氏遗传距离和遗传分化值为响应变量(response variable),使用R软件包ecodist的MRM函数进行距离矩阵多元回归(multiple regression on distance matrices,MRM)分析 (Wang,2013),以进一步确认与研究区域内茶小绿叶蝉种群Provesti氏遗传距离、Nei氏遗传距离和遗传分化指数有关的景观环境因子。
使用R软件包stats的一般线性模型(generalized linear mixed model,GLMM)分析采样点周围半径1 000 m范围内的草地面积占比 (1000m_Grassland) 和地理距离(geo)与茶小绿叶蝉种群Provesti氏遗传距离、Nei氏遗传距离和遗传分化指数的线性关系,并分析各样点1000m_Grassland值与茶小绿叶蝉种群遗传多样性(DIV,AR,He,Ho)的线性关系。
2 结果
2.1 种群遗传多样性
23个微卫星位点在18个种群中均表现出多态性,等位基因数量从9(Eo-1-57)到52(Eo-51)不等;等位基因丰富度(AR)介于3.686(FQ)和4.397(HL)之间,平均值为4.5195;基因多样性(DIV)介于0.676(FX)和0.734(HL),平均值为0.7081;期望杂合度(He)介于0.659(FQ)和0.729(HL),平均值为0.7567(表4),位于安溪县西部和北部样点的茶小绿叶蝉种群(如FQ,XG,HZ等)表现出较低的遗传多样性水平。
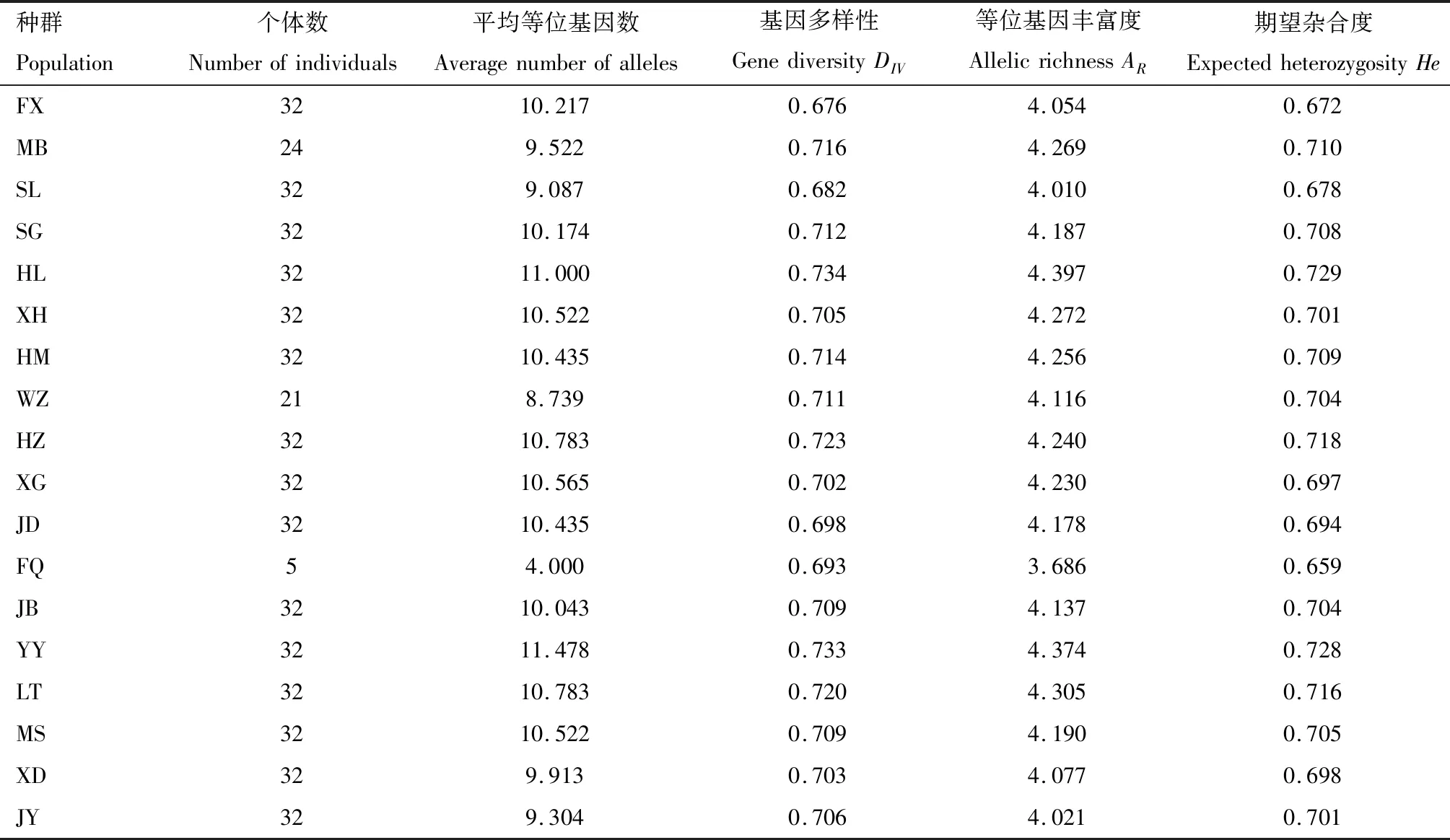
表4 福建安溪茶小绿叶蝉种群的遗传多样性Table 4 Genetic diversity of Empoasca onukii populations in Anxi,Fujian
2.2 种群遗传结构
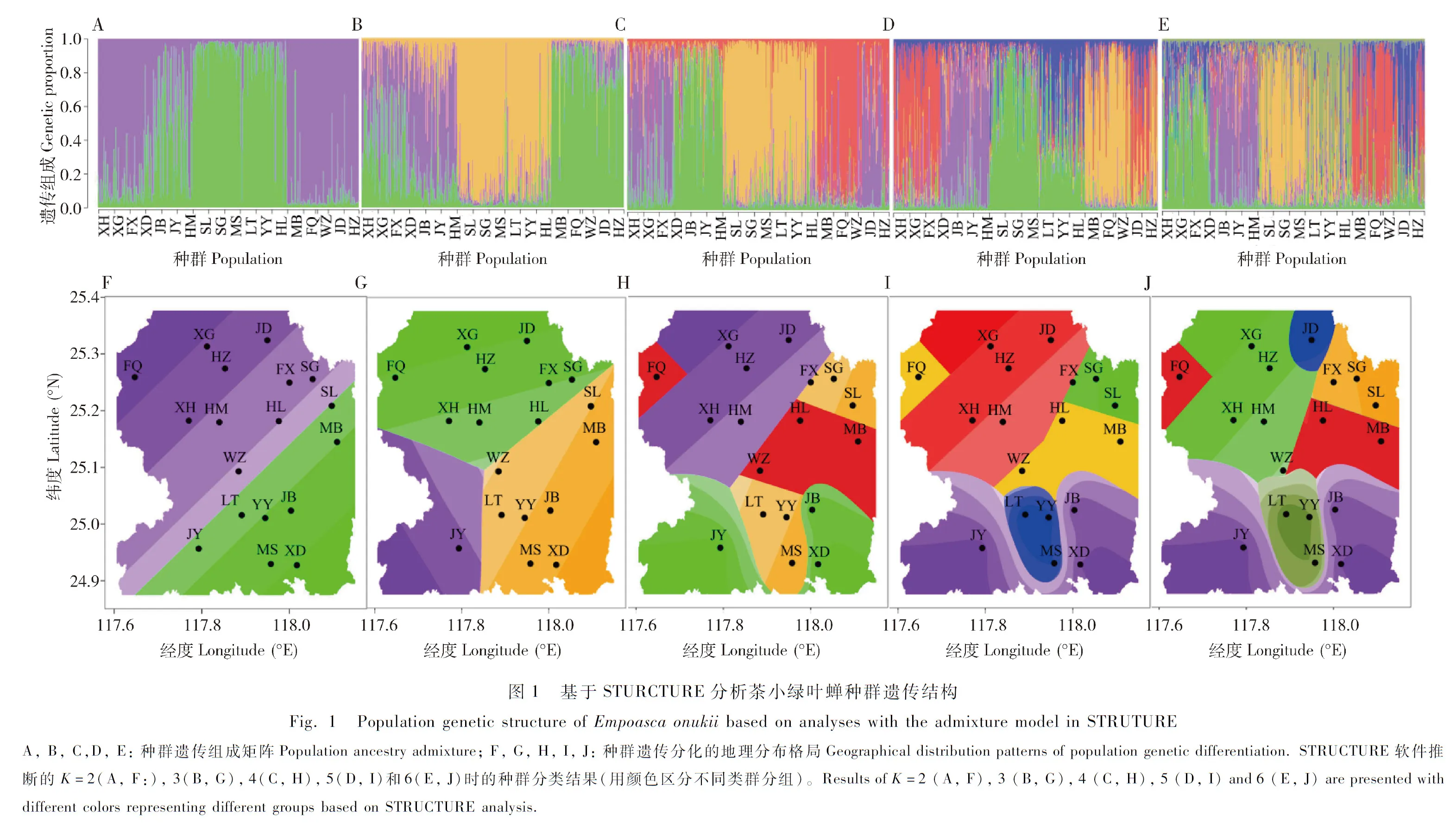
STRUCTURE软件推断的K的最大可能值是2,表明研究区域内茶小绿叶蝉种群可以划分为两个遗传类群(cluster):一个类群包括安溪县北部10个样点的种群,另一个类群包括安溪县南部8个样点的种群(图1:A,F)。在此基础上,K=3~6时,安溪县茶小绿叶蝉种群遗传分化格局比较明显(图1),大体上为位于安溪县西北部5个相邻样点(FQ,XG,HZ,XH和HM)的茶小绿叶蝉种群组成一个类群,分别位于北部和中部的JD和WZ种群各自单独成一个类群,位于东部的5个相邻样点的种群(FX,SG,HL,SL和MB)组成一个类群,位于南部的3个相邻样点的种群(LT,YY和MS)组成一个类群,同样位于南部的另外3个不相邻样点的种群(JY,JB和XD)组成一个类群。结合安溪县土地利用分类图来看,位于东部的5个样点(FX,SG,HL,SL和MB)以及FQ和WZ周围的茶叶种植较少,有较多的其他土地利用类型的分布,与其他样点之间处于相对隔离的状态(即与其他样点间没有适宜生境茶园的连接)。据此,我们推测周边景观格局不同的茶园中的茶小绿叶蝉种群遗传结构和基因交流模式可能存在差异。
不基于模型假设的DAPC分析将安溪县18个茶小绿叶蝉种群划分为4个类群(图2):JY,XD和JB为一个类群,FX,XG和XH为一个类群,FQ,MB,WZ,JD和HZ为一个类群,YY,MS,LT,SG和SL为一个类群。总体上仍然是南北种群分化明显。

图2 DAPC分析茶小绿叶蝉种群的遗传关系Fig.2 Genetic correlation analysis of Empoasca onukii populations using DAPC点代表每个分析个体,椭圆代表种群,椭圆中心点之间距离的大小代表种群亲缘关系的远近。Dots represent the study individuals,inertia ellipses represent the populations,and the distance between the center points of the ellipses represents the genetic distance between populations.
2.3 种群遗传分化
两个不同种群间的Nei氏遗传距离为0.042(MS-LT)~0.984(SL-FQ),Provesti氏遗传距离为0.207(MS-LT)~0.650(SL-FQ)(表5)。遗传分化指数为0.002(MS-LT)~0.222(SL-FQ),大部分均达到了统计学上的显著性水平(P<0.05)(表6)。根据Wright等(1978)提出的标准(0

表5 茶小绿叶蝉不同种群之间的Nei氏遗传距离(下三角)和Provesti氏遗传距离(上三角)值Table 5 Nei’s genetic distance (below diagonal) and Provesti’s genetic distance (above diagonal) values among different populations of Empoasca onukii
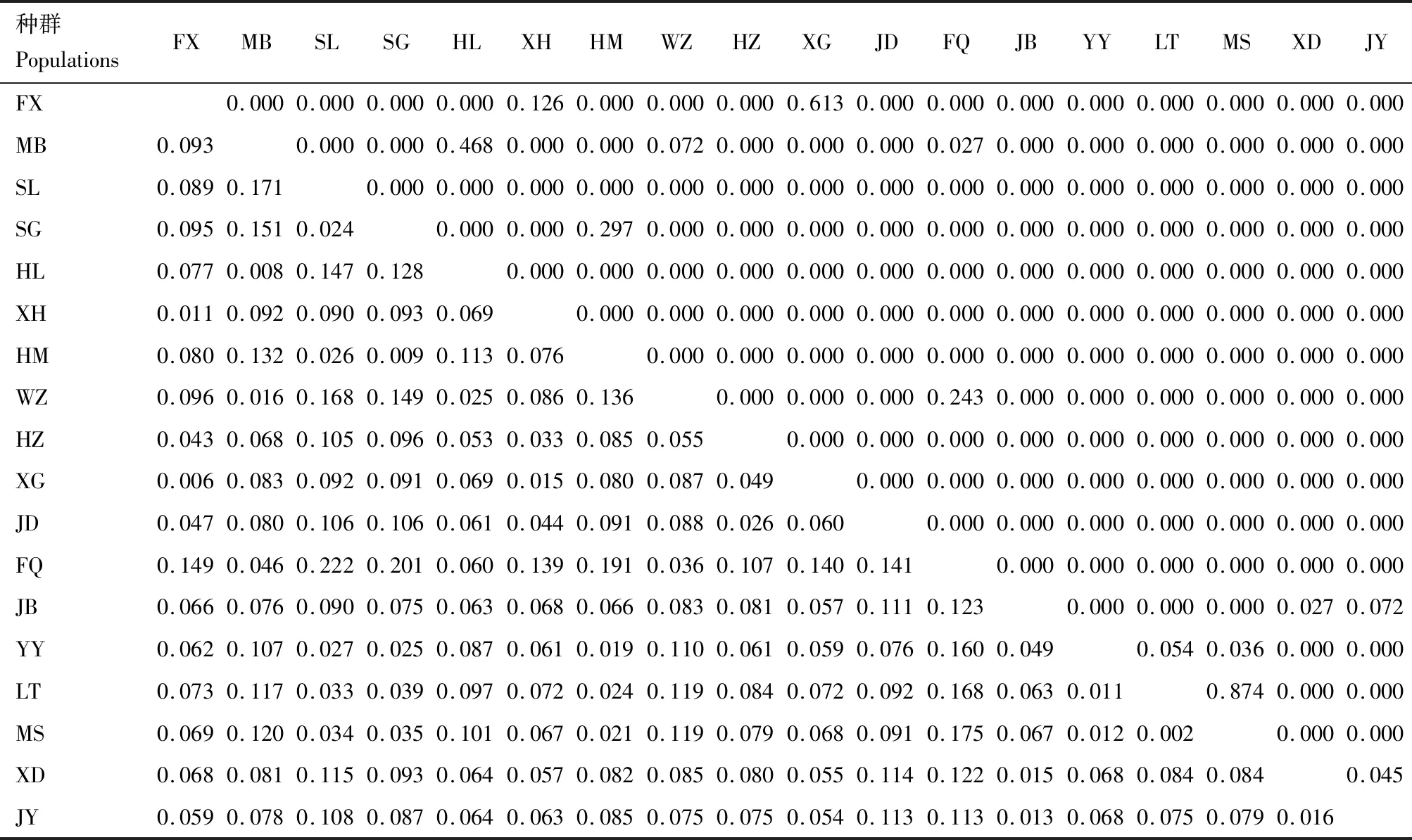
表6 茶小绿叶蝉不同种群之间的遗传分化指数值(下三角)及其统计检验值(上三角)Table 6 Pairwaise FST values (below diagonal) and their statistic P values (above diagonal) among different populations of Empoasca onukii
2.4 种群基因流格局
BAYESASS的多次单独运行获得了较为一致的结果,在检测的306对异地种群间,大部分种群间的现时基因流(Nm)很小,介于0.007~0.010之间(228对),其余70对介于0.010~0.097之间,8对介于0.108~0.180之间;而在种群内部个体之间基因流基本都很大,介于0.674~0.854之间(表7)。说明茶小绿叶蝉异地种群之间的现时基因交流可能受限,其现时基因流主要发生在种群内部个体之间。

表7 茶小绿叶蝉不同种群之间(上下三角)和种群内部个体之间(对角线)最近几代的基因流(Nm)值Table 7 Gene flow (Nm) values over recent generations among different populations of Empoasca onukii (below and above diagonal) and among individuals (on diagonal) in each population
2.5 茶园周边景观格局与茶小绿叶蝉种群遗传变异的相关性
Simple Mantel分析显示地理距离(geo)与Provesti氏遗传距离(r=0.2938,P=0.0088)、Nei氏遗传距离(r=0.2015,P=0.0455)以及遗传分化指数(r=0.2052,P=0.0301)之间均存在显著的相关性(表8)。在排除了地理距离(geo)的影响后,Partial Mantel分析显示,11个景观组成因子均与Provesti氏遗传距离、Nei氏遗传距离和遗传分化指数显著相关,而海拔(elevation)和与景观结构有关的所有因子均没有检测到与Provesti氏遗传距离、Nei氏遗传距离和遗传分化指数有显著相关性。

表8 景观因子与茶小绿叶蝉种群遗传分化的Simple Mantel和Partial Mantel相关性分析结果Table 8 Results of Simple Mantel and Partial Mantel tests among landscape factors and genetic differentiation of Empoasca onukii populations
距离矩阵多元回归分析表明,地理距离(geo)和采样点周围半径1 000 m范围内草地面积占比(1000m_Grassland)两个景观环境因子无论是组合在一起还是各自单独对茶小绿叶蝉种群Provesti氏遗传距离、Nei氏遗传距离和遗传分化指数的影响都是显著的;这两个因子对Provesti氏遗传距离、Nei氏遗传距离和遗传分化指数的解释比例有差异,决定系数R2分别为14.70%,7.72%和12.28%(表9)。

表9 距离矩阵多元回归分析相关的景观因子及其对茶小绿叶蝉种群遗传变异的解释比例Table 9 Related landscape factors and their explanatory proportion to the genetic variation of Empoasca onukii populations by multiple regression on distance matrix analysis
一般线性模型分析显示,采样点周围半径1 000 m范围内草地面积占比和地理距离与茶小绿叶蝉种群Provesti氏遗传距离、Nei氏遗传距离和遗传分化指数线性关系显著(图3),种群遗传距离随着地理距离的增加而显著变大(r=0.2015~0.2938,P=0.0002~0.0120)(图3:A,B,C),也随着采样点周围半径1 000 m范围内草地面积占比的变大而显著变大(r=0.1866~0.2604,P=0.0015~0.0226)(图3:D,E,F)。同时,采样点周围半径1 000 m范围内草地面积占比与茶小绿叶蝉种群He和DIV之间的线性关系也是显著的(图4:A,C),茶小绿叶蝉种群遗传多样性水平随着采样点周围半径1 000 m范围内草地面积占比的变大而变大(r=0.4829~0.5222,P=0.0267~0.0426)。
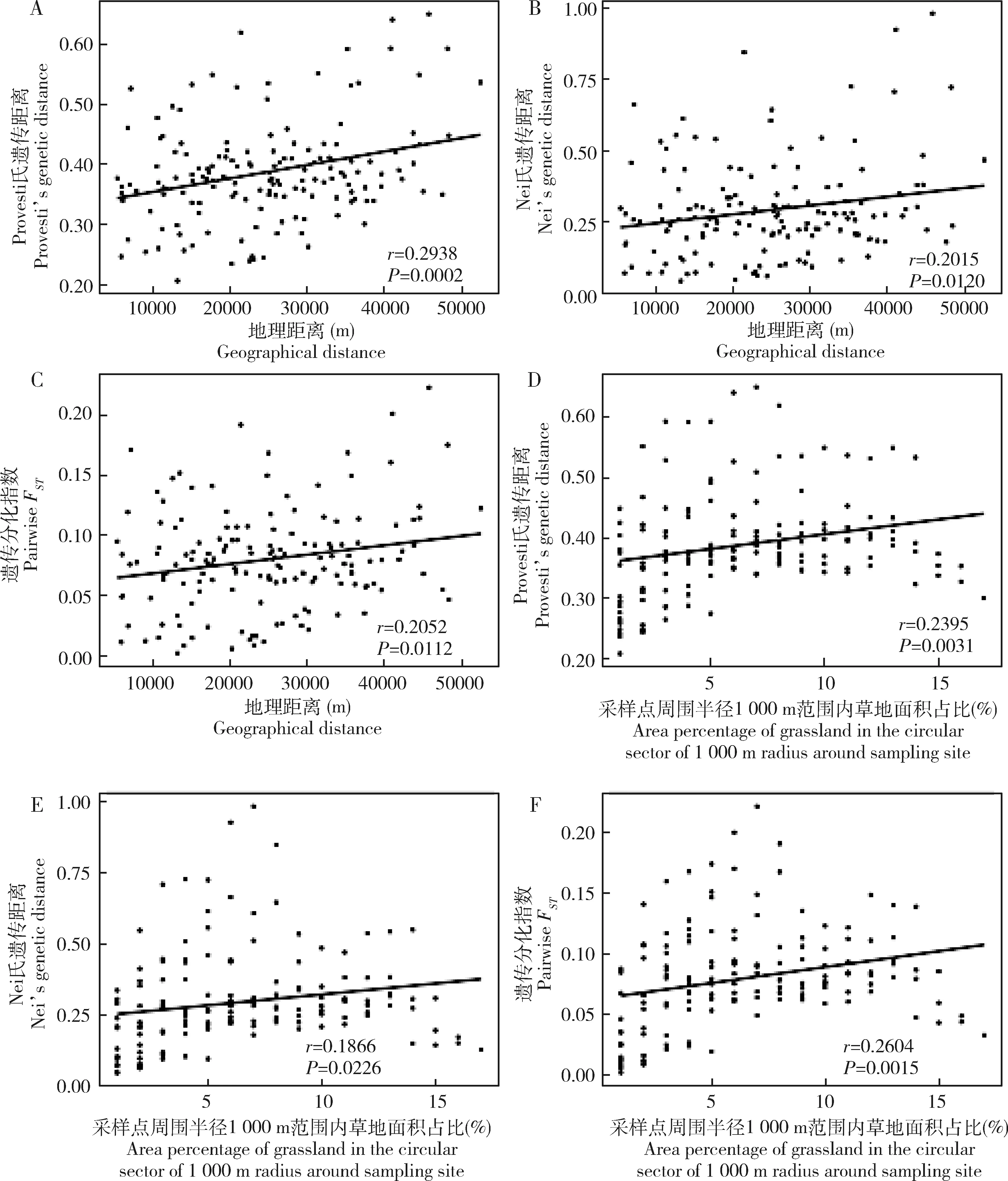
图3 一般线性模型分析茶小绿叶蝉种群Provesti氏遗传距离(A,D)、Nei氏遗传距离(B,E)和遗传分化指数(C,F)与地理距离和采样点周围半径1 000 m范围内草地面积占比的线性关系Fig.3 Linear correlation between Provesti’s genetic distance (A,D),Nei’s genetic distance (B,E) and pairwise FST (C,F) of Empoasca onukii populations and geographical distance and the area percentage of grassland in the circular sector of 1 000 m radius around sampling site based on generalized linear mixed model
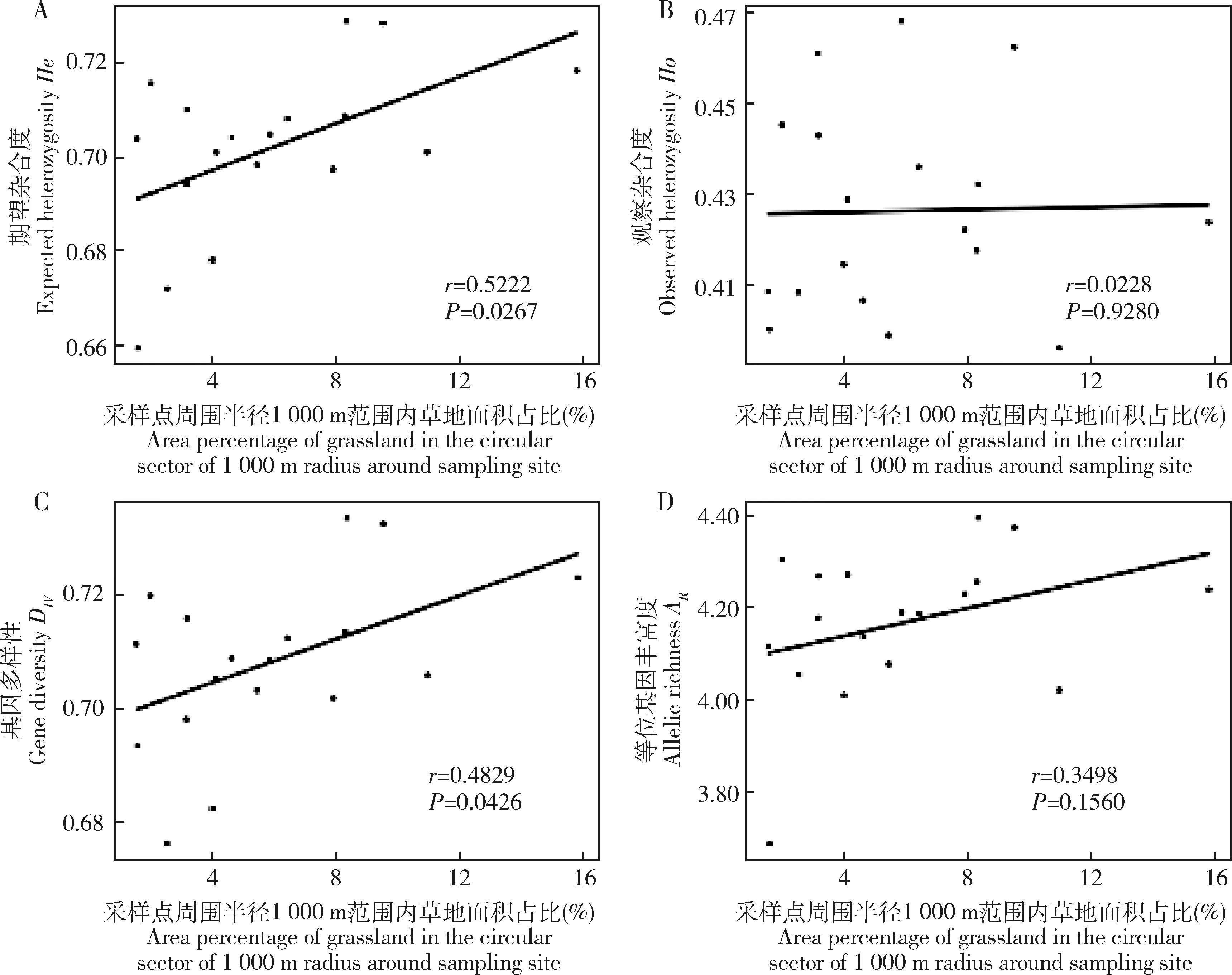
图4 一般线性模型分析茶小绿叶蝉种群He (A),Ho (B),DIV(C)和AR(D)与采样点周围半径1 000 m内草地面积占比的线性关系Fig.4 Linear correlation between He (A),Ho (B),DIV (C) and AR (D) of Empoasca onukii populations and the area percentage of grassland in the circular sector of 1 000 m radius around sampling site based on generalized linear mixed modelHe:期望杂合度Expected heterozygosity;HO:观察杂合度Observed heterozygosity;DIV:基因多样性Gene diversity;AR:等位基因丰富度Allelic richness.
3 讨论
3.1 茶小绿叶蝉的自主扩散能力
本研究基于23对微卫星位点的基因型数据对福建省安溪县18个茶小绿叶蝉种群进行了分析,虽然是小尺度区域的研究,基于模型假设和不基于模型假设的分析结果显示研究区域内茶小绿叶蝉种群遗传分化结构明显,遗传距离和分化指数的分析显示大部分种群间的遗传分化达到了中等水平,多元回归模型和一般线性模型以及Mantel分析显示茶小绿叶蝉种群遗传分化与地理距离显著正相关,种群表现地理距离隔离格局,这些信息表明茶小绿叶蝉种群有明显的遗传分化格局。近期有其他研究者报道了基于微卫星数据分析茶小绿叶蝉在中国四大茶区明显的种群遗传结构和地理距离隔离格局(Zhangetal.,2019)。Lo等(2019)基于微卫星标记发现澳大利亚16个榈椿Thaumastocorisperegrinus种群表现中等分化和地理距离隔离格局并据此推测榈椿的自主扩散能力有限;Cao等(2019)对15个瓜蓟马Thripspalmi种群的微卫星基因型分析发现种群表现出显著的遗传分化和地理距离隔离格局,基于BAYESASS估算的种群间的现时基因流明显受限,推测种群间可能没有持续的扩散事件。凡是具有远距离和大范围扩散的迁飞性昆虫种群,往往不具有明显的遗传结构,异地种群间通常可以检测到明显的基因流。例如,迁飞性害虫小菜蛾Plutellaxylostella种群的遗传分化不明显,具有明显的基因流格局(Weietal.,2013,2017);而采自相同区域的菜蛾盘绒茧峰Cotesiavestalis的种群间却表现出很强的遗传分化水平(Weietal.,2017),因此认为菜蛾盘绒茧峰可能不会跟随其寄主远距离迁飞。Zhang等(2019)虽然未对基因流进行直接的估算与分析,但认为茶小绿叶蝉不同地理种群之间存在大量基因流。本研究基于BAYESASS对茶小绿叶蝉异地种群间的迁移率(基因流)进行了估算,BAYESASS估算的迁移率(m)通常被认为是种群5代以内(<5代)的情况(Wilson and Rannala,2003),鉴于茶小绿叶蝉种群在中国茶区每年发生9~16代(Fuetal.,2014),本研究估算的基因流可以反映茶小绿叶蝉种群“当代”(不到一年)的迁移扩散事件,也即帮助我们推测茶小绿叶蝉种群的扩散过程和迁移能力。BAYESASS分析显示其研究区域内茶小绿叶蝉种群间的当代基因交流受限,其现时基因流往往仅限于种群内部个体之间,表明小尺度区域范围内茶小绿叶蝉异地种群间可能缺乏自主的扩散和交流过程。根据以上信息,我们推测茶小绿叶蝉很可能不具有远距离迁飞的习性,自主扩散的能力有限。周宁宁(2014)和陈世春等(2015)分别基于线粒体COI和16S rRNA基因序列对假眼小绿叶蝉Empoascavitis地理种群的分析也都认为茶小绿叶蝉可能是一类扩散能力有限的害虫。
尽管茶小绿叶蝉的自主扩散能力有限,人类活动对茶小绿叶蝉种群的影响是不可忽视的,其中最主要的是茶苗在异地间的调运和移植过程可能可以为茶小绿叶蝉种群实现跨地区扩散和远距离传播创造很好的条件和机会。在过去几十年中,随着无性繁殖技术在茶树上的运用和发展,由于其相对于茶籽播种来说更加快速高效,越来越多新茶园的建立选择从异地调运茶苗,这为茶小绿叶蝉种群实现跨地区扩散和远距离传播创造了很好的条件和机会。茶小绿叶蝉种群具有跟随寄主茶苗实现跨地区扩散和远距离传播的能力,茶小绿叶蝉通常将卵产入茶枝嫩梢或叶梢组织中,其成虫和若虫也很小(Shietal.,2015),又具有久坐不动和大部分时间均栖息在叶背或嫩梢上吸食汁液为害的习性(边磊等,2014)。在种植者没有更多农事操作和育苗知识的情况下,茶苗很可能受到茶小绿叶蝉侵染,并使其跟随茶苗异地调运而实现远距离传播和异地扩散。因此,新建茶园仍要关注茶小绿叶蝉侵染和危害的问题。随着未来新茶园的发展和旧茶园的替换,茶苗和茶树在异地间的调运应进行严格的检查以防止茶小绿叶蝉种群在更大区域范围内的扩散和传播。同时,相比于扦插等无性繁殖技术,组培和茶籽播种可以繁育出不带虫卵的茶苗,结合将这些“无虫”苗木在温室中培育的措施可以帮助减少潜在的茶小绿叶蝉侵染的风险,并避免更大范围的传播和扩散。
虽然人类活动影响不可忽视也不可控,但茶树是多年生作物,一经种植可以多年保持,因此在已经建立的成熟茶园中,生态系统相对于其他作物系统更稳定,人类活动对茶小绿叶蝉的助迁作用和机会会大大减少。因此研究中我们选择其中茶树具有一定树龄的茶园采集样本以避免近期茶苗移植对分析的影响,同时选用微卫星分子标记和BAYESASS分析种群遗传信息。一般认为高度可变的位点,如微卫星分子标记适合于研究和反映生物种群短期的小尺度区域的遗传结构和基因流(Wang,2010,2011)。而基于频率分配检测的方法BAYESASS被广泛用于估算当前或以前几代的迁移率来推断最近的基因流动。由此研究中分析的当代种群遗传分化结构和基因流空间格局主要反映茶小绿叶蝉种群的自主扩散或迁飞能力信息,而可能与茶苗异地调运过程关系较少。
3.2 茶园周边景观格局对茶小绿叶蝉种群遗传结构的影响
本研究中,研究区域内茶小绿叶蝉种群大体上分为东部、西北部和南部种群的格局,结合茶园周边景观格局分析结果和采样观察记录以及相关文献记录(税伟等,2017),位于南部的JB,LT和YY等种群所在乡镇,主要采用单一化和集约化的种植模式,在茶园周围少见自然和半自然生境或其他作物田;位于西北部的FQ和XG等种群所在乡镇,茶园大都分布于高海拔的山坡上,周围经常会有大量森林等自然和半自然生境分布。而位于东部的FX,SG,HL,SL和MB等种群所在的乡镇,种植模式则会更多样化,茶园周围经常会有更多居民点和其他作物田的分布。根据这些信息,我们推测周边景观格局不同的茶园中的茶小绿叶蝉种群遗传结构和基因交流模式可能存在差异。
借助于景观遗传学的原理和方法,使用Mantel相关性分析、距离矩阵多元回归模型MRM和一般线性模型GLMM定量化解析了地理距离、海拔、样点周围不同尺度范围的景观组成因子以及景观结构与茶小绿叶蝉种群遗传变异的相关性。结果显示地理距离(geo)和采样点周围半径1 000 m范围内草地面积占比(1000m_Grassland)是与研究区域内茶小绿叶蝉种群遗传分化显著相关的因子,茶小绿叶蝉种群遗传分化水平随着采样点间地理距离的变大而变大,也随着采样点间周围草地面积差异值的变大而显著变大,与周围草地面积较小的茶园相比,有较多草地分布的茶园中的茶小绿叶蝉种群表现出较高的遗传多样性水平。类似地,Dong等(2018)的研究发现棉蚜Aphisgossypii的遗传多样性水平与1 000 m和2 000 m范围内的草地面积比例呈显著正相关。已有的大量景观生态学研究表明一些生境(special habitat)或土地利用类型(land cover)会影响昆虫的定殖过程(Dongetal.,2015;Angelellaetal.,2016)。因此,景观组成会影响昆虫种群的建立和扩张过程,进而也可能影响其种群的遗传结构和分化(Dongetal.,2018),特别是对于这类自主扩散能力有限的昆虫来说,景观组成的影响作用可能会更明显。
本研究中的土地利用类型空间分布格局可以简单反映茶小绿叶蝉的栖境和寄主植物以及不适宜生境的分布。已有的研究还不能排除茶小绿叶蝉会在杂草上越冬、过渡和繁殖的可能性(陈李林等,2019;Chenetal.,2019)。杂草地对茶小绿叶蝉种群的遗传多样性具有积极作用,可能一方面茶小绿叶蝉种群在茶园受到农事操作(如茶枝修剪和农药喷洒等)干扰时移动到杂草地过渡,干扰解除后再从杂草地移回茶园栖息地,景观组成可能通过这样的方式影响其种群的建立、定殖过程,进而影响其遗传结构和分化;另一方面,杂草地生境本身也可能提供丰富的叶蝉资源,寄主和替代寄主分布通过选择和繁殖影响茶小绿叶蝉种群的遗传结构和分化。相比于杂草地,森林生境包含更多的多年生乔木树种,这些高大的树种可能大多数都不能成为茶小绿叶蝉合适的寄主植物,而且随着森林面积的增加,适合于茶小绿叶蝉种群栖息的茶园面积减少,同时也不能排除森林生境中的高大树种会阻断茶小绿叶蝉种群间的交流、扩散和连续性的可能性。因此,在周围有更多森林生境分布的样点中的种群如FQ和XG等,叶蝉种群表现较低遗传多样性水平,一般线性模型的分析也显示叶蝉种群遗传多样性水平与茶园周围的森林生境面积呈负相关(数据未展示)。赵紫华等(2010)报道与麦田相邻的自然和半自然生境栖境可有效地延缓麦蚜种群迁入作物田的时间并减少扩散传播事件。从本研究的结果来看,不同的自然生境类型对害虫种群的影响作用有差异。就茶小绿叶蝉而言,茶园周围的杂草地生境对其种群建立、过渡和扩张可能是有利的,因此清除杂草可能是茶小绿叶蝉可持续治理策略很重要的一项内容。当然,非作物生境通常也被认为可能栖息着大量可作为生物防治资源的天敌物种(Tscharntkeetal.,2012;Gurretal.,2016),茶小绿叶蝉有两种主要的卵寄生蜂叶蝉三棒缨小蜂Stethyniumempoascae和微小裂骨缨小蜂Schizophragmaparvulas在田间的寄生率超过30%,被认为是茶小绿叶蝉可持续控制的重要天敌(李慧玲和林乃铨,2008;毛迎新等,2008),那么杂草地生境对这两种寄生蜂是否有益?如果有益,那杂草地生境对茶小绿叶蝉种群的促进作用和在保育天敌资源以更好发挥防控效能作用间的比较和权衡也是一个值得探讨的复杂问题。
此外,景观尺度可能也对茶小绿叶蝉种群的扩散和定殖有一定的影响,本研究在5个空间尺度(景观半径0.5~2.5 km不等)上研究了景观组成因子对茶小绿叶蝉种群遗传多样性和变异的影响,结果显示茶园周围1 000 m半径内的草地面积解释了茶小绿叶蝉种群遗传多样性和变异的最大比例。尽管当前我们对茶小绿叶蝉种群的移动特性还了解得不多,但茶园周围1 000 m半径这个尺度范围的景观可能包含了茶小绿叶蝉种群的生态邻域(ecological neighborhood)(Dongetal.,2018),其中可能包含或者满足了其可以或需要利用的生境尺度。
综合以上信息来看,关于小尺度区域范围内茶小绿叶蝉景观遗传学的研究可能有助于增进对茶小绿叶蝉定殖及其相应空间尺度和相关景观格局的理解。但是,生物种群遗传变异是一个复杂的过程,微卫星分子标记(SSR)的统计功效也还比较有限,研究工作还比较初步,16个空间因子之外,可能还有其他相关的影响因子没有被检测或识别到。未来的工作可以考虑对茶小绿叶蝉种群及其相关天敌昆虫种群在茶园景观中的生态学过程进行更多的观察和研究,如检查可能充当茶小绿叶蝉替代寄主的植物物种、测量从茶园栖境迁出和从过渡栖境迁回茶园的最近(远)距离等等,以进一步确认茶园周边景观格局对茶小绿叶蝉种群建立、定殖、扩张的影响。
猜你喜欢
中国临床医学影像杂志(2022年5期)2022-07-26
烟台大学学报(自然科学与工程版)(2022年3期)2022-06-30
水产科学(2022年2期)2022-03-20
华人时刊(2021年17期)2021-12-02
南京师范大学学报(工程技术版)(2021年2期)2021-10-20
心声歌刊(2021年6期)2021-02-16
海峡姐妹(2020年3期)2020-04-21
创新作文(5-6年级)(2019年10期)2019-12-23
湘潮(上半月)(2019年3期)2019-05-22
娃娃画报(2018年3期)2018-08-15
