
生鲜猪肉肌细胞内外间隙和水分状态与持水性的关系
2020-11-27 10:45李华健杨波若李燕清卞健科舒国涛
食品科学 2020年21期
李华健,陈 韬,杨波若,李燕清,卞健科,舒国涛
(云南农业大学食品科学技术学院,云南 昆明 650201)
中国是猪肉生产和消费大国,2018年猪肉产量达5 404万 t,约占世界猪肉总产量(1.13亿 t)的48%[1],猪肉品质的优劣直接影响着我国肉类工业的经济效益。肉的持水性即肉类保持自身水分和人为加入水分的能力[2-3],是影响肉品质量和企业经济效益的主要指标之一,业内常用汁液流失率来反映肉的持水性[4-5]。据估计,肉的汁液流失率每增加1%就可能给一个大型肉类加工厂每天造成数千元的经济损失[6]。所以,汁液流失形成的机制和控制技术一直是肉类科学研究的热点。其中,肌肉微结构变化与持水性的关系是主要研究方向之一。Huff-Lonergan等[7-8]提出肌细胞结构的改变可以控制肌细胞和肌肉中水分的保留或损失,其认为宰后僵直过程中肌细胞间和肌束间空隙(又称为汁液流失通道)的形成会导致较多的汁液流失;李侠等[9]观察到猪肉贮藏过程中纤维束间隙增大,不易流动水逐渐态变为自由水,肉的持水性降低;Schäfer等[10]的研究表明肌细胞外间隙面积的变化解释了汁液流失率变异的39%。由于大多数研究采用光学显微镜观测肌肉微观结构,不能分清汁液流失通道形成于肌细胞内还是肌细胞外,所以,汁液流失通道形成位置依然存在争议。如Kristensen[11]和Bertram[12]等认为汁液流失通道形成于肌细胞外,而Lawson等[13]的汁液流失通道形成假说则认为汁液流失通道形成于肌细胞内。近年来,低场核磁共振技术被广泛应用于肉与肉制品的水分状态和含量的测定及相关研究中[14]。Han Minyi等[15]通过研究发现低场核磁共振的横向弛豫时间(T2)和猪肉肌原纤维蛋白凝胶微观结构变化具有良好的相关性。因此,本研究结合透射电子显微镜和低场核磁共振技术测定肌原纤维与肌细胞膜之间的距离(肌细胞内间隙)、肌细胞之间的距离(肌细胞外间隙)和不同状态水分的相对含量,分析它们与汁液流失率的关系,以便更好地了解汁液流失形成的原因。
1 材料与方法
1.1 材料与试剂
选择品种相同、 饲养条件相同、 体质量((105±5)kg)相似的商品肉猪在曲靖市某商业屠宰厂进行屠宰。
体积分数25%戊二醛、乙醇(均为分析纯) 广东西陇化工厂;丙酮(分析纯) 湖北齐飞医药化工有限公司;环氧树脂(Epon-618) 天门恒昌化工有限公司;柠檬酸铅 湖北津乐达化工有限公司;醋酸铀 北京中科光析化工技术研究所;四氧化锇 山东西亚化学工业有限公司;二甲苯(分析纯) 上海试剂厂;甲醛(分析纯) 汕头市达濠精细化学品有限公司。
1.2 仪器与设备
H I 9 0 2 5 C 便携式p H 计 意大利哈纳公司;CR-400/410色差仪 日本美能达公司;JEM-1011透射电子显微镜 日本电子株式会社;R型超薄切片机切片德国徕卡公司;NMI20-Analyst核磁共振成像分析仪上海纽迈电子科技有限公司;1、2 cm双片刀由实验室自制。
1.3 方法
1.3.1 生鲜猪肉的处理及分组
按照Warner等[16]的方法,根据宰后45 min的pH值和肉色选择出10 条疑似正常背最长肌肉和10 条疑似PSE(pale, soft, exudative)背最长肌肉。于宰后不同时间点用背最长肌测定相关指标,根据汁液流失率大小经聚类分析,将样品分为高汁液流失率组和低汁液流失率组。
1.3.2 pH值和肉色的测定
采用王娟等[17]的方法,在宰后45 min和24 h分别测定背最长肌的pH值、亮度(L*)、红度(a*)和黄度(b*)。每个样品测量3 次取平均值。
1.3.3 低场核磁共振测定肌肉中水分分布
在宰后12 h和24 h取背最长肌,参照黄子信等[18]的方法测定,具体操作修改如下:重复扫描次数为8、重复采样等待时间为3 500 ms、模拟增益为20、数字增益为3、回波个数为2 000、谱宽为100 kHz。油样预热仪器30 min后,沿肌纤维方向将肉样分割成2 cm(长)×1 cm(宽)×1 cm(高)大小的肉样(约2.0 g),放入核磁小管中并在管口用塑封膜包住,在32 ℃、19 MHz共振频率下,使用弛豫时间编辑CPMG(Carr-Purcell-Meiboom-Gill)脉 冲 序 列 ( 9 0 ° 脉 冲 和1 8 0 ° 脉 冲 之 间 的 时 间τ=200 μs)进行测定,反演得到不同状态水的相对峰面积比例(相对含量),每个样品测定3 次,取平均值。
1.3.4 透射电子显微镜观测肌肉微观结构变化
宰后45 min,3、9、12 h和24 h在背最长肌上切取4 mm(长)×2 mm(宽)×2 mm(高)的肉样放入体积分数3.5%戊二醛溶液中固定,保藏在4 ℃环境中,用于透射电子显微镜观测。样品的处理参照Luo Xin等[19]的方法,即在4 ℃环境中,将固定好的各时间点样品用0.1 mol/L磷酸缓冲液(pH 7.2)冲洗3 次,每次10 min。之后用体积分数为1%的锇酸后固定1.5 h,分别用体积分数为50%、70%、80%、90%和100%的乙醇进行逐级脱水,每次15 min。用Epon-618渗透包埋,采用超薄切片机切片,经醋酸铀、柠檬酸铅双染色后用透射电子显微镜放大4 000 倍对每个样品拍摄5 张图片,使用Image-Pro Plus 5.1软件测定肌细胞内外间隙的平均距离。
1.3.5 汁液流失率的测定
参照Honikel等[20]的方法测定汁液流失率,并略作修改:宰后24 h,在13肋处切取2.5 cm厚背最长肌一片,称质量m1,悬于塑料袋中,充气,肉样不能和塑料袋接触,在4 ℃下吊挂24 h,取出称质量m2,汁液流失率按下式计算。

1.4 数据处理与分析
采用SPSS 19.0软件计算数据,以平均值±标准误表示,并进行Duncan多重比较、独立性T检验、Pearson相关性分析和逐步回归分析。P<0.05认为具有统计学意义。在进行统计分析之前先对原始数据进行正态性检验,根据GB/T 4882—2001《数据的统计处理和解释正态性检验》[21],只有宰后45 min的肌细胞内间隙不服从正态分布,经lgx转换后服从正态分布,使用转换后的值进行统计分析。
2 结果与分析
2.1 肉样汁液流失率的聚类分析结果
根据样品汁液流失率的大小进行聚类分析。如图1所示,当欧氏距离增至10.5时,将20 个样品分成高汁液流失率组(n=10)和低汁液流失率组(n=10),两组汁液流失率的范围分别为5.01%~6.36%和0.57%~3.98%。
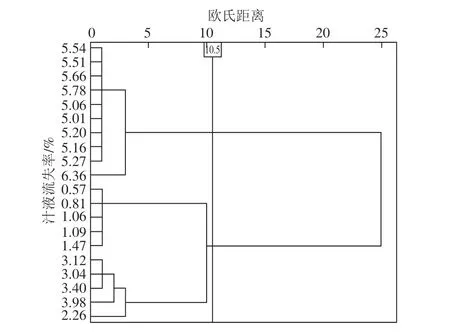
图1 样品汁液流失率的聚类分析树状图Fig. 1 Dendrogram obtained from cluster analysis of the drip loss rate of pork samples
2.2 两组肉样的pH值和肉色比较结果
如表1所示,宰后45 min,低汁液流失率组的pH值极显著高于高汁液流失率组(P<0.01),L*值极显著低于高汁液流失率组(P<0.01),此结果与何凡等[22]将不同品种羊肉分为不同滴水损失组所测得结果一致。Chmiel等[23]的研究也发现,与正常肉相比,宰后24 h,汁液流失严重的PSE肉具有较低的pH值和较高的L*、a*和b*值。综上,说明高低汁液流失率组之间的pH值和肉色存在一定的差异。

表1 高低汁液流失率组肉样的肉质指标(n= 10)Table 1 Meat quality attributes of pork samples from high and low drip loss rate groups (n= 10)
2.3 低场核磁共振横向弛豫时间(T2)分析结果
利用低场核磁共振反演计算的各个峰面积比例可以反映样品中不同状态水分的比例,峰面积比例变化情况可以表征样品中不同状态水分的分布状态和流动迁移情况[24]。图2为低场核磁共振测定的T2反演图谱,出峰位置位于T2b(0.1~10 ms)、T21(10~100 ms)和T22(100~1 000 ms),分别对应结合水、不易流动水和自由水,将3 种状态水的相对含量分别记为P2b、P21及P22。由于结合水与非水组分结合的牢固程度不同会导致结合水出现2 个信号峰[25]。对于该组分,根据陈琳莉等[25]的方法将两个峰面积比例叠加后得到结合水相对含量P2b。

图2 宰后12 h和24 h肉样的T2弛豫时间分布Fig. 2 Distribution of T2 relaxation time in pork samples at 12 h and 24 h post-mortem
不同状态水的相对含量如表2所示,在宰后12 h和24 h,结合水相对含量(P2b)在高、低汁液流失率组之间都没有显著差异(P>0.05);宰后24 h,高汁液流失率组的不易流动水相对含量(P21)极显著低于低汁液流失率组(P<0.01),而自由水相对含量(P22)极显著高于低汁液流失率组(P<0.01)。这与甄少波等[24]的研究结果一致,表明P21和P22与汁液流失率的关系较为密切,汁液流失率越高的肉,其P22越大,P21越小。

表2 肌肉中3 种状态水分相对含量的变化Table 2 Changes in relative contents of three water states in muscle at different times post-mortem
2.4 肌细胞内外间隙的变化
从结构上讲,肌肉中的汁液流失与排水力(肌原纤维和细胞的收缩)、膜通透性、细胞骨架蛋白降解、汁液流失通道的形成以及蛋白质之间的网络结构变化有关[4]。Mohan等[26]认为宰后肌肉的僵直会对肌肉的超微结构和品质性状(特别是质构、嫩度和持水性)产生显著影响。由于僵直过程中肌原纤维横向和纵向收缩,水从肌原纤维内转移到肌浆中,并由细胞膜维持在肌细胞内,使得肌细胞内肌浆空间增大[4,27]。所以在本研究中观察到肌细胞内外间隙的增加。
如图3所示,在宰后45 min,低汁液流失率组肉样肌原纤维结构排列紧密整齐,细胞膜附着在外肌原纤维上,只有轻微的分离(图3A1);与此相反,高汁液流失率组肌细胞外间隙比低汁液流失率组大,肌原纤维之间相互分离,膜也呈现大面积分离(图3A2);宰后12 h,低汁液流失率组肌细胞外间隙有所增加,肌细胞膜从外肌原纤维上分离(图3B1),相对于低汁液流失率组,高汁液流失率组肌细胞膜与外肌原纤维呈现更大程度的分离,肌细胞外间隙也更大(图3B2);宰后24 h,低汁液流失率组肌细胞膜呈现更大程度的分离,肌细胞外间隙也变得更大(图3C1),高汁液流失率组肌细胞外间隙相对宰后12 h稍微变小,肌细胞膜依然呈现大面积的分离(图3C2)。Veiseth-Kent等[28]对宰后13 d的牛肉微观结构进行观察也发现肌束膜从肌纤维上脱离,肌纤维间产生空隙。Chmiel等[23]的研究结果表明,宰后24 h正常肉的肌肉组织还保持相对完整,而PSE肉的肌肉组织遭到破坏。说明宰后早期高汁液流失率肉样比低汁液流失率肉样的微观结构变化更明显。

图3 宰后45 min(A)、12 h(B)和24 h(C)不同汁液流失率肉样的透射电子显微镜图Fig. 3 Transmission electron micrographs of muscle samples with different drip loss rate at 45 min (A), 12 h (B) and 24 h (C) post-mortem
为量化宰后猪肉肌细胞超微结构变化,使用图像分析软件Image-Pro Plus 5.1对肌细胞内间隙和肌细胞外间隙进行测量。如表3所示,低汁液流失率组的肌细胞内外间隙在宰后逐渐增大,宰后12 h和24 h没有显著差异,但都显著大于宰后45 min的肌细胞内外间隙(P<0.05)。高汁液流失率组肌细胞内外间隙宰后的变化不同于低汁液流失率组,高汁液流失率组的肌细胞内间隙在宰后12 h达到最大,而后呈减小趋势,肌细胞外间隙在宰后12 h达到最大,在宰后24 h显著减小(P<0.05)。Schäfer等[10]的研究也发现肌纤维束之间的空隙面积在3~9 h逐渐增大,在9~24 h减小。可能是由于骨架蛋白降解较快使得肌原纤维间连接遭到破坏产生横向膨胀[4,11],导致汁液流失率高的肉肌细胞内外间隙减小。
高汁液流失率组的肌细胞内间隙在宰后45 min显著大于低汁液流失率组(P<0.05)。高汁液流失率组的肌细胞外间隙在宰后9 h 显著大于低汁液流失率组(P<0.05)。表明宰后高汁液流失率组的肌原纤维收缩更快,形成的空隙更大。这与Hughes等[27]认为汁液流失率高的肉肌原纤维结构收缩更快,细胞外空隙面积更大的观点一致。较大的空隙会导致肌细胞外毛细管力减弱,使得肌细胞外自由水容易形成汁液流失[29]。采用低场核磁共振观察到高汁液流失率组的P22高于低汁液流失率组(表2)也证实了这一现象。
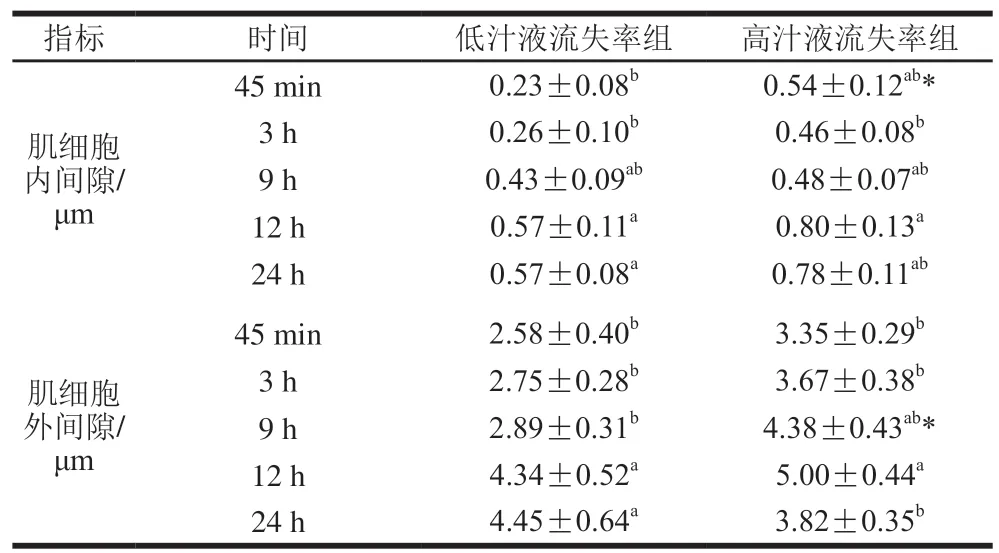
表3 两组肉样肌细胞内外间隙的差异(n=10)Table 3 Intra- and extracellular spaces in muscle samples from low and high drip loss groups (n= 10)
2.5 肌细胞内外间隙、不同状态水分相对含量和汁液流失率间的相关性和回归分析结果
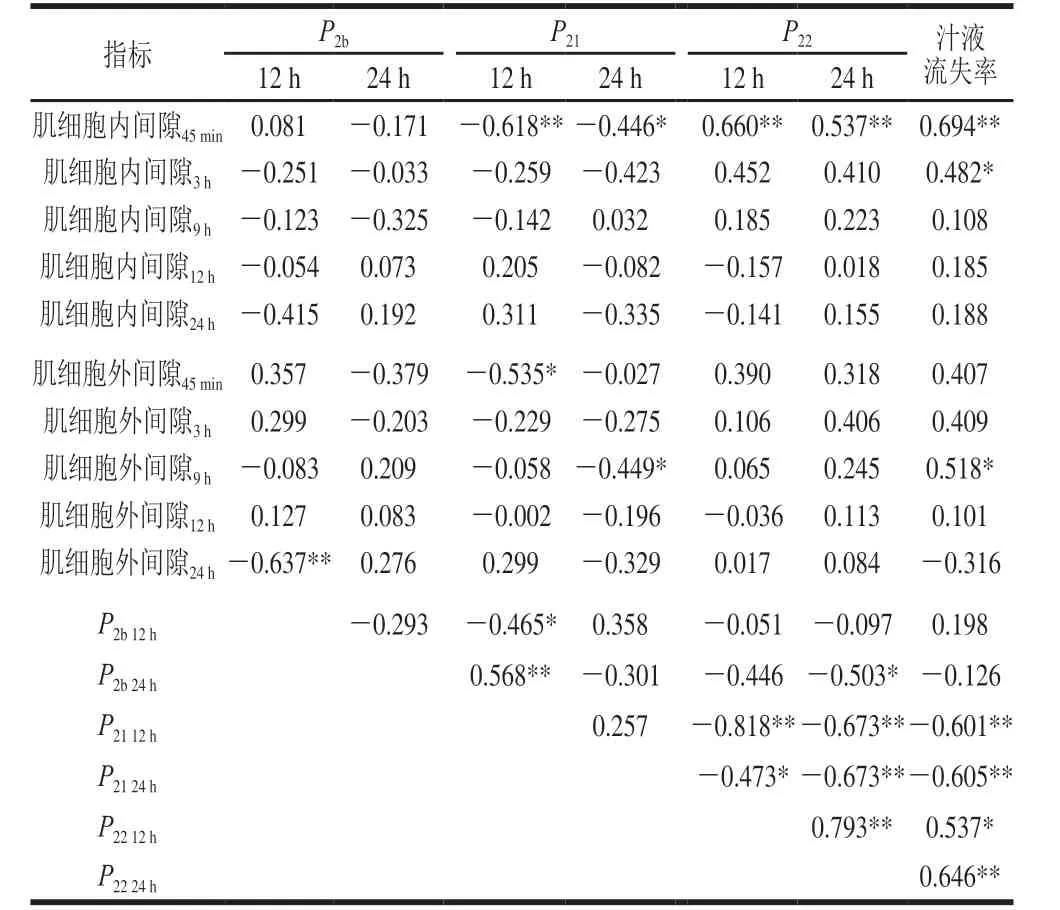
表4 肌细胞内外间隙、不同状态水分相对含量和汁液流失率间的相关性分析Table 4 Correlation analysis of intra- and extracellular spaces of myocytes with relative contents of three water states and drip loss rate
魏秀丽等[30]的研究结果表明结合水与不易流动水呈极显著负相关(P<0.01),不易流动水与自由水呈极显著负相关(P<0.01),不同状态水之间存在相互“态变”。本研究结果与之相似(表4),在宰后12 h,P2b与P21呈显著负相关(P<0.05),P21与P22呈极显著负相关(P<0.01)。在宰后24 h,P2b与P22呈显著负相关(P<0.05),P21与P22呈极显著负相关(P<0.01)。结果表明3 种状态水确实如魏秀丽等[30]所述,结合水会“态变”为不易流动水,不易流动水与自由水可以发生相互“态变”。
肌细胞内间隙在宰后45 min与宰后12 h和24 h的P21呈显著负相关(P<0.05,P<0.01)(表4)。肌细胞外间隙在宰后45 min与宰后12 h的P21呈显著负相关(P<0.05),在宰后9 h与宰后24 h的P21呈显著负相关(P<0.05)(表4)。宰后9 h的肌细胞外间隙解释了宰后24 hP21变异的15.7%(表5),相关系数=标准偏回归系数=-0.449。说明宰后9 h的肌细胞外间隙对宰后24 h的不易流动水影响最大,肌细胞外间隙越大,不易流动水含量越少。
宰后45 min的肌细胞内间隙与宰后12 h和24 h的P22呈极显著正相关(P<0.01)(表4),解释了宰后24 hP22变异的24.8%(表5)。说明在肌细胞内外间隙中,宰后45 min的肌细胞内间隙对P22的直接影响较大,宰后45 min的肌细胞内间隙越大自由水含量越多。
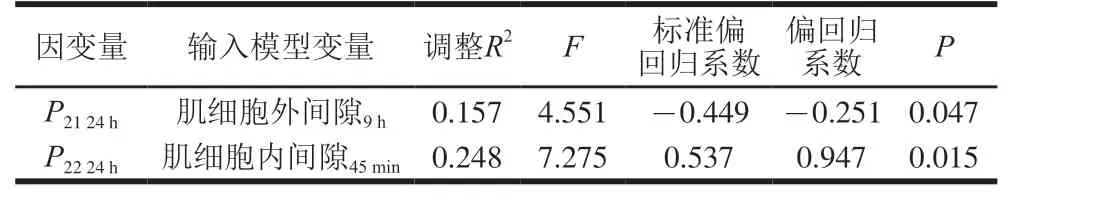
表5 肌细胞内外间隙解释宰后24 h的P21和P22变异的逐步回归模型Table 5 Stepwise regression models for explaining variation in P21 and P22with intra- and extracellular spaces of myocytes
肌细胞内间隙在宰后45 min和3 h与汁液流失率呈显著正相关(P<0.05,P<0.01),肌细胞外间隙在宰后9 h与汁液流失率呈显著正相关(P<0.05)(表4)。宰后45 min的肌细胞内间隙解释了汁液流失率变异的45.3%(表6)。说明宰后45 min的肌细胞内间隙对汁液流失率的贡献较大,且标准偏回归系数和相关系数都为正,表明宰后45 min的肌细胞内间隙越大,汁液流失率越高。
P2b在宰后12 h和24 h与汁液流失率均没有显著相关性(P>0.05),P21与汁液流失率呈极显著负相关(P<0.0 1),P22与汁液流失率呈显著正相关(P<0.05,P<0.01)(表4)。表明结合水与汁液流失率关系不大,不易流动水和自由水会对汁液流失率产生显著影响,这与王娟[17]、魏心如[31]和Bertram[32-33]等的研究结果一致。继续将宰后12 h和24 h时3 种状态水的相对含量与汁液流失率建立逐步回归模型(表6),结果只有宰后24 h的P22进入模型,解释了汁液流失率变异的39.5%,标准偏回归系数达0.655,相关系数为0.646。表明3 种状态水中自由水对汁液流失率的影响最大,自由水越多,汁液流失率越高。
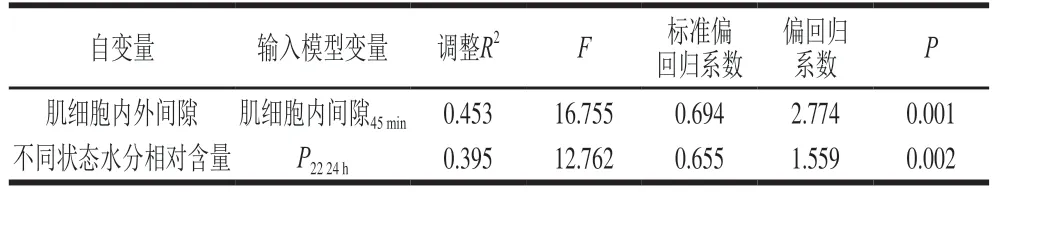
表6 肌细胞内外间隙和不同状态水分相对含量解释汁液流失率变异的逐步回归模型Table 6 Stepwise regression models for explaining variation in drip loss rate with intra- and extracellular spaces of myocytes and relative contents of different water states
3 结 论
生鲜猪肉在僵直过程中,肌原纤维的收缩会导致肌细胞内和肌细胞外都形成汁液流失通道。高汁液流失率样品的肌细胞内间隙形成较早,高、低汁液流失率组的肌细胞内间隙在宰后45 min已有明显差异,而肌细胞外间隙在宰后9 h才有差异。宰后45 min的肌细胞内间隙可以较好地解释自由水含量和汁液流失率的变异,宰后45 min的肌细胞内间隙越大,后期肌肉中自由水含量越多,汁液流失率越高,持水性越差。
猜你喜欢
食品科学(2022年17期)2022-09-30
青岛大学学报(医学版)(2020年6期)2020-11-16
河北农业大学学报(2020年5期)2020-10-30
食品工业科技(2020年7期)2020-05-05
食品研究与开发(2020年6期)2020-03-31
食品科学(2019年9期)2019-06-04
食品科学(2019年7期)2019-05-05
食品科学(2019年1期)2019-01-28
中国当代医药(2015年33期)2015-03-01
中国动物保健(2015年2期)2015-01-03
