
机采棉化学脱叶伴随着剧烈的乙烯及细胞分裂素信号响应
2020-12-08 03:59高瑜徐娇张冰孙伟男杨细燕
棉花学报 2020年6期
高瑜,徐娇,张冰,孙伟男,杨细燕
(华中农业大学作物遗传改良国家重点实验室,武汉430070)
棉花是我国重要的经济作物。 棉花全程机械化是稳定棉花种植面积及其产业链,实现“快乐植棉”的根本出路[1]。 机采机收是棉花全程机械化的关键环节。 自21 世纪初采棉机在我国推广以来,我国机采技术不断发展,机采棉花比例逐步上升,相关政策法规体系日趋完善,综合效益逐步呈现。 2019 年新疆机采棉面积达到77 万hm2,北疆90%棉田实现机采,黄河流域和长江流域棉区也在积极示范推广。
化学脱叶是机采前必需的技术措施,能有效减少机采棉杂质含量。 化学脱叶剂作用原理主要是通过调控植物内源激素平衡,诱导叶片形成离层进而脱落[2]。脱叶剂可以分为触杀型和内吸型。触杀型脱叶剂通过化学作用伤害植物细胞组织,刺激内源乙烯的生成, 达到脱叶催熟的效果,如脱叶膦(Butiphos)、百草枯(Paraquat)等;内吸型脱叶剂不伤害植物组织,通过喷施直接释放乙烯的化合物达到脱叶催熟效果, 如乙烯利(Ethephon)、噻唑隆(Thidiazuron)等[3]。 生产上常用的脱叶剂为内吸型脱叶剂,主要由乙烯利、噻唑隆及相关助剂组成。 噻唑隆具有细胞分裂素的效果,能促进细胞急剧分裂,其诱发的乙烯释放与化学伤害、衰老无关,有较好的脱叶效果[4]。
脱落是1 种普遍的自然现象,正常的脱落与成熟衰老有关, 具有高度的精确性和程序性[5-7]。植物激素对脱落具有重要的调控作用,乙烯和生长素是研究较多的2 类激素。Addicott[8]认为植物器官的脱落是由近轴端和远轴端生长素的相对浓度,即“生长素梯度”决定的。 生长素可能通过阻断离层细胞对乙烯信号的响应而抑制脱落[9-10]。乙烯依赖性信号转导途径是目前认为的主要脱落调控途径。 乙烯的含量直接影响离层的形成和器官的脱落。 这一过程主要有3 种基因的参与:(1) 乙烯信号转导途径中的乙烯受体基因,如ETR1 和EIN4 等;(2)乙烯响应转录因子基因,如ERF, 可以对乙烯做出反应, 诱导离层的形态变化;(3)器官脱落相关基因,如编码纤维素酶的基因等,能响应脱落信号,启动器官脱落[5]。 研究表明棉花叶片脱落期间内源乙烯含量有动态变化[4]。对棉花中脱叶剂敏感品种新陆早50 和不敏感品种新陆早33 研究发现, 脱叶剂处理后2 个品种离层组织乙烯含量均显著升高,且在脱叶剂敏感品种中极显著升高,并伴随着乙烯合成及信号转导途径相关基因的差异表达[11]。 超表达MADS 转录因子基因FYF 通过调控乙烯信号传导延迟脱落[12]。 脱落酸促进器官脱落,主要是通过促进乙烯合成发挥作用[13]。 细胞分裂素也参与器官脱落的调控。 对脱叶剂处理后不同棉花材料的表达谱研究也表明,多个细胞分裂素氧化酶基因在脱叶剂敏感型品种中提前差异表达,并伴随着细胞分裂素含量的降低,从而加快了敏感品种新陆早50 叶片脱落;进一步研究表明,干涉细胞分裂素氧化酶基因GhCKX3-1(Gh_D13G2062)表达,转基因棉花对脱叶剂的敏感性降低[11]; 分析干涉GhCKX3 表达植株中乙烯含量及信号转导途径相关基因的表达,发现细胞分裂素和乙烯在调控棉花对脱叶剂的响应中存在互作[11]。
脱落的发生还受到其他信号的调控,如IDA(Inflorescence deficient in abscission)小肽介导的脱落途径[14]。 脱落的激活受富含亮氨酸重复序列(Leacine-rich repeat, LRR)的受体激酶HAESA(HAE)和HAESA-like 2(HSL2)调控,HAE/HSL2与SERK1/2/3/4 协同作用[14-15],激活下游由丝裂原活化蛋白激酶4/5(Mitogen-activated protein kinase (MAPK) kinase 4/5, MKK4/5)和MPK3/6 组成的MAPK 级联[16],进而调控转录因子KNAT1、KNAT2/6 和AGL15[17]。 转录因子也参与调控脱落过程,番茄中通过图位克隆得到1 个MADS-box家族的编码基因JOINTLESS, 在jointless 突变体中花柄和果柄不能形成离层[18]。 DNA结合蛋白基因AtDOF4.7 也参与调控脱落过程[19]。 活性氧(Reactive oxygen species,ROS) 信号也参与器官脱落的调控,ROS 清除蛋白超氧化物歧化酶(Superoxide dismutase,SOD)基因和过氧化氢酶基因CAT1 的过表达也会延迟细胞分离[20]。 脱落过程中,一些重要的水解酶基因上调表达,如β-1,4-葡聚糖酶、木葡聚糖内转糖苷酶等[21]。最新研究发现,磺肽素(Phytosulfokine,PSK)参与调控干旱胁迫下番茄落花落果。 过表达肌醇六磷酸酶基因SlPhyt2 促进干旱胁迫下转基因番茄的花脱落,而SlPhyt2 基因沉默植株坐果率比过表达植株显著提高; 进一步研究发现, 植物PSK 的前体是SlPhyt2 的特异性底物[22]。
与其他植物相似,棉花叶片脱落是植株内源激素变化及互作平衡导致叶柄离层形成的结果,常常由衰老引起。 化学脱叶是使用脱叶剂使代谢旺盛的功能叶叶柄处形成离层而被动脱落。 化学脱叶效果受到各种环境因素(温度、湿度等)以及非环境因素(品种、种植方式、脱叶剂种类等)的影响[2,23-24]。 在影响棉花脱叶效果的诸多因素中,品种对脱叶剂的响应是最核心的。 现有研究对脱叶剂的组配及施用技术及生理机制研究较多,而对棉花品种响应化学脱叶机理还没有系统的研究报道[3,11,25-27]。 本研究广泛筛选棉花种质资源获得脱叶剂敏感和不敏感的品种,并检测前期表达谱筛选中差异表达的乙烯相关、细胞分裂素相关及水解酶相关基因在脱叶剂敏感性不同品种中的表达情况,为棉花机采化控技术研发提供理论支持。
1 材料与方法
1.1 试验材料
1.1.1 棉花材料。 200 份棉花种质资源来自于实验室收集的自然群体[28],包括来自于湖北17 份、湖南2 份、四川5 份、安徽2 份、北疆16 份、南疆13 份、甘肃1 份、云南1 份、贵州2 份、河北13份、河南27 份、浙江2 份、江苏30 份、江西1 份、辽宁10 份、山东18 份、山西19 份、陕西7 份、苏联3 份、美国11 份。
1.1.2 脱叶剂配制及用量。脱叶剂(TE)配制参照Xu 等[11],配方为400 mg·L-1噻唑隆+2 mL·L-140%(质量分数)乙烯利+2 mL·L-1脱叶剂助剂,每公顷用量600 L。
1.2 试验方法
1.2.1 温室内脱叶剂处理及材料筛选。 200 份棉花种质资源于2017 年4 月种植于华中农业大学表型组平台温室,试验设置3 个重复。 每盆1 株,互不干扰,分行排列,每行间隔50 cm。 对所有植株进行常规管理。 在初花期调查每个单株总真叶数目(NTL)后,喷施脱叶剂,在处理后第4 天统计植株脱叶后剩余真叶数目 (NLL)。 使用Microsoft Excel 2016 软件对得到的真叶数目、 剩余真叶数目进行统计分析,分别计算各单株的脱叶率rD。rD(%)=(NTL-NLL)/NTL×100。根据脱叶处理后植株表型及脱叶率,筛选得到对脱叶剂处理敏感和不敏感的棉花品种。
1.2.2 脱叶剂敏感和不敏感材料的大田种植处理、脱叶率统计及取样。 分别挑选粒大饱满的脱叶剂敏感和不敏感的棉花种子用硫酸脱绒后洗净晒干保存。 于2018 年4 月中旬种植于华中农业大学棉花试验田,采用营养钵育苗,待2 片子叶平展(5 月上旬)进行大田移栽,株距40 cm,行距50 cm,每个材料种4 行,设3 个重复。移栽后定期进行常规田间管理,按时浇水、整枝、打药等。
10 月上旬吐絮期时, 挑选连续7 d 晴天、气温20~25 ℃的下午, 对大田的棉花材料进行处理,2 行喷施脱叶剂,2 行喷施蒸馏水作为对照。在处理前及处理后第7 天统计棉株叶片数目,根据上述公式计算各单株第7 天脱叶率。 并在处理后第1 天和第3 天用刀片小心选取适量的植株离层样品, 放入2.0 mL 的离心管中, 用液氮速冻,取样后保存在-80 ℃冰箱中备用。
1.2.3 目标基因的选择。 基于实验室前期的研究[11]选择目标基因。 (1)细胞分裂素相关基因:细胞分裂素氧化酶基因GhCKX3-1(Gh_D13G2062)、GhCKX3-2(Gh_A06G0643)、GhCKX3-3(Gh_D07G2372)、GhCKX7(Gh_A05G0290)和细胞分裂素响应调节因子基因GhCTKRR2 (Gh_A01G0218)、GhCTKRR9(Gh_A05G3475)。(2)乙烯相关基因:乙烯合成相关基因GhACCO3(Gh_D07G0841)、GhACCO(Gh_D11G1565),乙 烯 受体 蛋 白 基 因GhETR1(Gh_D11G1181)、GhEIN3(Gh_D07G0364) 和乙烯 信 号 传 导 相 关 基 因GhERF113-1(Gh_03G1349)、GhERF113-2(Gh_A04G0869)、GhERF61(Gh_D08G1225)、GhERF92(Gh_A08G1686)、GhRAP23(Gh_A09G2077)。 (3)水解酶相关基因:GhBXL1(Gh_A10G1516,编码β-D-木糖苷酶)、GhCHI1(Gh_D01G1683,编码内切几丁质酶)、GhGUN1(Gh_D03G0779,编码内切葡聚糖酶)、GhRNS1(Gh_D06G1912,编码核糖核酸酶)、GhE13B-1(Gh_A09G0635,编码β- 葡萄糖苷酶)和GhE13B-2(Gh_A09G1126)。
1.2.4 基因表达量检测。 利用TIANGEN 公司的RNAprep Pure 多糖多酚植物总RNA 提取试剂盒提取各材料离层组织的总RNA, 后使用PROMEGA 公司的试剂对RNA 进行反转录,得到cDNA。 实时荧光定量聚合酶链反应(Quantitative real-time polymerase chain reaction,qRT-PCR)程序参照前期研究[11],采用BIO-RAD 公司的Sybr-Green-mix 试 剂, 利 用20 μL 体 系 进 行。 以GhUBQ7 基因为内参基因,本研究中所使用引物见表1, 所有试验进行3 次生物学重复。 使用Microsoft Excel 2016 对水处理和脱叶剂处理下各品种中的基因表达量进行单因素方差分析,判断不同处理下基因表达量差异显著性。

表1 本研究中使用的引物Table 1 Primers used in this study
2 结果与分析
2.1 脱叶剂敏感型材料的筛选
根据温室内各品种脱落剂处理前后叶片数目,计算处理后第4 天的脱叶率。 根据脱叶后表型及脱叶率(图1),筛选出6 个脱落率高的脱叶剂 敏 感 型 材 料:ZY35 (晋 棉28 号)、ZY210(57-681)、ZY305(晋棉24)、ZY321(辽棉6 号)、ZY335(鲁棉22)、ZY364(秦远4 号)和6 个脱落率低的脱叶剂不敏感型材料:ZY9 (鲁棉1 号)、ZY36 (晋棉36 号)、ZY41 (冀棉12 号)、ZY164(中棉所40)、ZY219(北农1 号)、ZY239(敦棉1号),上述材料名称在下文以编号代替。
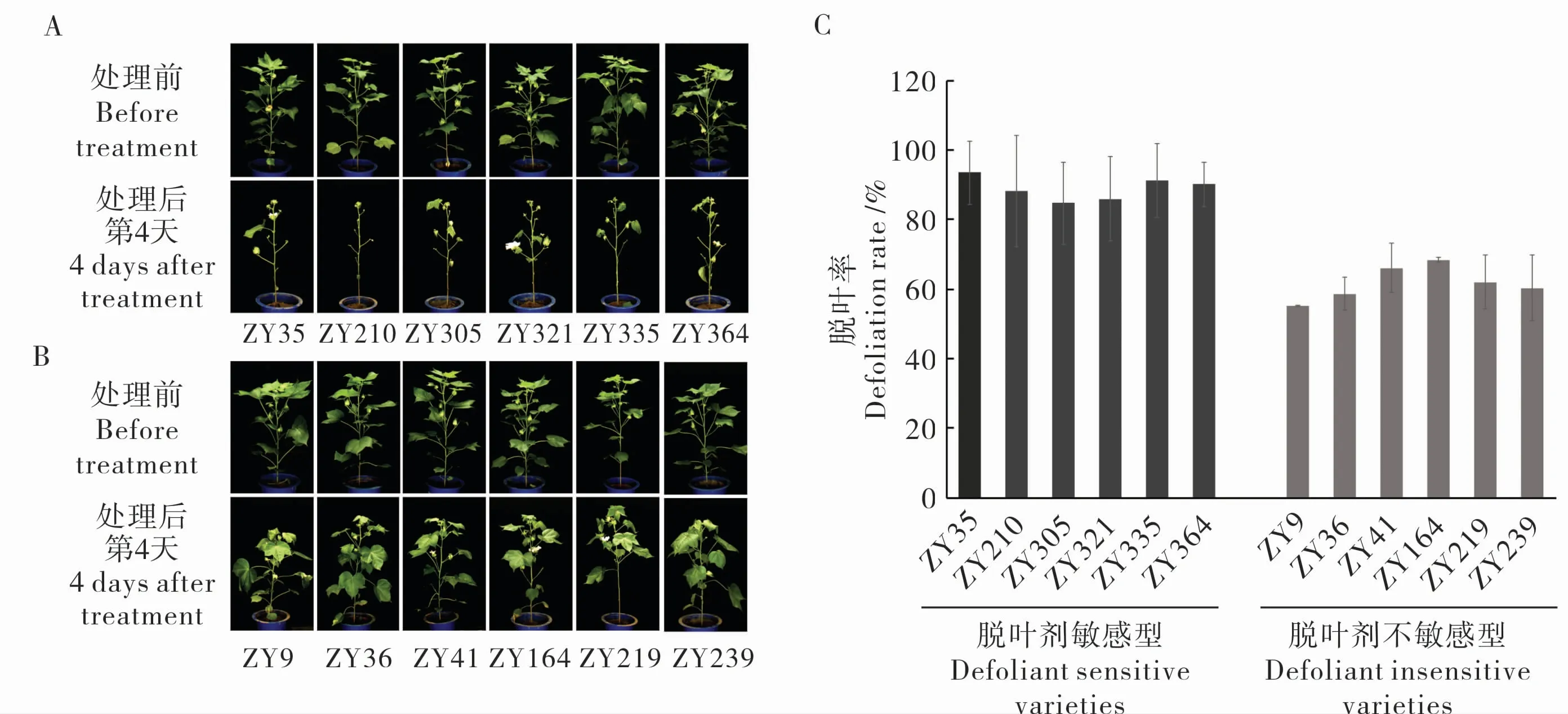
图1 脱叶剂处理后敏感型、不敏感型材料的表型及脱叶率Fig. 1 Phenotype and defoliation rate of defoliant sensitive and insensitive varieties after defoliant treatment
将温室中筛选出的脱叶剂敏感型和不敏感型棉花材料在大田种植, 由于种子量不足等情况,实际种入大田的棉花材料有脱叶剂敏感材料ZY35、ZY335、ZY364 和脱叶剂不敏感材料ZY9、ZY36、ZY164、ZY219、ZY239。 在处理前及处理后第7 天统计各棉花材料的叶片数,计算脱叶率,发现大田中敏感型材料脱叶率较高, 而不敏感型材料脱叶率相对较低(表2),与温室筛选结果相符。

表2 筛选出的棉花材料在脱叶剂处理后第7 天的脱叶率Table 2 Defoliation rates at the 7th day after defoliant treatment in field of the selected cotton varieties
2.2 细胞分裂素相关基因参与棉花响应脱叶剂处理
在脱叶剂处理后第1 天和第3 天,对脱叶剂敏感的ZY35、ZY364 和不敏感的ZY219、ZY239的离层组织取样,抽提RNA 并逆转录成cDNA,通过qRT-PCR 检测6 个细胞分裂素相关基因、9个乙烯相关基因及6 个水解酶活性相关基因在这4 个棉花材料中的表达模式。
结果(图2)显示,在脱叶剂敏感材料中,细胞分裂素氧化酶基因GhCKX3-1、GhCKX3-2、GhCKX3-3和GhCKX7 基因表达模式表现相似的趋势,但是差异表达的倍数不同。 相对于水处理对照,脱叶剂处理后第1 天和第3 天,4 个基因在脱叶剂敏感材料ZY35 的离层中均上调表达, 在脱叶剂处理后第1 天上调幅度更大 (GhCKX3-1 上调8.9倍,GhCKX3-2 上 调79.4 倍,GhCKX3-3 上 调140.8 倍,GhCKX7 上调120.0 倍);而在敏感型材料ZY364 中,GhCKX 基因表达在处理后第3 天达到较高水平(GhCKX3-1 上调3.3 倍,GhCKX3-2上调31.7 倍,GhCKX3-3 上调33.1 倍,GhCKX7上调65.8 倍)。 在不敏感型材料ZY239 中,GhCKX3-2、GhCKX3-3 和GhCKX7 基因的表达在脱叶剂处理后有微弱上调或基本没有上调(图2A~D);然而,不敏感型材料ZY219 经脱叶剂处理后所有GhCKXs 基因均上调表达,只是上调程度比敏感型材料ZY35 低。 细胞分裂素响应调节因子基因GhCTKRR2 和GhCTKRR9 则表现出复杂的表达模式。 与水处理对照相比,在脱叶剂处理后第1 天,GhCTKRR2 在2 个脱叶剂敏感材料中均上调表达,而在2 个脱叶剂不敏感材料中均下调表达;在脱叶剂处理后第3 天,除了脱叶剂敏感材料ZY364,其他3 个材料中GhCTKRR2均上调表达(图2E)。 在2 个脱叶剂敏感材料中和不敏感材料ZY239 中,GhCTKRR9 在脱叶剂处理后第1 天和第3 天均下调表达(图2F)。综上所述,大部分细胞分裂素相关基因参与棉花响应脱叶剂处理。
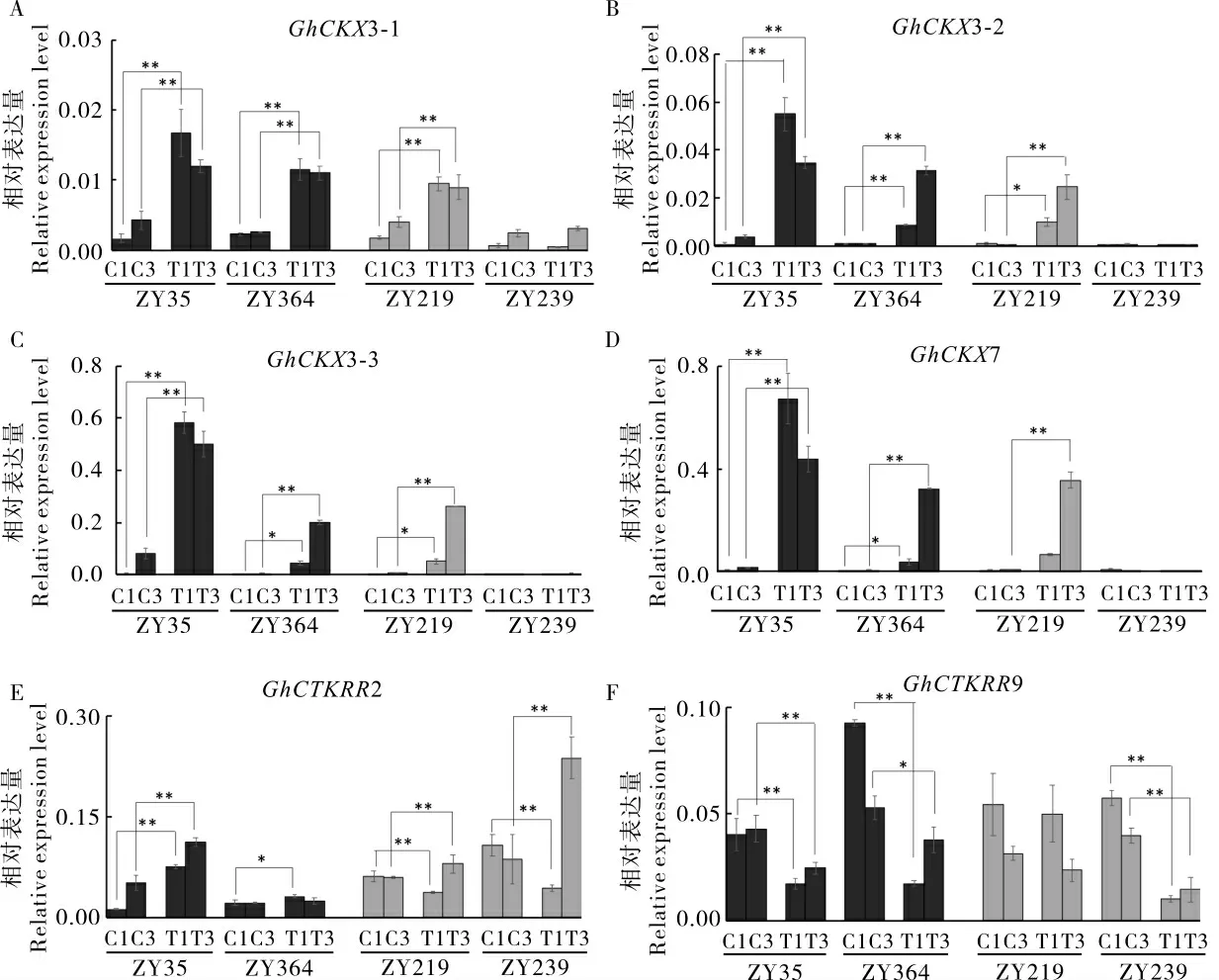
图2 细胞分裂素相关基因在不同棉花材料中的表达模式Fig. 2 Expression patterns of cytokinin-related genes in the selected cotton varieties
2.3 乙烯相关基因参与棉花响应脱叶剂处理
水处理下, 编码1- 氨基环丙烷-1- 羧酸(ACC)氧化酶的GhACCO 和GhACCO3 基因在4 个棉花材料中均低水平表达或不表达。 在敏感型材料ZY35 中, 脱叶剂处理后GhACCO 和GhACCO3 基因明显上调表达, 且在脱叶剂处理后第1 天的表达量上调幅度极高;而在不敏感型材料ZY219 和ZY239 中的表达量没有明显上升,或与敏感型材料相比上调幅度较低(图3A、B)。乙烯受体基因GhEIN3 和GhETR1 也呈现类似 的 表 达 模 式 (图3C、D)。 转 录 因 子 基 因GhERF113-1 和GhERF113-2 表达模式相似,脱叶剂处理后在敏感型材料ZY35 和ZY364 中上调表达; 而在不敏感型材料ZY239 中,GhERF113-1 和GhERF113-2 在处理后第1 天上调表达 (GhERF113-1 上调2.7 倍,GhERF113-2上调8.4 倍),在第3 天却显著下调表达(图3E、F)。在脱叶剂处理后第3 天,相对于水处理而言,GhERF61 在敏感型材料中上调表达, 而在不敏感型材料中下调表达(图3G);水处理下转录因子基因GhRAP23 在4 个材料中基本不表达,而在脱叶剂处理下均上调表达, 在敏感型材料中上调的幅度更大(图3H)。 上述结果说明乙烯相关基因参与棉花响应脱叶剂处理。
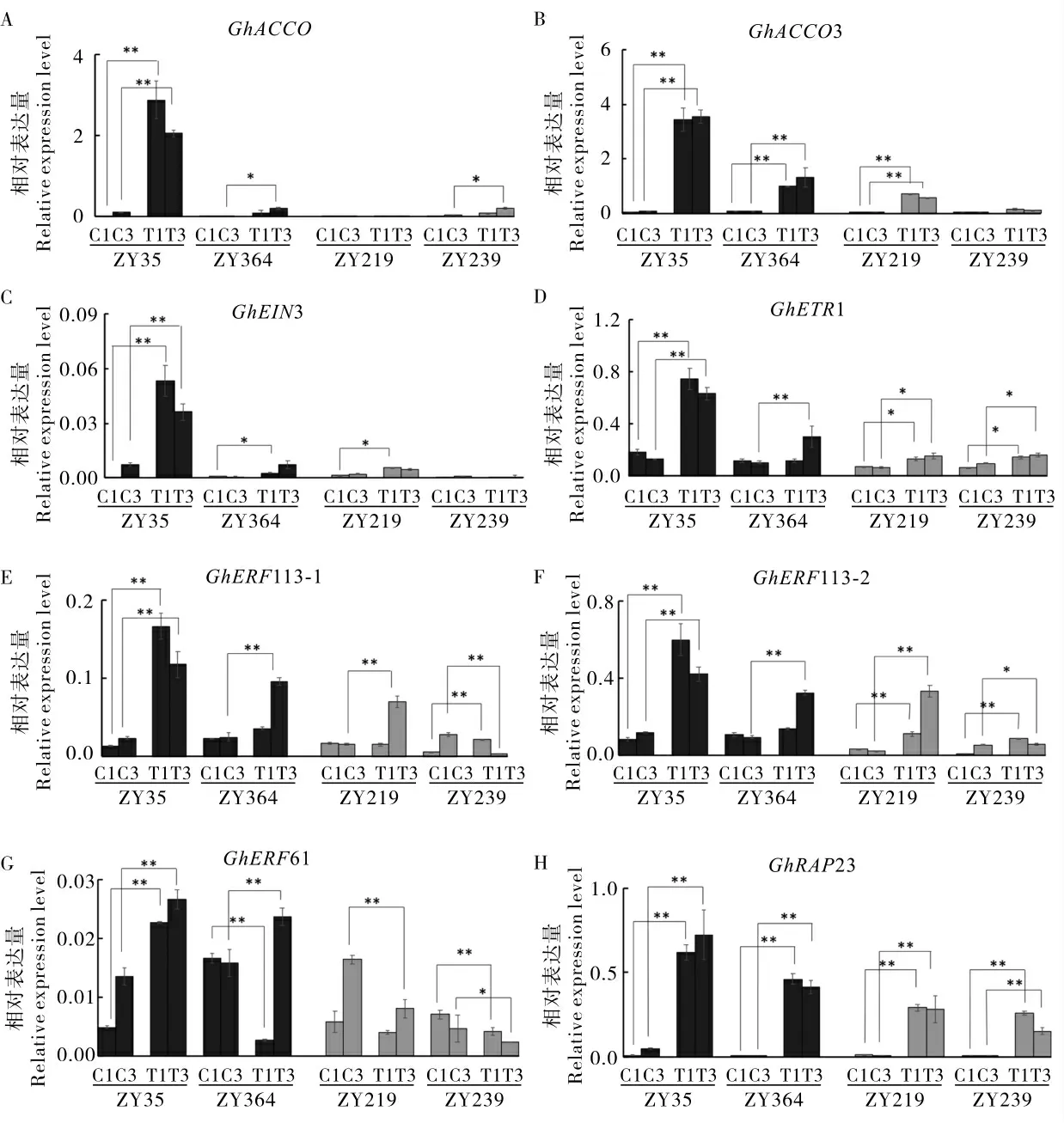
图3 乙烯相关基因在筛选的棉花材料中的表达模式Fig. 3 Expression patterns of ethylene-related genes in the selected cotton varieties
2.4 水解酶活性相关基因参与棉花响应脱叶剂处理
对6 个具有水解酶活性基因的表达量结果分析发现,4 个水解酶相关基因(GhBXL1、GhCHI1、GhE13B-1 和GhE13B-2)在4 个材料中,在水处理后的第1 天和第3 天基本不表达或者低量表达; 而在脱叶剂处理后第1 天,GhBXL1、GhCHI1、GhE13B-1 和GhE13B-2 在敏感型材料ZY35 中的表达量分别是水处理的629.4 倍、220.5 倍、45 287.3 倍和1 754.6 倍,在敏感型材料ZY364 中脱叶剂处理后也上调表达,但是上调倍数比ZY35 中低(图4)。 GhBXL1、GhE13B-1 和GhE13B-2 在脱叶剂不敏感型材料ZY239 中水处理和脱叶剂处理下均基本不表达 (图4A、C、D)。 编码核糖核酸酶的GhRNS1 基因在脱叶剂处理后的敏感型材料ZY35 中均上调表达 (第1天上调274.5 倍,第3 天上调8.7 倍),而该基因在不敏感型材料ZY239 中于脱叶剂处理后第3天下调表达(图4E)。 脱叶剂处理后,编码内切葡聚糖酶的GhGUN1 基因在2 个敏感型材料中相对于水处理均极显著上调表达;而在2 个不敏感材料中, 于脱叶剂处理后第1 天显著上调表达,在处理后第3 天的表达量无显著变化(图4F)。脱叶剂处理后,6 个具有水解酶活性的基因在脱叶剂敏感型材料离层组织中均有较高水平的上调表达,说明这些水解酶基因参与棉花响应脱叶剂处理。

图4 水解酶相关基因在筛选的棉花材料中的表达模式Fig. 4 Expression patterns of hydrolase genes in the selected cotton varieties
3 讨论
3.1 温室及大田的200 份资源材料筛选
在温室条件下,对广泛筛选得到的200 份资源材料进行了脱叶剂处理,并统计脱叶率,从而从200 份种质资源中筛选出敏感型材料ZY35、ZY210、ZY305、ZY321、ZY335、ZY364 和不敏感型材料ZY9、ZY36、ZY41、ZY164、ZY219、ZY239。为了与生产上田间脱叶剂处理一致,后续将部分棉花材料在大田种植, 于吐絮期进行脱叶剂处理,并根据表型及脱叶率筛选出敏感型材料ZY35、ZY364 和不敏感型材料ZY219、ZY239。
比较温室处理与大田处理的脱叶率,可以发现温室处理后第4 天的脱叶率比大田处理后第7天高。 温室处理时间为6 月中下旬,正值初花期;田间处理时间为10 月上旬,正值棉花的吐絮期,温室处理棉花处于旺盛生殖发育时期对外界因子更敏感; 且比起大田较为恶劣的自然环境,温室中棉花长期处于舒适的生长环境,所以对脱叶剂胁迫更敏感,较大田的棉花易于形成离层从而脱叶。
3.2 脱叶剂处理下棉花落叶特点
棉花正常叶片脱落常常由衰老引起,乙烯被认为是棉花落叶最关键的生理因素[3,27,29];研究表明, 处于旺盛期的功能叶对乙烯的反应最迟钝,而幼叶和衰老叶片对乙烯较敏感[27,30]。 化学脱叶是在脱叶剂作用下使代谢旺盛的功能叶叶柄处形成离层而被动脱落,除乙烯之外,还有其他的重要因素参与调控。 生产上使用的化学脱叶剂主要由乙烯利、噻唑隆及相关助剂组成,具有脱叶和催熟功能, 但是脱叶功能与催熟功能不能等同[3]。 研究发现脱叶剂敏感型材料在脱叶剂处理后,叶柄离层快速形成,叶片迅速脱落[11]。 而不敏感材料在脱叶剂处理后常表现3 种情况:第1 种是植株对脱叶剂没有反应, 正常完成生长发育,这种情况比较少见; 第2 种是叶柄离层形成缓慢,叶片脱落延缓;第3 种情况是植株对乙烯过于敏感,导致叶片迅速失水干枯,而不形成离层,叶片干枯不脱落。 现在生产上使用的机采棉材料较多较杂, 除了脱叶剂敏感型材料外, 有些第3种脱叶剂不敏感材料因为其强烈的乙烯敏感性也被归为适宜机采的材料。
本研究中通过脱叶率筛选出的脱叶剂敏感型和不敏感型材料在部分细胞分裂素相关基因、乙烯相关基因及水解酶相关基因表达上存在差异:总体上脱叶剂处理后这些基因在脱叶剂敏感型材料中均显著上调表达;然而这些基因在2 个脱叶不敏感型材料中的表达呈现不同的特点,这说明不敏感型棉花对脱叶剂的响应可能存在多种表现形式和调控模式。 另外,脱叶剂处理后,与乙烯响应相关的基因在脱叶剂敏感型和不敏感型材料中多上调表达,验证了乙烯参与棉花脱叶响应的结论[4,11]。而关于乙烯和细胞分裂素在响应棉花化学脱叶过程中的关系需要进一步探讨。
4 结论
从棉花种质资源中筛选得到脱叶剂敏感型和不敏感型材料,并进行化学脱叶分子机理的解析,对机采棉品种种质资源的评价、筛选和改良及脱叶剂组配和开发具有一定的理论意义。
猜你喜欢
纺织标准与质量(2022年2期)2022-07-12
人人健康(2022年11期)2022-07-07
煤(2022年3期)2022-03-17
煤田地质与勘探(2021年3期)2021-07-17
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
煤田地质与勘探(2021年1期)2021-03-30
农民致富之友(2017年4期)2017-04-10
现代农业科技(2016年23期)2017-04-06
中学教学参考·理科版(2016年2期)2016-04-05
