
大麦重组自交系群体籽粒总花色苷含量和千粒重QTL定位
2020-12-11 02:59:24杨晓梦普晓英MuhammadKazimAli杨加珍曾亚文
作物学报 2020年1期
杨晓梦 李 霞 普晓英 杜 娟 Muhammad Kazim Ali杨加珍 曾亚文,* 杨 涛
大麦重组自交系群体籽粒总花色苷含量和千粒重QTL定位
杨晓梦1,2,**李 霞1,2,**普晓英1,2杜 娟1,2Muhammad Kazim Ali3杨加珍1,2曾亚文1,2,*杨 涛1,2
1云南省农业科学院生物技术与种质资源研究所, 云南昆明 650205;2云南省农业生物技术重点实验室, 云南昆明 650205;3卡拉奇乌尔都艺术与科技大学生物技术系, 卡拉奇 75300, 巴基斯坦
以云南特有的紫色大麦紫光芒裸二棱和澳大利亚引进的黄色大麦Schooner构建的193个重组自交系为材料, 对 2013—2015年3年3个试验点的大麦籽粒总花色苷含量和千粒重进行相关性分析和QTL定位。大麦总花色苷含量和千粒重之间呈显著或极显著负相关。共检测到12个总花色苷含量QTL, 分别位于1H、2H、4H、6H和7H染色体, 贡献率为5.06%~23.86%; 8个千粒重QTL, 分别位于2H、4H和7H染色体, 贡献率为4.67%~42.32%。贡献率大于10%的QTL有10个, 大于20%的有5个, 最大的可达42.32%。其中至少2年2点重复检测到2个总花色苷含量QTL, 分别位于2H Bmag0125–GBM1309和7H EBmatc0016–Bmag0206区间, 可分别解释表型变异的13.66%~17.76%和13.07%~16.43%; 3年3点重复检测到2个千粒重QTL, 分别位于2H Scssr03381–scssr07759和7H GBM1297−GBM1303区间, 可分别解释表型变异的4.67%~14.55%和34.51%~42.32%, 其加性作用方向均一致。控制总花色苷含量与千粒重的主效QTL同位于2H和7H染色体。
大麦重组自交系; 总花色苷含量; 千粒重; 相关性; QTL定位
有色大麦是人类健康关键的功能成分和次生代谢产物的重要来源, 具有减轻一些人类慢性病(心血管疾病、糖尿病等)的活性物质, 其多种生物活性都与花色苷有关[1-3]。花色苷是植物在长期适应环境衍生的一类自我保护的次生代谢产物[4], 通过其较强的抗氧化作用清除自由基对预防心血管疾病和促进人类身体健康有至关重要的作用[3]。有色大麦在理想保健食品[5-6]、食品安全添加剂[7]、化妆品安全染色剂[8]等多方面具有巨大的开发潜力。此外, 提高产量仍然是当前和未来大麦育种的首要目标, 而千粒重是大麦理论产量主要决定因素之一[9], 可从侧面反映出不同材料的适应、稳定及丰产属性, 决定不同材料在特定区域的产量, 是选育高产品种的主要依据。因此, 研究有色大麦花色苷和千粒重的遗传分子机制对高花色苷大麦优质育种及高产有着重要的理论和实践意义。
花色苷和千粒重一般是由多基因控制的数量性状, 目前已有一些相关研究报道。127种基因型不同颜色大麦籽粒花色苷含量在13.0~1037.8 μg g–1之间, 紫色和蓝色大麦花色苷含量显著高于黑色大麦[10]。研究籽粒颜色的分离比, 发现籽粒颜色是由一个或两个基因位点控制[11-12]。不同颜色的大麦籽粒相关基因被定位于不同染色体。控制黑色大麦的显性基位于1HL染色体, 决定黑色深浅程度[13]。蓝色大麦籽粒颜色由5个基因控制(、和基因位于4HL、和位于7HL染色体)[14]。而控制蓝色大麦糊粉层花色苷合成的候选基因有3个, 分别为(4HL)、(4HL)和(6HL)。其中,是影响颜色变化的主要基因, 且所在区域也发现基因簇和花色苷合成结构基因()[15]。大多研究报道控制紫色大麦的相关基因位点(如和[16-17]、[11][18]等)主要位于2H染色体。千粒重与不同大麦材料、气候因子及环境条件等紧密相关。有研究表明千粒重与粒长、粒宽、粒厚均存在显著或极显著正相关, 且二棱大麦显著高于六棱大麦[19]。张小燕等[20]、李喜焕等[21]及吕毅[22]发现国内、外不同大麦材料千粒重变幅为10.20~65.5 g, 其中大于60 g的属于稀有大粒种质。王建林等[23]从大尺度上研究了青藏高原栽培大麦千粒重的空间分布规律, 揭示影响千粒重最大环境因子为抽穗-成熟期降水量, 其次为土壤速效钾含量, 最小影响因子为地理经度。大麦千粒重遗传受多个效应不同的QTL控制[24], 不同学者在不同材料中检测到的千粒重QTL在大麦7条染色体上都有报道[25-27]。此外, 大麦产量相关基因与水稻的基因同源, 减少该类基因会增加开花期的各组织细胞分裂素, 使生殖器官细胞的数目增多进而提升产量[28]。
一直以来, 因定位群体、鉴定方法和环境差异所获得的表型差异, DNA标记和统计方法不同, 加之基因与环境互作等, 使所检测到的QTL数量和座位不同, 限制了大麦花色苷和产量相关QTL精细定位和克隆。继续挖掘不同遗传资源中的花色苷含量与产量相关QTL对于了解它们的遗传机制有着重要意义, 同时为遗传育种提供更多的优异基因资源。以同一个大麦群体同时检测花色苷含量和千粒重QTL的报道以及花色苷含量与产量性状相关性的研究鲜见报道。本研究对亲本间差异明显的总花色苷含量和千粒重进行QTL定位, 分析花色苷含量与产量性状之间的相关性, 为进一步的精细定位、克隆和分子辅助育种提供依据。
1 材料与方法
1.1 试验材料
所用大麦材料为母本紫光芒裸二棱(云南特有的地方青稞品种, 成熟时全株紫色)和父本Schooner (澳大利亚优质啤酒大麦, 成熟时全株黄色)杂交衍生的193个F9~F11重组自交系。
1.2 试验方法
1.2.1 田间种植及性状考察 2013、2014和2015年秋季, 分别在云南玉溪、白邑和嵩明试验基地种植亲本及其F9~F11RIL群体的193个株系。试验地肥力均匀, 随机区组排列, 行长2 m, 行距为0.3 m, 2次重复, 常规田间管理。待群体植株成熟后, 室内称重统计千粒重。
1.2.2 大麦籽粒总花色苷含量的测定 参照PH示差法[29-30]测定大麦籽粒总花色苷含量。
1.2.3 统计分析 利用SPSS20.0软件对总花色苷含量和千粒重进行数据统计及相关分析。
1.2.4 QTL定位 利用本实验室已构建的大麦SSR标记遗传连锁图谱[31]定位QTL, 采用QTL Ici- Mapping V3.3软件进行完备区间作图分析(Inclusive Composite Interval Mapping, ICIM)[32-33], 以LOD≥2.5作为QTL存在的阈值, 确定总花色苷含量和千粒重QTL及估算遗传效应。
2 结果与分析
2.1 大麦总花色苷含量和千粒重表型分析
由表1可知, 该群体亲本间及其RIL之间总花色苷含量均存在极显著差异(<0.01), 千粒重存在显著(<0.05)或极显著差异(<0.01)。母本紫光芒裸二棱在总花色苷含量上属于高值亲本, 父本Schooner属于低值亲本; 反之, 紫光芒裸二棱在千粒重上属于低值亲本, Schooner属于高值亲本。RIL群体间总花色苷含量和千粒重具有广泛的变异幅度。总花色苷含量偏度和峰度绝对值基本>1, 呈单峰分布(图1), 而千粒重两者绝对值基本<1, 近似正态分布(图1)。从频率分布趋势可看出各性状有一定超亲分离现象, 具数量性状特征, 适合基因定位分析。

表1 大麦籽粒总花色苷和千粒重表型值
数值后不同大写字母表示RIL群体在不同试验点间差异在0.01水平上显著。
<
Different capital letters following the values indicate significant difference among RIL populations at different sites at the 0.01 probability level. TAC: total anthocyonin content; TKW: 1000-kernel weight; ZGMLEL: Ziguangmangluoerleng.
2.2 大麦总花色苷含量和千粒重的相关分析
如表2所示, 3个试验点间大麦籽粒总花色苷含量(0.918**、0.873**、0.833**)和千粒重(0.832**、0.775**、0.720**)均呈极显著正相关。表明该群体总花色苷含量和千粒重主要受基因型控制。大麦总花色苷含量与千粒重基本呈显著或极显著负相关, 说明在一定程度上, 籽粒千粒重越小, 总花色苷含量可能越高。
2.3 大麦总花色苷含量和千粒重的QTL定位
对玉溪(2013年)、白邑(2014年)和嵩明(2015年)试验点大麦籽粒总花色苷含量及千粒重, 共定位到20个QTL。分布于除3号、5号染色体外的其余5条染色体上, LOD值介于2.50~26.14, 分别解释相应表型变异的4.67%~42.32%, 贡献率大于20%的QTL有5个(表3和图2)。玉溪点, 定位于1H、2H、4H、6H和7H 等5条染色体上的总花色苷含量和千粒重QTL共8个(表3和图2)。1H和6H染色体上分别发现1个QTL, 2H、4H和7H染色体上各发现2个QTL。其中, 控制总花色苷含量的QTL有5个, 分别位于1H、2H、4H、6H和7H染色体。LOD值为4.04~10.42, 表型变异率为5.91%~13.66%。单个QTL贡献率没有超过20%的, 但5个QTL总贡献率达到了 46.55%。除和加性效应为负值外, 增效等位基因源于亲本Schooner; 其余3个QTL的加性效应为正值, 增效等位基因来自亲本紫光芒裸二棱。控制千粒重的QTL有3个, 分别位于2H、4H和7H染色体。LOD值为3.81~26.14, 表型贡献率为4.73%~42.32%。的加性效应为正值(1.396), 增效基因来自紫光芒裸二棱; 其余2个主效QTL的加性效应值为负值, 增效基因来自Schooner。位于7H 染色体的表型贡献率高达42.32%, 是一个来源于Schooner的主效位点。

图1 大麦籽粒总花色苷含量和千粒重的次数分布
横坐标分别代表3年3个试验点(玉溪2013、白邑2014和嵩明2015年)大麦RIL群体总花色苷含量和千粒重; 纵坐标分别代表大麦RIL群体总花色苷含量和千粒重的株系数。
The abscissa represents total grain anthocyain content and 1000-kernel weight in barley RIL population grown at three sites (Yuxi, Baiyi, Songming) for three consecutive years (2013−2015), respectively; ordinate represents number of lines in total grain anthocyain content and 1000-kernel weight, respectively.
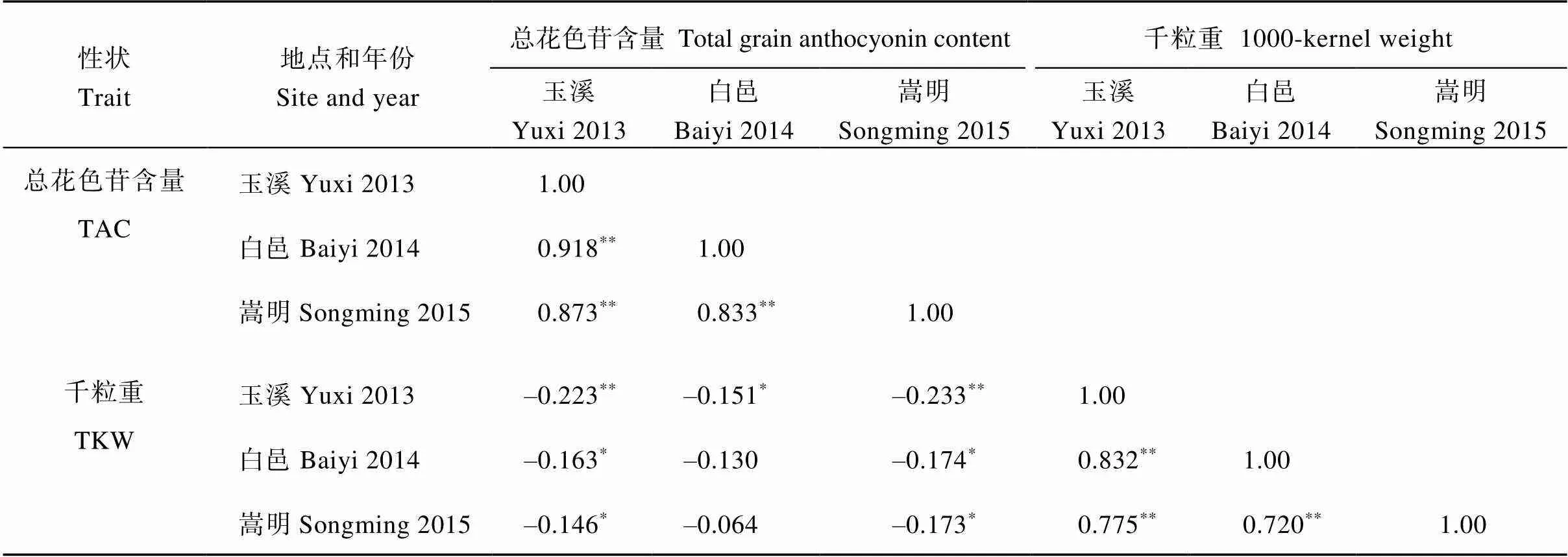
表2 大麦籽粒总花色苷含量和千粒重的相关系数
*和**分别表示在0.05和0.01水平上显著相关。各性状缩写见表 1。
*and**indicate significant correlation at the 0.05 and 0.01 probability levels, respectively. Abbreviations of traits correspond with those given in Table 1.

表3 大麦籽粒总花色苷含量和千粒重的QTL分析
加性效应正值和负值分别表示增效等位基因来自紫光芒裸二棱和Schooner。各性状缩写见表1。
Positive value and negative value of additive indicate that the positive allele came from ZGMLEL and Schooner, respectively. Abbreviations of traits correspond with those given in Table 1.
白邑点, 定位于4条染色体2H、4H、6H和7H上的总花色苷含量和千粒重QTL共7个(表3和图2)。2H染色体上发现3个QTL, 4H和6H染色体上分别发现1个 QTL, 7H染色体上发现2个 QTL。其中, 控制总花色苷含量的QTL有4个, 分别位于2H、4H、6H 和 7H。LOD 值为3.58~11.38, 表型变异率为5.06%~17.76%。单个QTL贡献率没有超过20%的, 但4个QTL总贡献率达到了45.74%。的加性效应为–0.085, 增效基因源于Schooner, 其余3个QTL的加性效应为正值, 增效基因来自紫光芒裸二棱。控制千粒重的QTL有3个, 分别位于2H和7H, 其增效等位基因均来自Schooner。LOD值为2.75~18.73, 表型贡献率为4.67%~34.51%。位于7H染色体的表型贡献率高达34.51%, 是一个来源于Schooner的主效位点。
嵩明点, 定位于2H、6H和7H染色体上的总花色苷含量和千粒重QTL共5个(表3和图2)。2H和7H染色体上分别发现2个QTL, 6H染色体上发现1个 QTL。其中, 控制总花色苷含量的QTL有3个, 分别位于2H、6H和7H。LOD值为3.77~12.16, 表型变异率为6.24%~23.86%。除的加性效应为–0.104外, 是增效基因源于Schooner的主效位点(贡献率为23.86%)。位于2H染色体的表型贡献率达20.09%, 是源于紫光芒裸二棱的主效位点。控制千粒重的QTL有2个, 分别位于2H和7H, 其增效等位基因均来自Schooner。其中位于7H上的主效QTL, LOD值为17.37, 表型贡献率为35.37%。

图2 大麦籽粒总花色苷含量和千粒重QTL在染色体上的分布
三角形图案表示总花色苷含量QTL, 五角星图案表示千粒重QTL。白色、灰色和黑色图案分别表示在玉溪(2013年)、白邑(2014年)和嵩明(2015年)检测到的QTL。图中右侧标示的是2H和7H部分分子标记对应的物理位置。
Triangle pattern indicates QTL for total anthocyanin content (TAC), pentagram pattern indicates QTL for 1000-kernel weight (TKW). The white patterns, gray patterns and black patterns represent the QTL loci detected in Yuxi (2013), Baiyi (2014), and Songming (2015), respectively. The physical positions corresponding to some molecular markers on 2H and 7H are indicated on the right side of the Figure.
2H Bmag0125−GBM1309和7H EBmatc0016− Bmag0206区间至少2年2点能重复检测到2个总花色苷含量QTL; 2H Scssr03381−scssr07759与7H GBM1297−GBM1303区间3年3点能重复检测到2个千粒重QTL, 相应位点加性作用方向均相同, 说明这些QTL比较可靠。
3 讨论
3.1 大麦总花色苷含量及千粒重的遗传变异
关于不同作物花色苷含量及千粒重的遗传变异已有一些研究报道。孙明茂等[34]分析表明, F5家系糙米总花色苷含量和千粒重变异幅度分别为0~ 5459.34 mg kg–1和11.96~26.24 g, 平均值分别为834.47 mg kg–1和17.75 g, 变异系数分别为191.96%和12.89%, 均呈右偏态、尖顶峰分布。黄莹莹[35]报道, 水稻F3家系籽粒总花色苷含量和千粒重变幅分别为0.15~5.97 μg g–1及19.00~30.16 g, 平均值分别为1.80 μg g–1及24.05 g, 变异系数分别为67.72%及8.93%, 均呈近正态分布。吕毅[22]调查表明, 大麦F2群体千粒重呈连续正态分布。王楠等[36]研究表明不同大麦品种花色苷质量分数在0~182.41 μg g–1之间。胡家坤等[37]分析25个不同颜色地方大麦品种籽粒花色苷质量分数变幅为0~291.16 μg g–1。
本研究结果表明, 玉溪(2013年)、白邑(2014年)及嵩明(2015年) 3个试点大麦RIL群体总花色苷含量变幅分别为0.36~1.38、0.50~1.50和0.50~1.58 mg g–1, 变异系数为25.00%~33.85%; 千粒重变幅分别为31.65~58.00、23.05~60.50和25.00~57.60 g, 变异系数13.19%~17.78%。大麦RIL群体总花色苷含量变异幅度大于王楠等[36]、胡家坤等[37]的报道, 千粒重变幅大于孙明茂等[34]、黄莹莹[35]的结果。表明紫光芒裸二棱/ Schooner构建的RIL群体总花色苷含量与千粒重呈现广泛遗传变异, 适合用于相关基因定位研究。大麦总花色苷含量呈右偏态、尖顶峰分布, 千粒重呈近正态分布, 这与孙明茂等[34]、黄莹莹[35]及吕毅[22]的结果基本一致。
3.2 大麦总花色苷含量和产量构成性状相关性
孙明茂等[34]分析表明水稻F5家系糙米总花色苷含量与千粒重呈极显著负相关(–0.34**)。Zheng等[38]报道紫黑水稻产量(包含产量构成性状)越低, 籽粒花色苷含量越高。常汇琳[39]研究表明RIL群体糙米总花色苷含量与千粒重极显著负相关。大麦产量性状与农艺性状、品质性状之间的关系已有不少报道[40-41], 但大麦总花色苷含量和产量构成性状间的相关性未见报道。本研究表明, 大麦RIL群体总花色苷含量与千粒重呈显著或极显著负相关。说明在一定程度上, 总花色苷含量高的材料, 千粒重可能小一些, 这与前人的研究结果基本一致。因此, 在筛选高花色苷含量材料的同时, 应注意千粒重等产量构成性状的影响。
很多研究报道相同或相邻近的染色体区段常发现同时控制多个相关性状的QTL。Woodward和Thieret[12]发现控制大麦紫色的两个互补基因与二棱及六棱大麦花序基因()密切相关。Hori等[25]、Bauer等[42]、赖勇等[43]在2H上检测到标记Bmag0125与千粒重紧密连锁。李静烨等[9]在2HGBM1121标记附近发现千粒重关联位点。Marquez-Cedillo等[44]在2H染色体发现的千粒重QTL可能是基因多效性或紧密连锁基因表达的结果。本研究在前人已报道的2H染色体上与千粒重关联标记区段也检测到花色苷QTL, 表明花色苷和千粒重之间可能存在较紧密的遗传关系。本研究在2H和7H染色体上发现了控制花色苷和千粒重的QTL, 且大多为贡献率大于10%的主效位点, 但由于标记密度不够, 控制花色苷和千粒重的相关位点间遗传距离较大。故下一步工作需通过加密标记来进一步解析花色苷和千粒重之间负相关的遗传基础。
3.3 QTL一致性分析
目前已经有许多关于大麦花色苷和千粒重QTL定位的研究报道。本研究通过3年3试点研究, 共发现12个大麦籽粒总花色苷含量QTL, 其中2H Bmag0125–GBM1309和7H EBmatc0016–Bmag0206区间至少2年2点能重复检测到总花色苷含量QTL。Jia等[17]在大麦2HL染色体Bmag0125–GBMS244间发现1个控制紫粒颜色的基因位点, 并将其精细定位于InDel标记PQJ1056和HvOs04g47170之间。Shoeva等[18]发现大麦类黄酮物质合成途径调控基因位点位于2H Bmag0140–Bmag0125之间。Zhang等[45]在大麦2HL M1573231–BMac0144i间检测到控制紫色皮大麦主要花色苷成分P3G和C3G的QTL, 且标记BMac0144i与Bmag0125位置相同。本研究在2H Bmag0125–GBM1309区间重复检测到的对总花色苷的贡献率均大于10%, 属于主效 QTL, 与前人所报道的结果基本一致。Zhang等[45]在大麦7HS M1557873–M44011之间也检测到控制紫色皮大麦主要花色苷成分P3G和C3G的QTL。本研究在7H EBmatc0016–Bmag0206区间重复检测到的主效QTL与Zhang等[45]、Himi等[46]发现的位点在同一染色体臂上。相比之下, 本研究在1H染色体检测到的、4H上的2个QTL和6H上的3个QTL, 均属于微效QTL, 在相关染色体臂上未见报道。通过与大麦参考基因组比对, 以上能重复检测到的2个主效QTL区间与对应物理位置有较好的一致性。然而是否有新的基因位点参与花色苷合成仍有待进一步研究。
本研究共检测到8个大麦籽粒千粒重含量QTL, 其中2H Scssr03381–scssr07759及7H GBM1297– GBM1303区间均重复检测到千粒重QTL。关伟伟[47]、Pillen等[48]发现与标记GMS3连锁的千粒重QTL, 与本研究2H Scssr03381–scssr07759区间重复检测到的微效QTL较近, 而2H上的另一个千粒重QTL与Beheshtizadeh等[49]所报道的区间相近。司二静等[50]用GLM关联分析模型在4H染色体GMS89附近发现千粒重关联位点, 与本研究的微效QTL接近。本研究第7H染色体GBM1297− GBM1303区间重复检测到的主效QTL与关伟伟[47]、司二静等[50]发现的QTL位于同一染色体臂。通过与大麦参考基因组比对, 以上能重复检测到的2个QTL区间与对应物理位置有较好的一致性, 且大多为主效QTL。本研究检测到的QTL与已报道的相关QTL间的关系还需要深入探究。
目前, 已有很多研究报道了不同大麦材料花色苷和千粒重性状的相关QTL, 但能利用的不多。其中有很多原因, 如环境条件不稳定、QTL遗传效应小及基因互作等。本研究定位的部分QTL区间都有相关报道, 说明其结果可靠性较大且遗传稳定性较好。此外, 本研究检测到的QTL贡献率大于10%的有10个, 贡献率大于20%的有5个, 最大的可达42.32%。对这些效应值较大的QTL, 后续工作可通过构建近等基因系分离群体及设计分子标记加密进行基因精细定位及克隆并加以利用。
4 结论
利用大麦RIL群体, 共检测到12个总花色苷含量QTL, 分别位于1H、2H、4H、6H和7H染色体; 8个千粒重QTL, 分别位于2H、4H和7H染色体。贡献率大于10%的QTL有10个, 大于20%的有5个, 最大的达42.32%。其中控制总花色苷含量的2个QTL至少在2年2点、千粒重的2个QTL在3年3点可被重复检测到。控制总花色苷含量与千粒重的主效QTL同位于2H和7H染色体。大麦籽粒总花色苷含量和千粒重之间呈显著或极显著负相关。
[1] Zhu F. Anthocyanins in cereals: composition and health effects., 2018, 109: 232–249.
[2] 华为, 朱靖环, 尚毅, 贾巧君, 汪军妹, 杨建明. 有色大麦研究进展. 植物遗传资源学报, 2013, 14: 1020–1024. Hua W, Zhu J H, Shang Y, Jia Q J, Wang J M, Yang J M. Research advances in colored barley., 2013, 14: 1020–1024 (in Chinese with English abstract).
[3] Ma LY, Sun Z H, Zeng Y W, Luo M C, Yang J Z. Molecular mechanism and health role of functional ingredients in blueberry for chronic disease in human beings., 2018, 19: 1–19.
[4] Khlestkina E. The adaptive role of flavonoids: emphasis on cereals., 2013, 41: 185–198.
[5] 杜连起, 李润丰. 黑大麦营养价值及其开发利用. 粮食与油脂, 2001, (2): 40–41. Du L Q, Li R F. Nutritional value and development of black barley., 2001, (2): 40–41 (in Chinese with English abstract).
[6] Martínez M, Motilva M J, de las Hazas M L, Romero M P, Vaculova K, Ludwig I A. Phytochemical composition and β-glucan content of barley genotypes from two different geographic origins for human health food production., 2018, 245: 61–70.
[7] Deng G F, Xu X R, Zhang Y, Li D, Gan R Y, Li H B. Phenolic compounds and bioactivities of pigmented rice., 2013, 53: 296–306.
[8] Cimino F, Ambra R, Canali R, Saija A, Virgili F. Effect of cyanidin-3-O-glucoside on UVB-Induced response in human keratinocytes., 2006, 54: 4041–4047.
[9] 李静烨, 王楠, 陈升位, 吴亚琼, 王茹媛, 沈真辉, 莫凡. 不同大麦品种(或品系) 3个穗部性状的差异分析和QTLs检测. 分子植物育种, 2018, 16: 3973–3979. Li J Y, Wang N, Chen S W, Wu Y Q, Wang R Y, Shen Z H, Mo F. Difference analysis and QTLs detection on three spike traits of different barley varieties (or strains)., 2018, 16: 3973–3979 (in Chinese with English abstract).
[10] Kim M J, Hyun J N, Kim J A, Park J C, Kim M Y, Kim J G, Lee S J, Chun S C, Chung I M. Relationship between phenolic compounds, anthocyanins content and antioxidant activity in colored barley germplasm., 2007, 55: 4802–4809.
[11] Mayler J I, Stanford E H. Color inheritance in barley., 1942, 34: 427–436.
[12] Woodward R W, Thieret J W. A genetic study of complementary genes for purple lemma, palea and pericarp in barley (L.)., 1953, 45: 182–185.
[13] Choo T M, Vigier B, Ho K M, Ceccarelli S, Grando S, Franckowiak J D. Comparison of black, purple and yellow barleys., 2005, 52: 121–126.
[14] Finch R A, Simpson E. New colours and complementary colour genes in barley., 1978, 81: 40–53.
[15] Strygina K V, Börner A, Khlestkina E K. Identification and characterization of regulatory network components for anthocyanin synthesis in barley aleurone., 2017, 17: 109–117.
[16] Lundqvist U, Franckowiak J D, Konishi T. New and revised descriptions of barley genes., 1997, 26: 22–516.
[17] Jia Q J, Zhu J H, Wang J M, Yang J M, Zhang G P. Genetic mapping and molecular marker development for the genecontrolling purple grains in barley., 2016, 208: 215–223.
[18] Shoeva O Y, Mock H P, Kukoeva T V, Börner A, Khlestkina E K. Regulation of the flavonoid biosynthesis pathway genes in purple and black grains of hordeum vulgare., 2016, 11: 1–16.
[19] 杜欢, 张颖, 薛梦瑶, 靖姣姣, 白志英, 李存东. 大麦株高近等基因系的籽粒性状差异及相关分析. 华北农学报, 2015, 30(5): 97–103. Du H, Zhang Y, Xue M Y, Jing J J, Bai Z Y, Li C D. Difference and correlation analysis of grain traits in the near-isogenic line of plant height of barley., 2015, 30(5): 97–103 (in Chinese with English abstract).
[20] 张小燕, 张跃进, 潘高峰. 日本不同棱型大麦种质资源农艺性状的差异. 麦类作物学报, 2006, 26(6): 39−41. Zhang X Y, Zhang Y J, Pan G F. Study on germplasm resource among different rowed barley., 2006, 26(6): 39−41 (in Chinese with English abstract).
[21] 李喜焕, 常文锁, 张彩英, 李保元, 马峙英. 引进春播啤酒大麦品种生育时期及产量性状鉴定与筛选. 河北农业大学学报, 2007, 30(3): 22−25. Li X H, Chang W S, Zhang C Y, Li B Y, Ma Z Y. The evaluation on growing stages and yield traits of seven introduced malting beer barley varieties., 2007, 30(3): 22−25 (in Chinese with English abstract).
[22] 吕毅. 青藏高原一年生野生大麦若干农艺性状的QTL定位. 华中农业大学硕士学位论文, 湖北武汉, 2007. pp 18−25. Lyu Y. QTL Analysis of Some Agronomic Traits in Qing-Tibetan Plateau Annual Wild Barley. MS Thesis of Huazhong Agricultural University, Wuhan, Hubei, China, 2007. pp 18−25 (in Chinese with English abstract).
[23] 王建林, 冯西博, 付刚, 侯维海, 王改花, 大次卓嘎, 钟志明. 青藏高原栽培大麦千粒重空间分布格局及其与环境因子的关系. 生态学报, 2018, 38: 1114−1123. Wang J L, Feng X B, Fu G, Hou W H, Wang G H, Dacizhuoga, Zhong Z M. Relationship between spatial distribution pattern and factors affecting weight per 1000-seeds of cultivated barley in Qinghai-Tibet Plateau., 2018, 38: 1114−1123 (in Chinese with English abstract).
[24] Chen G D, Li H B, Zheng Z, Wei Y M, Zheng Y L, Mclntyre C L, Zhou M X, Liu C J. Characterization of a QTL affecting spike morphology on the long arm of chromosome 3H in barley (L.) based on near isogenic lines and a NIL-derived population., 2012, 125: 1385–1392.
[25] Hori K, Kobayashi T, Shimizu A, Sato K, Takeda K, Kawasaki S. Efficient construction of high-density linkage map and its application to QTL analysis in barley., 2003, 107: 806–813.
[26] Baghizadeh A, Taleei A, Naghavi M. QTL analysis for some agronomic traits in barley (L.)., 2007, 9: 372–374.
[27] Wang J, Sun G, Ren X, Li C, Liu L, Wang Q, Du B, Sun D. QTL underlying some agronomic traits in barley detected by SNP markers., 2016, 17: 1–13.
[28] Ashikari M, Sakakibara H, Lin S, Yamamoto T, Takashi T, Nishimura A, Angeles E R, Qian Q, Kitano H, Matsuoka M. Cytokinin oxidase regulates rice grain production., 2005, 309: 741−745.
[29] 覃鸿妮, 晏萌, 王召辉, 郭莹, 王辉, 孙海燕, 刘志斋, 蔡一林. 玉米籽粒中花色苷和黑色素含量的QTL分析. 作物学报, 2012, 38: 275–284. Qin H N, Yan M, Wang Z H, Guo Y, Wang H, Sun H Y, Liu Z Z, Cai Y L. QTL mapping for anthocyanin and melanin contents in maize kernel., 2012, 38: 275–284 (in Chinese with English abstract).
[30] Hosseinian F S, Li W, Beta T. Measurement of anthocyanins and other phytochemicals in purple wheat., 2008, 109: 916–924.
[31] Zeng Y W, Du J, Yang X M, Pu X Y, Wang L X, Yang J Z, Du L J, Yang T, Yang S M, Sun Z H. Identification of quantitative trait loci for mineral elements in grains and grass powder of barley., 2016, 15(4): 1–13.
[32] Li H H, Ribaut J M, Li Z L, Wang J K. Inclusive composite interval mapping (ICIM) for digenic epistasis of quantitative traits in biparental populations., 2008, 116: 243−260.
[33] 王建康. 数量性状基因的完备区间作图方法. 作物学报, 2009, 35: 239−245. Wang J K. Inclusive composite interval mapping of quantitative trait genes., 2009, 35: 239−245 (in Chinese with English abstract).
[34] 孙明茂, 韩龙植. 粳稻龙锦1号/香软米1578杂交组合F5家系群糙米总花色苷含量变异及相关性分析. 植物遗传资源学报, 2017, 18: 186−192. Sun M M, Han L Z. Variation and correlation analyses of total anthocyanin content of brown rice () in F5lines fromrice cross Longjin 1/Xiangruanmi 1578., 2017, 18: 186−192 (in Chinese with English abstract).
[35] 黄莹莹. 水稻子粒花色苷、矿质元素含量及相关性状的QTL定位. 东北农业大学硕士学位论文, 黑龙江哈尔滨, 2013. pp 1−41. Huang Y Y. QTL Identification Underlying Anthocyanin and Mineral Elements Content Related Traits in Rice. MS Thesis of Northeast Agricultural University, Harbin, Heilongjiang, China, 2013. pp 1−41 (in Chinese with English abstract).
[36] 王楠, 王家曦, 谢文英, 陈升位, 杨映宝, 王乐. 25个地方大麦品种籽粒中花色苷成分及其含量差异. 云南农业大学学报, 2015, 30: 829−835. Wang N, Wang J X, Xie W Y, Chen S W, Yang Y B, Wang L. Difference of the anthocyanin components and their contents of 25 local barley variety grains., 2015, 30: 829−835 (in Chinese with English abstract).
[37] 胡家坤, 谢文英, 陈升位, 王楠, 杨映宝, 王乐. 25个大麦地方品种籽粒总黄酮和花色苷的含量差异. 云南农业大学学报, 2015, 30: 522−527. Hu J K, Xie W Y, Chen S W, Wang N, Yang Y B, Wang L. The content difference of total flavonoids and anthocynin from the grains of 25 local barley varieties., 2015, 30: 522−527 (in Chinese with English abstract).
[38] Zheng G Z. Effects of Blackish Purple Colored Seed Coat on Grain Quality and Yield Associated Characters in Rice (L.). MS Thesis of Seoul University, Seoul, Republic of Korea, 2000. pp 1−41.
[39] 常汇琳. 水稻花色苷和原花色素含量的QTL分析及与产量性状关系的研究. 东北农业大学硕士学位论文, 黑龙江哈尔滨, 2015. pp 1−55. Chang H L. QTL Analysis of Anthocyanin and Proanthocyanidin Content and Research on Relationship between Yield Traits in Rice. MS Thesis of Northeast Agricultural University, Harbin, Heilongjiang, China, 2015. pp 1−55 (in Chinese with English abstract).
[40] 杜欢, 马彤彤, 侯晓梦, 张颖, 白志英, 李存东. 20对大麦株高近等基因系农艺与产量性状差异及相关性分析. 华北农学报, 2016, 31(5): 114−121. Du H, Ma T T, Hou X M, Zhang Y, Bai Z Y, Li C D. Difference and correlation analysis of agronomic and yield characters in twenty pairs near-isogenic line of plant height of barley., 2016, 31(5): 114−121 (in Chinese with English abstract).
[41] 张宇, 司二静, 孟亚雄, 马小乐, 李葆春, 王化俊. 不同来源大麦材料的农艺性状和籽粒蛋白质含量及群体遗传结构分析. 麦类作物学报, 2015, 35: 940−947. Zhang Y, Si E J, Meng Y X, Ma X L, Li B C, Wang H J. Analysis on agronomic traits, grain protein content and population structure of barley germplasm from different geographical origins., 2015, 35: 940−947 (in Chinese with English abstract).
[42] Bauer A M, Hoti F, Korff M V, Pillen K, Léon J, Sillanpä M J. Advanced backcross: QTL analysis in spring barley (ssp.) comparing a REML versus a Bayesian model in multi-environmental field trials., 2009, 119: 105–123.
[43] 赖勇, 贾建磊, 王晋民, 任龙, 吕仲昱, 朱惠琴, 马辉, 杨莉娜, 李宗仁. 外引大麦SSR标记遗传多样性及其与农艺性状的关联分析. 麦类作物学报, 2017, 37: 197–204. Lai Y, Jia J L, Wang J M, Ren L, Lyu Z Y, Zhu H Q, Ma H, Yang L N, Li Z R. Analysis of genetic diversity and association with agronomic traits in barley (L.) introduced from abroad using SSR markers., 2017, 37: 197–204 (in Chinese with English abstract).
[44] Marquez-Cedillo L A, Hayes P M, Kleinhofs A, Legge W G, Jones B L, Rossnagel B G, Sato K, Ullrich S E, Wesenberg D M. QTL analysis of agronomic traits in barley based on the doubled-haploid progeny of two elite North American varieties representing different germplasm groups., 2001, 103: 625–637.
[45] Zhang X W, Jiang Q T, Wei Y M, Liu C. Inheritance analysis and mapping of quantitative trait loci (QTL) controlling individual anthocyanin compounds in purple barley (L.) grains., 2017, 12(8): e0183704.
[46] Himi E, Taketa S. Isolation of candidate genes for the barleyand wheatgenes controlling anthocyanin pigmentation in different vegetative tissues., 2015, 290: 1287–1298.
[47] 关伟伟. 大麦主要农艺性状的QTL定位及过氧化物酶、酯酶和细胞色素氧化酶的遗传分析. 华中农业大学硕士学位论文, 湖北武汉, 2011. pp 25–31. Guan W W. QTL Analysis of Main Agronomic Traits and Inheritance of Isozymes Peroxidase, Esterase and Cytochrome Oxidase in Barley. MS Thesis of Huazhong Agricultural University, Wuhan, Hubei, China, 2011. pp 25–31 (in Chinese with English abstract).
[48] Pillen K, Zacharias A, Léon J. Advanced backcross QTL analysis in barley (L.)., 2003, 107: 340–352.
[49] Beheshtizadeh H, Fakheri B A, Aghnoum R, Mahdinezhad N, Pourdad S S, Masoudi B. QTL mapping of grain yield and its components under normal and drought stress conditions in barley (L.)., 2018, 78: 69–80.
[50] 司二静, 张宇, 汪军成, 孟亚雄, 李葆春, 马小乐, 尚勋武, 王化俊. 大麦农艺性状与SSR标记的关联分析. 作物学报, 2015, 41: 1064–1072.Si E J, Zhang Y, Wang J C, Meng Y X, Li B C, Ma X L, Shang X W, Wang H J. Association analysis between SSR marker and agronomic traits in barley., 2015, 41: 1064–1072 (in Chinese with English abstract).
QTL mapping for total grain anthocyanin content and 1000-kernel weight in barley recombinant inbred lines population
YANG Xiao-Meng1,2,**, LI Xia1,2,**, PU Xiao-Ying1,2, DU Juan1,2, Muhammad Kazim Ali3, YANG Jia-Zhen1,2, ZENG Ya-Wen1,2,*, and YANG Tao1,2
1Biotechnology and Germplasm Resources Institute, Yunnan Academy of Agricultural Sciences, Kunming 650205, Yunnan, China;2Agricultural Biotechnology Key Laboratory of Yunnan Province, Kunming 650205, Yunnan, China;3Department of Biotechnology, Fedural Urdu University of Arts, Science and Technology, Karachi 75300, Pakistan
In this study, 193 recombinant inbred lines (RILs) derived from a cross between Ziguangmangluoerleng (purple barley, endemic to Yunnan, China) and Schooner (yellow barley introduced from Australia), grown at three sites of Yunnan province of China for three consecutive years (2013−2015), were used to determine total grain anthocyanin content, 1000-kernel weight and their correlations, and mapped QTLs. There was a significant negative correlation between total grain anthocyanin content and 1000-kernel weight. Twelve QTLs for total grain anthocyanin content were located on chromosomes 1H, 2H, 4H, 6H, and 7H which showed 5.06% to 23.86% of phenotypic variation. Eight QTLs for 1000-kernel weight were located on chromosomes 2H, 4H, and 7H, which explained phenotypic variation from 4.67% to 42.32%. Ten QTLs had phenotypic variation ≥10% and five QTLs ≥ 20%, while the highest phenotypic variation was 42.32%. In addition, two QTLs for total grain anthocyanin content were repeatedly detected at two sites for two years, and located on 2HBmag0125 to GBM1309and 7HEBmatc0016 to Bmag0206 interval, respectively, which accounted for 13.66% to 17.76%, and 13.07% to 16.43% of the phenotypic variation, respectively. Two QTLs for 1000-kernel weight were repeatedly detected at three sites for three years and located on 2H scssr03381 to scssr07759and 7H GBM1297 to GBM1303interval, which showed 4.67% to 14.55% and 34.51% to 42.32% of phenotypic variations, respectively, and their contribution rates of additive effects were consistent. The common major QTLs for two agronomic attributes were mainly distributed on chromosomes 2H and 7H. These results provide a basis for further fine mapping, cloning and marker-assisted breeding of beneficial genes related to total grain anthocyanin content and 1000-kernel weight.
barley RIL; total anthocyanin content; 1000-kernel weight; correlation; QTL mapping
2019-03-18;
2019-08-09;
2019-09-18.
10.3724/SP.J.1006.2020.91024
曾亚文, E-mail: zengyw1967@126.com
**同等贡献(Contributed equally to this work)
杨晓梦, E-mail: yxm89ccf@126.com
本研究由云南省应用基础研究计划项目(2017FD021)和国家现代农业产业技术体系建设专项(CARS-05-01A-04)资助。
This study was supported by Yunnan Applied Basic Research Projects of China (2017FD021) and China Agriculture Research System (CARS-05-01A-04).
URL:http://kns.cnki.net/kcms/detail/11.1809.S.20190918.0908.002.html
猜你喜欢
作文小学高年级(2022年5期)2022-06-16 06:22:50
环境卫生工程(2021年4期)2021-10-13 06:52:26
疯狂英语·新读写(2021年2期)2021-02-25 08:58:46
课堂内外(高中版)(2021年7期)2021-01-17 07:30:28
军事运筹与系统工程(2020年2期)2020-11-16 01:11:04
军事运筹与系统工程(2018年3期)2018-03-26 06:33:02
中学生数理化·七年级数学人教版(2016年2期)2016-05-30 21:20:57
华人时刊(2016年19期)2016-04-05 07:56:10
中亚信息(2016年10期)2016-02-13 02:32:45
VOGUE服饰与美容(2015年6期)2015-05-30 10:48:04
