
毛白杨融合基因4CL1-CCR对烟草木质素沉积的影响*
2020-11-30 00:35胡嘉祺蒋湘宁
林业科学 2020年10期
胡嘉祺 齐 芪 蒋湘宁 盖 颖
(1.北京林业大学生物科学与技术学院 北京 100083; 2.中国农业大学园艺学院 北京 100193)
木质素是陆生植物细胞壁中含量仅次于纤维素的天然大分子聚合物,在植物的生长过程中有着十分重要的意义,为植物向上生长提供支撑力(Voelkeretal.,2011);并且在植物抵御生物侵害中起到了天然屏障作用(Vanceetal.,1980)。虽然木质素的存在对于植物十分重要,但它会降低传统造纸的出浆率和纸张质量,并且降低牲畜对草料的消化和吸收,所以长期以来木质素一直被当做废料排放到自然环境中(Baucher,2003)。但近些年发现木质素在工业生产中有着重要作用,如,作为粘合剂中酚类的替代物(Fangetal.,2011),作为塑料中的新型偶联剂(Justoetal.,2013)。由于在不同行业中对木质素有着不同的需求,所以对工业原料中木质素含量的调控有着重要意义。
木质素是一类由肉桂醇、松柏醇和芥子醇氧化聚合成的大分子物质,在木质素的生物合成和氧化聚合过程中共有11个酶参与,这些酶被分成木质素生物合成途径中参与纵向反应的酶PAL(苯丙氨酸裂解酶phenylalanine ammonia-lyase)、C4H(肉桂酸4-羟化酶cinnamate 4-hydroxylase)、4CL(4-香豆酸辅酶A连接酶4-coumarate:coenzyme A ligase)、CCR(香豆酰辅酶A还原酶 coumaryl:coenzyme A reductase)和CAD(肉桂醇脱氢酶cinnamyl alcohol dehydrogenase),参与横向反应即木质素单体之间转化的酶C3H(对香豆酸-3-羟基化酶p-coumarate-3-hydroxylase)、F5H(阿魏酸5-羟化酶 ferulate 5-hydroxylase)、CSE(咖啡酰莽草酸酯酶caffeoyl shikimate esterase)、COMT(咖啡酸-O-甲基转移酶caffeic acid-O-methyltransferase)和CCoAOMT(咖啡酰辅酶A氧甲基转移酶caffeoyl-CoA 3-O-methyltransferase),以及木质素单体氧化聚合的酶POD(过氧化物酶peroxidase)和LAC(漆酶laccase)。调控纵向反应的酶主要引起木质素总量的改变,通过下调4CL酶活性使得拟南芥(Arabidopsisthaliana)中G型木质素单体含量显著降低(Leeetal.,1997),通过抑制4CL基因表达获得的转基因烟草(Nicotianatabacum)、毛白杨(Populustomentosa)和柳枝稷(Panicumvirgatum)的木质素都明显减少(Kajitaetal.,1997;Voelkeretal.,2010;Xuetal.,2011;Tianetal.,2013)。在白桦杂交种(Betulaplatyphylla×B.pendula)中过表达CCR基因木质素含量增加了14.6%,而抑制CCR表达后木质素含量降低了6.3%(Zhangetal.,2015)。而调控横向反应的酶会影响木质素总含量和单体组成,在分别下调C3H和F5H的拟南芥中,F5H的表达量的改变只引起木质素各个单体比例的改变而并没有影响木质素总量,而C3H表达量的变化不仅减少了木质素总量也改变了3种木质素单体所占比例(Reddyetal.,2005)。
在以往的转基因研究中多是调控木质素合成途径中的单个基因(Wengetal.,2010;Prashantetal.,2011;Tamasloukhtetal.,2011),尚未见将木质素合成过程中两步酶促反应的酶融合为一个复合体来调控木质素代谢的报道。由于在植物体内4CL和CCR的催化是一个连续反应的过程,所以作者所在课题组将毛白杨(Populustomentosa)的4CL1和CCR基因的编码区连接在一起,构建了融合基因4CL1-CCR并在体外对其编码的融合蛋白4CL1-CCR进行酶活性分析,发现它具有4CL1和CCR 2个酶的催化活性(侯佳音,2014),并且此融合蛋白在大肠杆菌(Escherichiacoli)内可以高效催化4-香豆酸、咖啡酸和阿魏酸生成相应的醛类,与双转4CL1+CCR的大肠杆菌比较,转融合基因的菌株有更高的催化效率,增加了对底物酚酸的利用率(Liuetal.,2015)。在前期研究的基础上,本研究将毛白杨融合基因4CL1-CCR转到烟草中,分析了转基因烟草木质素含量和沉积状态的变化,期望在转基因植株中验证融合蛋白的功能并为今后融合基因的应用提供分子研究基础。
1 材料与方法
1.1 试验材料
重组原核表达载体pQE31-4CL1-CCR和无菌烟草组培苗均为实验室保存,融合基因为毛白杨4CL1(GenBank 登录号:AY043495.1)和CCR(GenBank 登录号:EU760388.1),将4CL1的终止密码子和CCR的起始密码子去除后,在2个基因中间添加一段DNA序列作为linker,这段DNA序列编码的是3个以4个色氨酸和1个苯丙氨酸为重复单位的多肽,这个多肽保证了融合酶的2个蛋白在空间上的独立性从而保证了2个酶的活性。
1.2 试验方法
1.2.1 烟草的转化及分子鉴定 利用PCR技术在融合基因4CL1-CCR两端引入酶切位点,再用酶切连接将其构建到植物表达载体pBI121上,并将重组载体转化到根癌农杆菌(Agrobacteriumtumefaciens)GV3101中,通过叶盘法获得转基因烟草。剪取2片通过抗性筛选(筛选培养基为MS+50 mg·L-1Kan)的烟草无菌苗叶片,用CTAB法提取基因组用于PCR鉴定。将PCR初筛的阳性的转基因烟草用Southern 杂交进一步鉴定。
1.2.2 烟草的木质素单体含量测定 将1月龄的转基因烟草和野生型烟草组培苗移栽至花盆中,截取移栽的3月龄转基因烟草和野生型烟草第5至第13茎节,经过液氮研磨后称取100 mg粉末,参照Tian等(2013)测定木质素时的前处理方法获得细胞壁组分,将细胞壁组分按Qi等(2019)方法对细胞壁木质素进行衍生化,反应完成后,取1 μL反应液进行GC/MS(Thermo Finnigan)(色谱柱:DB 5 MS 30 m×0.25 mm 0.25 μm Agilent USA)分析。每个样品重复3次,取平均值。
1.2.3 烟草的解剖结构观察 取移栽的3月龄转基因烟草和野生型烟草(非温室培养土培苗),将茎剪成5 cm长的段,用刀片将其切成薄且透明性好的完整切片放置于载玻片上,用盐酸-间苯三酚染色法对切片进行染色并在Nikon E200光学显微镜下观察,并通过Image Pro Plus软件处理照片,计算所有木质部细胞细胞壁厚度、木质部宽度和导管周长。
另取未经间苯三酚染色的烟草经切片后平铺于载玻片上,盖好盖玻片,在低倍物镜下找到烟草木质部,换高倍镜并切换到紫外灯下激发木质素使其自发荧光并进行观察拍照。
1.2.4 烟草茎的木质素合成基因半定量PCR 分别提取经PCR-Southern 杂交鉴定过的转基因烟草的总RNA,经反转录获得cDNA。以烟草的管家基因(Actin)为内参基因分别设计正向引物Actin-sense和反向引物Actin-antisense,以纵向反应基因(4CL1-CCR,CAD)和横向反应基因(C3H,COMT和F5H)为目的片段设计特异性引物,具体见表1。以内参基因调整PCR循环数和各烟草模板添加量,对各转基因株系进行半定量PCR分析。
2 结果与分析
2.1 叶盘法获得烟草转基因植株的分子鉴定
分别提取经抗性筛选的转基因烟草植株总DNA,以35S启动子部分序列+4CL1基因部分序列为模板设计特异引物进行PCR扩增,产物经1%琼脂糖凝胶电泳分析,阳性对照与部分筛选植株均扩增出了约1 100 bp的特异条带,将PCR产物用于Southern 杂交检测(杂交探针序列为35S启动子)。由图1可以看出经抗性筛选的烟草转基因株系具有明显的杂交信号,而野生型烟草中无信号,此结果可以证明引入的外源基因已经整合到烟草基因组中。经鉴定共获得5个转基因株系。

图1 转基因烟草的PCR-Southern 杂交鉴定Fig.1 Identification of transgenic tobacco plants by Southern Blotting+:阳性对照;1:WT;2,3,4,5,6:PCR鉴定阳性转正义株系4CL1-CCR 2、4CL1-CCR 5-2、4CL1-CCR 10-1、4CL1-CCR 16-1、4CL1-CCR 20-2。+:Positive control;1:WT;2,3,4,5,6:Positive plants identified by PCR,4CL1-CCR 2,4CL1-CCR 5-2,4CL1-CCR 10-1,4CL1-CCR 16-1 and 4CL1-CCR 20-2,respectively.
2.2 转基因烟草木质素单体含量分析
采用优化后的硫代硫酸解法断裂木质素的链接键后结合气相色谱质谱联用仪来测定转基因烟草的木质素单体含量,该方法具有减少试验操作时间、减少待测样品需求量、可重复性好和灵敏度高等优点,因为在硫代硫酸解过程中破坏掉的是木质素单体中的β—O—4链接键,该链接键占木质素链接键的65%~75%,所以该方法常用来裂解分析木质素单体组成(Qietal.,2019)。该方法中加入二十四烷作为内标可在定性的同时做到对待测物的定量分析(图2)。过表达异源4CL1-CCR融合基因,不仅可以影响烟草细胞壁中木质素的含量,也可以改变木质素单体的组成(表2)。与野生型对照相比,转基因烟草不同株系中,G+S型木质素含量均升高,从野生型的16.59%升高到23.62%(4CL1-CCR 5-2)。在所有转基因株系中G型木质素单体均有增加,其中4CL1-CCR 5-2增幅为34.64%;而同时S型木质素的含量在不同株系中变化并不一致,在株系4CL1-CCR 5-2和4CL1-CCR 2中S型木质素单体有一定升高,但在其他转基因株系中无显著变化。虽然在不同株系中G型木质素和S型木质素含量变化不一致,但在所有转4CL1-CCR的烟草中G∶S都有所升高(表2)。另一方面,在所有烟草株系(WT和4CL1-CCR)中都未检测到H型木质素单体。

图2 转基因烟草细胞壁木质素单体气相色谱峰Fig.2 Characteristic peaks of monolignols in transgenic tobacco cell wall analyzed by gas chromatographIS:内标;G:G型木质素单体;S:S型木质素单体。IS:Internal standard;G:G-unit monolignol;S:S-unit monolignol.

表2 转基因烟草中木质素单体的组成①Tab.2 Lignin monolignols composition of control and transgenic tobacco
2.3 转基因烟草茎的解剖分析
经GC-MS测定木质素可知,株系4CL1-CCR 5-2中木质素含量增加最多,株系4CL1-CCR 20-2木质素含量增加最少,分别对这2个转基因烟草株系和野生型烟草的茎横截面进行显微观察。在靠近烟草植株基底部3 cm处截取茎段用间苯三酚染色观察发现,转基因烟草的木质部宽度明显大于野生型对照组(图3A、B、C),且转基因烟草的导管内径增大。这一结果与Tian等(2013)和Chen等(2019)分别在毛白杨和烟草中转入单个4CL基因引起的植物木质部变化一致。
木质素在紫外线照射下可以产生强烈的荧光且荧光的亮度与木质素局部密度呈正相关。为更好地揭示转基因烟草细胞壁中木质素变化的空间信息,用紫外显微镜对WT和4CL1-CCR株系的成熟茎横截面进行观测。结果显示,在野生型烟草与转基因烟草中木质素沉积密度的分布均为细胞角隅(图3D、E、F中红色箭头所示)和复合胞间层密度高,而次生细胞壁中木质素沉积密度低。由结果可知融合基因4CL1-CCR并未改变木质素沉积的状态,即高密度区依然在细胞角隅和复合胞间层,低密度区依然在次生壁。
用Image Pro Plus软件统计木纤维细胞壁厚度、导管壁周长及木质部宽度(图3A、B、C中绿线所示)。与野生型烟草相比,转融合基因4CL1-CCR的烟草木纤维细胞壁显著增厚,其中4CL1-CCR 5-2株系增加17.2%,4CL1-CCR 20-2株系增加7.11%(图3G);导管显著加粗,其中4CL1-CCR 5-2株系导管周长增加100.2%,4CL1-CCR 20-2株系增加43.66%(图3H);木质部宽度增加,其中4CL1-CCR 5-2株系增加86.4%,4CL1-CCR 20-2株系增加44.6%(图3I)。

图3 转基因烟草茎横截面解剖结构Fig.3 The transverse anatomical structure of stem of transgenic tobaccoA,D:WT;B,E:4CL1-CCR 20-2;C,F:4CL1-CCR 5-2.A,B,C:间苯三酚染色图(绿线:木质部宽度);D,E,F:木质素自发荧光图(红色箭头为细胞角隅,白色箭头为复合胞间层);G,H,I:转基因烟草茎结构元件测量。*表示平均值差异显著(P <0.05),用ANOVA法做差异分析,用Dunnett做事后检验;下同。A,B,C:Phloroglucinol stain(Green line:Xylem width);D,E,F:Lignin autofluorescence(Red arrow:Cell corner;White arrow:Compound middle lamella);G,H,I:Measurements of some elements of stem.Significantly different values at P<0.05 are represented by asterisk(*),and determined by ANOVA followed by Dunnett’s post-hoc test;the same below.
2.4 木质素生物合成相关基因的半定量PCR分析
通过对木质素单体合成途径中的4个相关基因的表达进行半定量PCR分析后发现,调控4CL1-CCR基因的表达将会对下游基因产生协同调控作用(图4)。位于CCR下游的CAD和位于4CL1下游的C3H和COMT的表达量都有不同程度的升高,但是F5H变化不显著。C3H和COMT都参与了木质素单体从H型向G型的转化过程,因为这2个基因的表达量升高所以在转基因烟草中G型木质素含量增加。在G型木质素单体向S型木质素单体转化过程中最关键的酶是F5H,但在转基因烟草中F5H基因表达量没有显著变化,所以与WT相比转基因烟草的S型木质素含量变化并不显著。
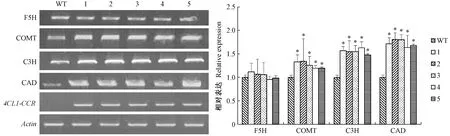
图4 烟草木质素合成基因半定量PCRFig.4 The semi-quantitative PCR of lignin biosynthesis genes in tobacco 1:4CL1-CCR 2;2:4CL1-CCR 5-2;3:4CL1-CCR10-1;4:4C1L-CCR 16-1;5:4CL1-CCR 20-2.F5H:阿魏酸5-羟化酶;COMT:咖啡酸O-甲基转移酶;C3H:香豆酸-3-羟化酶;CAD:肉桂醇脱氢酶;4CL1-CCR:融合基因。F5H:Ferulate 5-hydroxylase;COMT:Caffeic acid O-methyltransferase;C3H:Coumarate-3-hydroxylase;CAD:Cinnamyl alcohol dehydrogenase;4CL1-CCR:Fusion gene.
3 讨论
本研究将2个功能上有联系的基因融合后在烟草细胞内表达,发现引入此融合基因4CL1-CCR使得烟草细胞壁中木质素总量增加了36.44%;由于4CL1和CCR均为木质素合成中参与纵向反应的酶,木质素的增加表现为G型木质素单体增加,而S型单体变化不显著。
对转基因烟草进一步做半定量分析发现,在上调4CL1-CCR基因表达后增加了CAD的表达,CAD参与了木质素单体合成的最后一步酶促反应。在被子植物中CAD对酚醛的催化被认为没有底物特异性(Chaoetal.,2014),但在烟草中可以检测到G型和S型木质素单体,但是并未检测到H型木质素单体,推测烟草的CAD蛋白对对羟基肉桂醛的催化活性可能很低。对于参与横向反应的基因影响表现为转基因烟草中C3H和COMT表达量升高,其中C3H和COMT是H型木质素单体转向G型木质素单体必须的2个酶,因此,在纵向反应蛋白4CL1和CCR含量增加后,这2个酶催化产生的产物不能被进一步催化生成羟基肉桂醇,而是由C3H和COMT更多地转化成为G型的前体物质,最终表现为G型木质素单体含量显著增加。4CL1可以催化肉桂酸、阿魏酸和咖啡酸生成相应的CoA酯,但不能催化芥子酸(侯佳音,2014),所以S型木质素单体的生物合成需要经COMT和F5H催化G型木质素的前体物质才可以生成,在这一过程中最重要的F5H的表达量没有改变,所以S型木质素单体含量与野生对照植株相比无显著差异。
由烟草茎的盐酸-间苯三酚染色结果可以发现转基因烟草和野生型烟草木质素均沉积在木质部。虽然融合基因4CL1-CCR是在非组织特异性的强启动子CaMV35S(花椰菜花叶病毒35S启动子)下启动表达,但是木质素单体合成过程中其他基因具有组织特异性(MacMillanetal.,2017),只在木质部细胞中可以形成完整的木质素生物合成途径,因此融合基因并没有改变木质素出现的组织特异性。但是融合基因的表达使转基因烟草木质化的细胞增加最终表现为木质部宽度增加,木质部中导管壁周长增加,木纤维细胞壁厚度明显增加(图3)。这种调控木质素单体合成的基因影响植物导管形态及木质部厚度在多种转基因植物中均有报道,下调杂交杨树(Populustremula×P.alba)CCR基因表达发现其木质部中导管明显塌陷(Lepleetal.,2007);上调CCR的白桦杂交种中导管数量增加,并且呈现出异常的导管成串现象,从而增加导管占木质部的面积比例(Zhangetal.,2015);下调CCR表达的番茄(Solanumlycopersicum)茎中被染色的木质部细胞层数少于野生型(Van der Restetal.,2006);在抑制4CL基因表达的新西兰辐射松(Pinusradiata)茎的横切面中,木质化细胞的面积显著低于野生型(Wagneretal.,2009);过表达4CL基因的烟草(Chenetal.,2019)和毛白杨(Tianetal.,2013)中也观察到了木质部细胞数量增多。
由荧光显微观测可知,转融合基因4CL1-CCR的烟草和野生型烟草中具有相同的木质素密度分布,即在细胞角隅和复合胞间层中木质素密度高,而在次生细胞壁中木质素密度低。但与野生型烟草对比,转基因烟草中木质素高密度区域的面积有所增加,在次生壁中木质素沉积的面积增加。这种基因影响木质素沉积的现象与已报道的研究结论一致。在下调CAD基因表达的毛果杨(Populustrichocarpa)中,高密度木质素区域(细胞角隅和复合胞间层)和低密度木质素区域(次生细胞壁)的面积都减少,但是并没有出现木质素密度分布紊乱现象(Segmehletal.,2019)。推测控制木质素密度分布可能与细胞木质化进程有关,细胞木质化过程首先在细胞角隅和复合胞间层中出现木质素然后再沉积在次生细胞壁中。并且在细胞木质化的前期主要积累的是G型木质素单体,G型木质素单体可以形成更多的链接键,所以此时的木质素网状结构紧密呈现出高密度,当次生壁加厚时S型木质素单体逐渐增加而形成的网状木质素结构相对松散所以呈现出低密度状态(Terashimaetal.,1988)。在本研究中融合基因使烟草细胞壁中G型木质素单体含量增加,所以增加了转基因烟草的木质素高密度区面积,但并不能改变木质素单体在细胞壁区域内沉积的顺序,所以并未改变木质素密度的分布状态。
4 结论
由毛白杨4CL1和CCR基因构建的融合基因4CL1-CCR不仅增加了转基因烟草茎中木纤维细胞壁的厚度,同时也使得烟草茎木质部宽度增加、木质化细胞数量增加,最终表现为木质素含量增加。但是由于这2个基因编码的酶均参与木质素生物合成中纵向反应,其自身的表达量升高只增加了木质素单体反应中纵向反应量,通过半定量PCR分析,发现横向反应的关键基因F5H并未受到外源基因的影响,所以木质素单体组成中只有G型单体的含量增加。融合基因4CL1-CCR在转基因烟草中的表达并未改变木质素沉积的状态,即细胞角隅和复合胞间层为高密度木质素区域,而次生细胞壁为低密度木质素沉积区域,但是转基因烟草中高密度的复合胞间层面积略有增加。
猜你喜欢
亚热带植物科学(2022年1期)2022-05-17
植物研究(2021年2期)2021-02-26
三农资讯半月报(2020年18期)2020-10-14
四川农业科技(2019年5期)2019-07-01
数码世界(2018年1期)2018-12-23
浙江工业大学学报(2017年5期)2018-01-22
广东农业科学(2017年5期)2017-08-29
浙江柑橘(2016年1期)2016-03-11
天津农业科学(2015年11期)2015-12-03
安徽农学通报(2014年7期)2014-04-29
