
水稻矮杆小粒突变体dsg1 的表型鉴定及粒形基因精细定位
2020-12-09 07:00:32吕召坤玉一岚李兰英张德春
农业现代化研究 2020年6期
吕召坤 ,玉一岚 ,李兰英,张德春
(1.中国科学院亚热带农业生态研究所亚热带农业生态过程重点实验室,湖南 长沙 410125;2.中国科学院大学,北京 100049;3.三峡大学生物技术研究中心,湖北 宜昌 443002)
稻米的产量和品质对于确保国民粮食安全和促进农业可持续发展具有重要意义。水稻(Oryza sativa)是世界主要粮食作物之一,全世界有接近一半的人口以稻米为主食。虽然水稻半矮杆基因的发掘和应用以及杂种优势的利用使得水稻产量取得了巨大的飞跃[1],然而,面对世界人口的持续增长(预计2050 年将达到90 亿)[2],水稻育种家和植物遗传学家需要进一步提高水稻的产量。
水稻产量是一种多性状共同决定的复杂农艺性状,主要受有效穗数、每穗粒数和千粒重三因素制约[3]。有效穗数在很大程度上取决于单株分枝数、分蘖角度和株高;每穗粒数主要取决于穗部结构,如穗长、每穗分枝数、籽粒密度和育性;而千粒重直接受到籽粒的形状和大小的影响[4]。此外,粒形也是评价稻米品质的重要指标之一,印度、越南和美国偏爱细长型稻米,而中国东北、韩国和日本更喜欢短圆型稻米[5]。因此,剖析稻米粒形的遗传基础对水稻产量和品质具有重要意义。
粒形可以划分为粒长、粒宽、粒厚三个不同的影响因子,且三者都是由多基因控制的数量性状[6]。揭示水稻粒长、粒宽和粒厚的分子遗传机理,对于全面解析控制粒形性状的分子调控网络和挖掘可利用的优异育种基因资源具有重要意义。因此,寻找并克隆控制相应性状的数量性状位点(Quantitative trait loci, QTL)及剖析相关信号转导网络是研究粒形的重要途径。目前,在水稻12 条染色体上检测到与粒形相关的QTL 已经超过600 个,其中93 个已被成功克隆并进行了功能分析[7-8]。它们通过不同的信号途径调控水稻粒形的发育,主要包括泛素—蛋白酶体途径、G 蛋白信号途径、转录调控途径、MAPK 信号途径以及植物激素调控途径。GW2 和GW5 通过泛素—蛋白酶体途径调控水稻籽粒大小。GW2编码一个E3 泛素连接酶,通过降解促进细胞分裂的蛋白酶负调控水稻颖壳大小,进而影响籽粒大小[9]。GW5编码的蛋白质与多聚泛素有相互作用,通过泛素—蛋白酶体途径调控籽粒颖壳的细胞分裂数目对籽粒大小进行调控[10-11]。GS3和DEP1通过G 蛋白信号途径参与稻米粒形的调控。GS3编码一个由5 个外显子组成的Gγ 亚基,它的功能缺失会导致籽粒明显变小;而DEP1是一种功能获得性突变基因,功能获得性突变dep1能够促进细胞分裂,并使穗粒数增加,促进水稻增产[12-14]。GLW7、GW8、LGY3等在转录调控因子途径中参与水稻粒形的调控,GW8和GLW7的编码蛋白都是调控粒形的正向调控因子,其高表达水平能够促进颖壳细胞分裂,导致籽粒变大,LGY3编码的转录因子在第7 内含子和第8 外显子的连接区发生突变,产生一个新的可变剪切位点,导致籽粒变长[15-17]。SMG1编码一个丝裂原活化蛋白激酶MAPK4,通过MAPK 信号途径调控籽粒形状;而GS2/GL2参与了植物激素油菜素内酯调控途径[18-21]。虽然这些基因部分解析了调控稻米粒形的分子机理,但这对于了解水稻粒形遗传网络还远远不够。因此,发现新的调控粒形的QTL 位点及其调控机制更有利于全面阐述整个遗传网络,为今后水稻产量和稻米品质的提高提供遗传基础。
在本研究中,矮杆小粒突变体dsg1(dwarf and short grain)是由TB309 经甲基磺酸乙酯(thymethylsulfone,EMS,体积比为0.8%)溶液诱变后筛选得到的。通过对dsg1进行表型和农艺性状考察,发现粒宽、粒厚和千粒重显著降低,并且穗长和地上部节间距也显著缩短。对候选基因进行精细定位,将突变基因dsg1定位于水稻第4 号染色体,位于分子标记ID2798 与ID2803 之间的52.28 kb 区间内,并利用Gramene 数据库在该定位区间找到11 个候选基因,为dsg1基因的克隆和功能研究奠定了基础。
1 材料与方法
1.1 试验材料
dsg1突变体由TB309 经EMS 诱变获得,经连续多年自交已稳定遗传。dsg1与9311 杂交后构建F2分离群体进行基因定位。
1.2 试验方法
1.2.1 主要农艺性状考察 将实验材料TB309、突变体dsg1、9311 和F2群体种植于湖南宁乡关山和海南三亚水稻田,单株种植,行距为26 cm,株距为16 cm,大田常规管理。在水稻成熟期进行株高测量,分别测100 株;并测定茎秆的节间长度,测10 个重复。种子成熟后,单株收取各家系植株,用恒温干燥箱60 ℃烘烤直至籽粒重量恒重为止。利用游标卡尺对实验材料TB309、突变体dsg1、9311 和F2群体的成熟种子进行粒长、粒宽和粒厚等指标的测定,每个性状测10 个重复。将TB309 和dsg1植株的整个穗子取回,测量其穗长、千粒重、结实率等性状,每个性状测10 个重复[22]。
1.2.2 遗传分析 以籼稻9311 和突变体dsg1为亲本,经过杂交后自交得到由68 个单株组成的F2群体,对F2群体的粒形分离情况进行统计分析,并进行卡方测验。
1.2.3 基因定位 根据实验室已有的存在于水稻12条染色体上的Indel 引物,筛选在dsg1和9311 两亲本间存在多态性的引物。从dsg1/9311 的F2分离群体挑选10 株极端矮化小粒的单株和10 株极端长粒表型的单株,采用极端个体法进行连锁分析。利用均匀分布于12 条染色体的171 对有多态性的分子标记扩增极端个体的DNA,用3%的琼脂凝胶进行检测,并做好数据记录[23]。使用作图软件MAPMAKER(EXP3.0b)构建突变基因所在范围的分子标记连锁图谱,将突变基因定位在一条染色体上的两个标记之间,完成初定位。初定位后,扩大F2群体,同时使用16 个具有多态性的标记进行候选基因的精细定位,并绘制突变基因所在位置的遗传图谱。精细定位所用引物序列见表1。
1.2.4 BR 激素敏感性实验 将去壳的饱满水稻种子经75%乙醇和5%次氯酸钠消毒处理后,在无菌条件下分别播种至含有表油菜素内酯(EpiBL)和不含EpiBL 的1/2 MS 固体培养基,生长8 d 后,观察根长生长情况,记录数据[24]。
1.2.5 数据分析 所有数据的平均值计算、显著性差异分析、卡平方测验均用Microsoft Excel 软件进行。根据x2的数值和自由度得到概率值,以P=0.05 为适合性测验符合的下限。
2 结果与分析
2.1 突变体dsg1 的表型分析
在自然生长条件下,对dsg1与TB309 进行表型考察。利用t测验对表型数据进行分析,分析结果显示:与野生型相比,突变体dsg1表现出明显的矮杆表型(表2,图1a),其株高约为TB309 的79.00%,倒一节、倒二节、倒三节和穗长的长度显著低于TB309(表2,图1b、图1d)。与TB309 相比,dsg1的籽粒出现明显的小粒表型,粒长、粒宽、粒厚分别减小15.53%、7.26%和10.45%,千粒重降低了34.14%(表2,图1c)。另外,与野生型相比,dsg1分蘖数也显著减少,单株总产量显著降低。

表1 本研究使用的引物Table 1 The primers used for this study
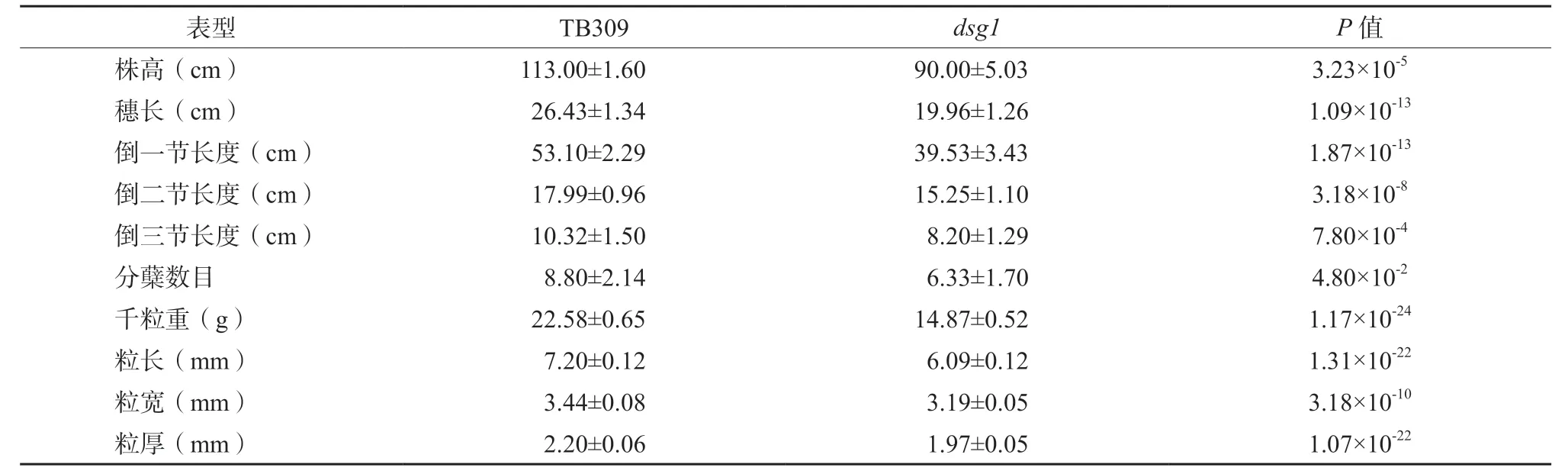
表2 野生型TB309 与突变体dsg1 的表型鉴定Table 2 Comparison of agronomic trait of the wild type TB309 and the dsg1 plants
2.2 油菜素内酯激素敏感性分析
由于突变体不仅在粒形性状上发生改变,在植株高度、分蘖数目、节间长度和稻穗长度等方面也表现出显著性变化,这与先前报道的油菜素内酯的合成或代谢缺陷表型相似[25-26],因此,我们猜测dsg1突变体可能与BR 激素有关。为了验证这个猜测,我们对野生型TB309 和突变体dsg1进行了BR激素响应的敏感性实验。我们通过外施EpiBL 的方式检测DSG1是否参与BR 响应。在施加EpiBL 培养后,野生型TB309 的主根长度明显缩短,说明外源施加EpiBL 抑制了根部的伸长;然而突变体dsg1的主根长度并没有显著性变化(图2a, 图2b)。由此表明,dsg1并不是一个BR 合成缺陷突变体,而有可能是BR 信号转导过程中出现缺陷导致了突变。

图1 野生型TB309 与突变体dsg1 的表型鉴定Fig. 1 Phenotypic identification of the wild type TB309 and the dsg1 mutant
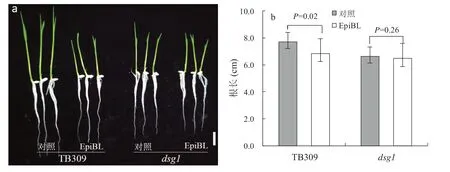
图2 突变体dsg1 对BR 响应敏感性Fig. 2 Mutant dsg1 as a BR-insensitive mutant
2.3 遗传分析
为了克隆该突变体的基因,把粒长作为目标性状对DSG1 进行遗传效应分析和精细定位研究。我们将dsg1与9311 进行杂交,得到F1代植株,其表型与9311 表型相似。将F1代自交获得F2群体,在F2群体中出现9311 粒长表型和突变体小粒表型分离,说明该突变表型是由隐性基因控制的。统计F2群体中粒长表型的单株数量,发现粒长表型频率呈现双峰分布(图3),表明存在主效QTL。在F2群体中,长粒和短粒表型数目分别为42 和26,。对粒长表型进行x2测验,两种表型在F2群体中的分离比符合3 ∶1(x2=0.16,x20.05=3.84,df=1),符合孟德尔遗传规律,表明该dsg1的粒长性状受单基因控制。
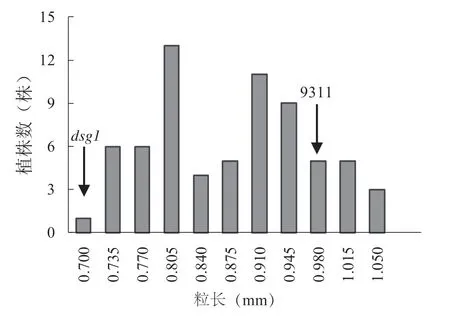
图3 F2 群体粒长性状频率分布Fig. 3 Frequency distribution of grain length in a F2 population
2.4 基因定位
将dsg1与9311 进行杂交,在F2分离群体中分别选取10 株极端小粒表型单株和极端长粒表型的单株,利用均匀分布于12 条染色体的Indel 分子标记进行连锁分析,将DSG1基因初定位于第4 染色体标记ID2610 和ID3004 之间。为了进一步对候选QTL 进行精细定位,在初定位区间内设计14 对新的多态性分子标记,利用交换单株缩小定位区间,最终将突变基因定位在分子标记ID2798 与ID2803的区间内,物理距离约为52.28 kb,其间包含11 个候选基因(图4)。
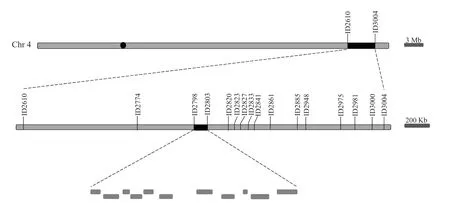
图4 水稻突变体dsg1 的基因定位Fig. 4 Fine mapping of rice mutant dsg1 gene
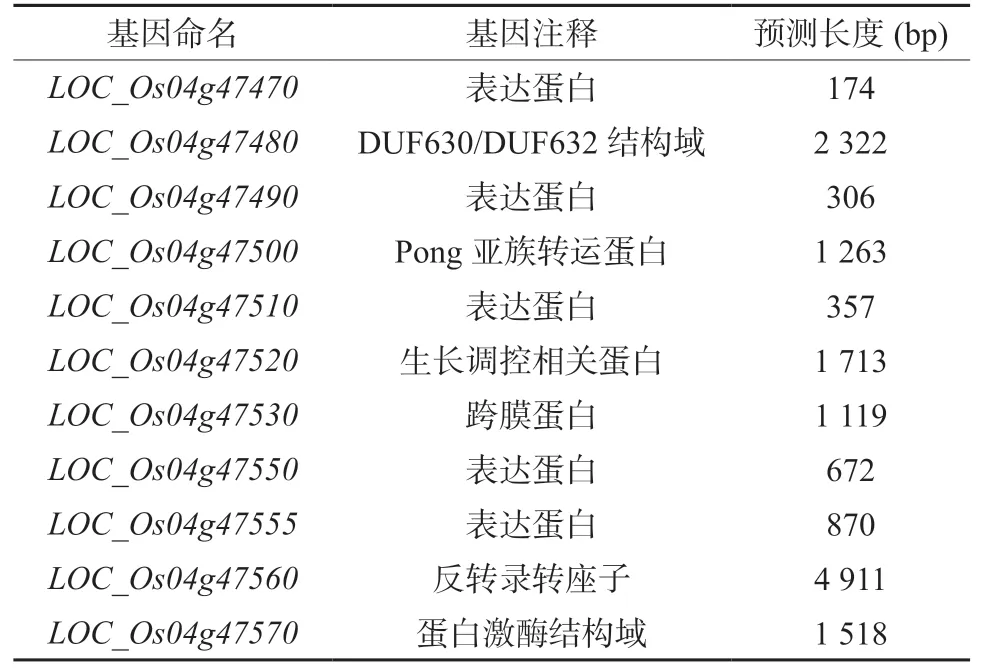
表3 定位区间内的候选基因注释Table 3 Annotation of DSG1 candidate genes in the location interval
2.5 候选基因的预测及分析
根据水稻基因组数据库(http://rice.plantbiology.msu.edu)提供的基因注释信息,在52.28 kb 候选区间内存在11 个注释基因(表3)。其中3 个编码表达蛋白,2 个功能未知蛋白,其余分别编码转座蛋白、逆转录转座子蛋白、跨膜蛋白、含有DUF630/DUF632 域的蛋白、含有激酶结构域的蛋白和生长调控相关蛋白。通过Gramene 网站进行生物信息学分析后发现,该区间内存在3 个可能与水稻株高、穗型和粒性发育相关的基因。LOC_Os04g47480预测结果显示,其编码蛋白可能含有DUF630/DUF632 结构域。未知功能结构域蛋白家族(domains of unknown function protein families, DUFs)是一类进化保守且有功能的蛋白家族,其不同成员参与不同的生长发育过程及胁迫响应[27]。含有DUF630/DUF632 结构域的蛋白参与了叶片形态建成,进而导致了稻米籽粒大小的变化。LOC_Os04g47570的表达蛋白含有激酶结构域,其同源基因LOC_Os03g40400 的表达产物参与了GS3 和qGL3 调控稻米籽粒大小的信号转导过程,其表达水平上调了1.5 倍,该基因的表达可能会影响稻米籽粒的大小[28]。LOC_Os04g47510 在水稻中有一个同源基因LOC_Os07g27670,该基因的编码蛋白是WRKY 家族中的一员。WRKY 家族蛋白是一种转录因子,广泛参与并调控植物生长发育、形态建成和各种生物或非生物胁迫反应[29]。WRKY 蛋白能够与生长素(IAA)、赤霉素(GA)、脱落酸(ABA)以及油菜素内酯(BR)等多种植物激素相互作用调控植物生长发育[30-31]。在拟南芥中,WRKY46、WRKY54 和WRKY70 能促进与BR 相关的生长基因表达并抑制抗旱基因的表达,调节拟南芥生长[32]。在苹果中,转录因子MdWRKY9 直接抑制油菜素类固醇限制合成酶MdDWF4 的转录,减少BR 产生,调控植株高度[33]。在水稻中,OsWRKY53 能够正向调控BR信号进而调控水稻叶片角度及籽粒大小,其过表达植株的叶角增加,稻米籽粒变大;而OsWRKY78的RNAi 植株可能由于细胞长度的缩减而表现出半矮化和小籽粒的表型[34-35]。因此,这3 个基因都有可能是DSG1的候选基因。
3 讨论
水稻粒形是重要的农艺性状之一,对水稻的产量和品质起到重要作用,是多基因、多途径调控的复杂遗传性状,也是分子设计育种的主要性状之一[36]。分子设计育种是在解析作物重要农艺性状遗传机理的基础上,通过品种设计对复杂性状进行定向改良,达到聚合优异性状的目标,实现作物复杂性状的精准改良[37]。实现精准设计育种的难点在于调控目标性状的遗传网络非常复杂,所以,分子设计育种的前提和要求是对目标性状的分子机理和遗传基础有较为深刻的理解。虽然在水稻中已克隆93个与粒型相关的QTL,然而,可应用于分子设计育种的控制稻米籽粒的基因非常有限。利用Ghd7不同等位基因之间地理分布不同的特点,Ghd7在不同生态区的水稻生育期改良中得到了高效利用,获得了生育期改良的新品系[38]。Wang 等将GW8和GW7导入我国高产水稻品种,在不减产的基础上提升稻米品质[39]。还有研究表明,将GS3、DPE1、qSW5、SD1、Ghd7、Ghd8和Wx等基因聚合获得的改良品种与超级杂交稻“两优培九”相比,该新品种表现出更高的产量潜力和更好的籽粒品质[40]。所以,随着分子设计育种构想的发展,需要挖掘更多的粒形相关基因。在本研究中,我们利用EMS 诱变粳稻品种TB309,筛选得到遗传性状稳定的矮杆小粒突变体dsg1,并将其定位于第4 染色体分子标记ID2798 与ID2803 之间52.28 kb 的区间内,以期克隆到相关基因,以便应用于分子设计育种。
此外,我们发现dsg1突变体不仅粒形性状发生显著性变化,而且株高、穗长、分蘖数目和节间长度等其他农艺性状也呈现显著性减小,说明该基因能够控制多种不同的表型,这可能与“一因多效”有关。一因多效是自然界中广泛存在的现象,其概念最早是由孟德尔所提出的,用来描述一个单独的基因或位点影响两个或两个以上不同性状的现象[41]。例如,GS3编码的Gγ 亚基,可通过G 蛋白信号途径调控稻米粒长和粒重,同时对粒宽和粒厚也具有微效作用[15];DEP1不仅能够调控粒长和千粒重,还对穗长、每穗实粒数、耐旱性等其他性状有影响[42];WTG1同时控制每穗粒数和分蘖数目2 种不同的性状[43-44];Ghd7同时调控水稻抽穗期、每穗粒数和株高3 种性状,猜测可能是抽穗期延迟增加了穗部和秸秆等器官的发育进程从而使产量增加和植株变高[39]。同样,DSG1可能能够影响不同的生化代谢途径,形成复杂的调控网络,进而影响多种农艺性状。本研究完成了对该基因的精细定位,并通过基因注释信息分析和激素敏感性实验得到3个可能是DSG1的候选基因,为进一步克隆该基因并剖析其分子机制奠定了基础,同时对于利用分子设计育种的方法改善稻米产量和品质也具有重要意义。
4 结论
1)本研究利用EMS 技术诱变粳稻品种TB309获得了一个矮杆小粒的突变体dwarf and short grain(dsg1)。与野生型相比,突变体dsg1的株高、粒长、粒宽、粒厚和千粒重显著降低,并且突变体的穗长和节间距也显著缩短。
2)通过植物激素敏感性实验证实dsg1有可能是BR 信号转导过程中出现缺陷导致了突变,而不是BR 合成缺陷突变体。在施加EpiBL 培养后,野生型TB309 的主根长度明显缩短,但是突变体dsg1的主根长度并没有显著性变化。
3)对突变体dsg1与9311 杂交获得的F2群体进行表型考察和遗传分析,F2代出现表型分离,且符合孟德尔遗传规律,说明该粒长突变基因是由隐性单基因控制。
4)通过精细定位,将突变基因定位于水稻第4染色体分子标记ID2798 与ID2803 之间52.28 kb 的区间内,并利用Gramene 数据库对定位区间进行基因预测,发现该区间存在11 个基因,这为该突变基因的克隆和功能分析奠定了基础。
猜你喜欢
广东农业科学(2021年3期)2021-04-23 04:34:48
现代园艺(2017年21期)2018-01-03 06:41:32
安徽医科大学学报(2016年12期)2017-01-15 14:21:44
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
广东农业科学(2016年11期)2016-03-29 07:52:11
华北农学报(2016年1期)2016-03-18 07:58:57
中国康复理论与实践(2015年10期)2015-12-24 05:42:44
医学研究杂志(2015年5期)2015-06-10 06:43:26
农业科技与装备(2015年5期)2015-05-30 10:48:04
山东医药(2015年40期)2015-02-28 14:28:45
