
白菜型冬油菜NCED3基因、启动子的克隆及其表达分析
2020-11-21 09:44武军艳张亚宏马学才刘丽君牛早霞李学才李爱国孙万仓
干旱地区农业研究 2020年5期
武军艳,马 骊,方 彦,张亚宏,马学才,刘丽君,牛早霞,李学才,李爱国,孙万仓
(1. 甘肃农业大学农学院/甘肃省油菜工程技术研究中心,甘肃 兰州 730070;2. 甘肃天水市农业科学研究所,甘肃 天水 741001;3. 河北省农作物抗旱研究实验室,河北 衡水 053000)
白菜型冬油菜是一种新选育的强冬性油菜,可在极端低温为-20℃~-32℃的地区种植[1-2],它的成功选育使得我国北方地区农田冬季具有了覆盖作物,为北方耕作制度改革带来益处。在我国北方,冬油菜于8月中旬—9月上旬播种,次年3月中、下旬返青,5月下旬开始收获,收获后可复种其他作物(玉米、马铃薯、向日葵、荞麦、蔬菜等),具有显著的经济效益。其次,种植冬油菜可充分利用北方的光热资源,将北方传统的一年一熟模式转变为两年三熟模式[3],增加冬季土地覆盖,避免春耕,减少农田土壤表面灰尘[4],而农田表面灰尘是沙尘暴沙尘的重要来源。
脱落酸(Abscisic acid,ABA)与植物的多种抗逆性有关[5]。在高等植物中,ABA生物合成主要通过间接途径进行,在此合成途径中有多种酶参与[5],其中9-cis环氧类胡萝卜素双加氧酶(NCED)基因是关键的限速基因[6-7]。因此,在许多植物中NCED均被克隆[8-11]。拟南芥基因组中编码NCED的同源基因有9个,其中AtNCED3基因的表达可由干旱胁迫诱导[12];AtNCED3过表达可提高转基因植物对脱水胁迫的耐受性,这表明AtNCED3的诱导调节了ABA在脱水过程中的积累[13]。干旱胁迫后,烟草的NCED3基因表达上调,ABA含量增加[14]。关于NCED基因在不同胁迫条件下的表达调控及其作用的研究也有很多报道[15-16]。
研究发现,越冬前随着温度逐渐降低,白菜型冬油菜的叶片逐渐变黄,ABA含量增加[17],至越冬期叶片完全干枯,次年返青后自生长点重新长出心叶而完成生殖生长(图1)。因此,本研究克隆了白菜型冬油菜的NCED3基因及启动子序列,比较其与甘蓝型油菜的差异,并分析了不同胁迫处理条件下NCED3的表达特性,为了解NCED3基因的表达调控和NCED3启动子的结构特征,培育新的抗逆油菜品种奠定基础。
1 材料与方法
1.1 试验材料
选用2个抗寒性不同的白菜型冬油菜品种。陇油6号为超强抗寒性品种,可在我国北方极端低温为-20℃~-32℃的地区安全越冬,2010―2017年多点平均越冬率为90%;天油2号为强抗寒性品种,可在甘肃天水等地极端低温为-20℃的地区种植,2010―2017年多点平均越冬率为60%[1,3]。
选取饱满的上述油菜种子发芽,发芽后将材料播种于10 cm×10 cm的营养钵中,至幼苗生长至5片叶,置于低温培养箱中,4℃培养10 h后,采集叶片用于RNA的提取。
1.2 基因及启动子克隆
参照马骊等[18]方法提取RNA并合成 cDNA 第一链。根据GenBank 中公布的在甘蓝型油菜A3染色体上的NCED3 (登陆号:LOC103870025)基因的序列,在白菜型油菜基因组中寻找匹配序列,用匹配序列作模板,利用Primer 5.0软件设计引物(引物序列:ATGACTTCTTTCACGGCGACT,TTACACCTGATTCGCCAAGT)克隆基因。
1.3 序列分析
利用 NCBI 网站的软件BLAST 和 ORF finder进行核苷酸序列比较和开放阅读框分析。用Expasy Protparam 软件分析蛋白质的分子量、等电点和氨基酸组成及疏水性。用Conserved Domains分析蛋白质的结构功能域。蛋白质的二级结构使用在线工具 SOPMA 进行预测,三级结构使用在线软件Swiss-model 进行模拟。利用BLAST软件从NCBI的GenBank库中挑选十字花科植物的NCED3基因编码蛋白质的氨基酸序列,用DNAMAN8软件进行氨基酸序列多重比较,用MEGA软件分析基因同源性并构建系统进化树。启动子预测采用Plant CARE 及PLACE在线软件分析。
1.4 基因表达分析
当幼苗长至5片叶开始进行胁迫处理,分别为低温胁迫处理:将幼苗置于低温培养箱中于4℃培养;干旱胁迫处理:叶面喷施20% PEG-6000模拟干旱;ABA处理:叶面喷施100 μM ABA。于各处理后0(对照,CK)、2、4、8、12 h分别取幼苗相同部位的叶片及根系,液氮冻存后提取RNA用于基因表达分析。根据上述1.2节所得基因序列设计引物BN9DN,以油菜“看家基因”Actin作为内参,引物序列见表1。经普通PCR扩增后电泳检测其特异性,以浓度一致的陇油6号和天油2号叶片cDNA为模板,进行荧光定量PCR,目的基因与内参基因对应各3次重复,采用2-ΔΔCt方法计算[18]。
1.5 数据分析
采用Excel和SPSS进行数据的统计分析和作图。
2 结果与分析
2.1 NCED3序列分析
陇油6号NCED3的cDNA序列含有一个长度为1 794 bp的完整ORF,编码含597个氨基酸的蛋白质,属RPE65超家族,由20种氨基酸组成,其中以Leu、Ala所占比例最高,分别为8.0%、8.2%,相对分子质量约65.74 kD,理论等电点为5.81,总平均亲水指数为-0.293,表明该蛋白为亲水性蛋白;含有其蛋白家族保守结构域;具有1处最大疏水峰。蛋白质二级结构中含有 27.81% 的α螺旋、23.12% 的延伸链、9.72%的β折叠, 39.36%的无规则卷曲,α螺旋和无规则卷曲是NCED3二级结构的主要成分(图2)。
天油2号NCED3的cDNA序列含有一个长度为1,794 bp的完整ORF,编码含597个氨基酸的蛋白质,由20种氨基酸组成,其中以Leu、Ala所占比例最高,均为8.0%,相对分子质量约65.78 kD,理论等电点为5.94,总平均亲水指数为-0.318,表明该蛋白为亲水性蛋白;含有其蛋白家族保守结构域;具有1处最大疏水峰。蛋白质二级结构中含有25.46%的α螺旋、23.62%的延伸链、10.55%的β折叠,40.37%的无规则卷曲,无规则卷曲是NCED3二级结构的主要成分(图3)。

表1 PCR引物序列

图1 越冬前冬油菜叶片变黄(A),返青后心叶重新长出(B)Fig.1 Winter rapeseed leaves turn yellow (A) and then dry before wintering and grow new leaves (B) next year
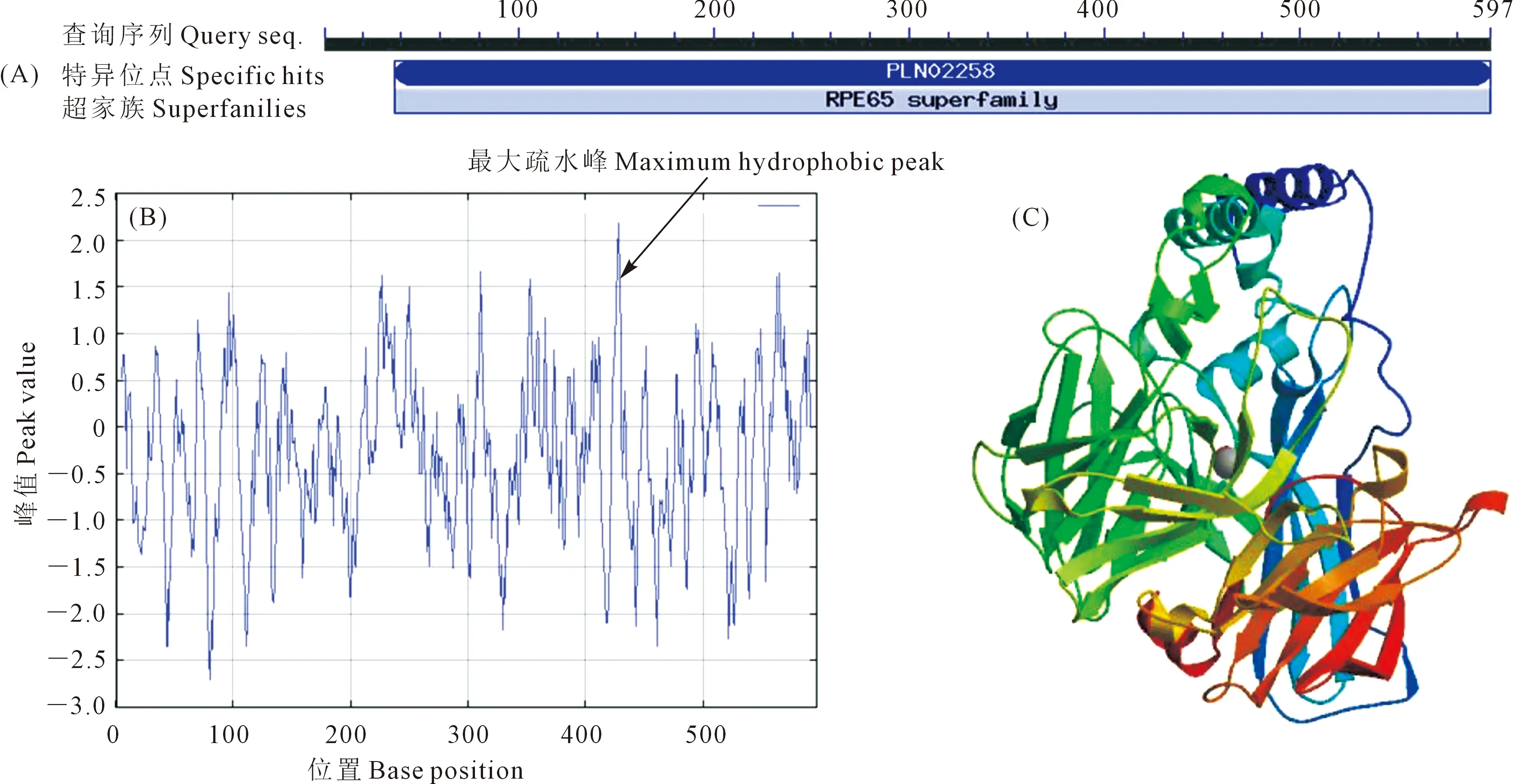
图2 陇油6号NCED3的结构域(A)、疏水性(B)及三级结构模型(C)Fig.2 Analysis of protein domain (A), hydrophobicity (B), and tertiary structure model (C) of NCED3 in Longyou 6
2.2 NCED3序列比对及进化发育分析
由图4可见,L6NCED3与BnNCED3蛋白相比,有5处发生氨基酸变化,相似度为99.16%,与BrNCED3有1处氨基酸不同。T2NCED3与BnNCED3蛋白相比,有6处发生氨基酸变化,相似度为98.99%,与BrNCED3有6处氨基酸不同。L6NCED3 与 T2NCED3有5处发生氨基酸变化,相似度为99.16%。对白菜型冬油菜及其他近缘种的NCED3进行同源性比较及系统进化分析(图5),陇油6号与白菜的同源性最高,相似度为99.83%,其次为甘蓝型油菜、天油2号。陇油6号与天油2号的相似度为99.16%。其中,陇油6号与白菜的相似度最高,天油2号与甘蓝型油菜的相似度最高。从进化关系分析(图5),所选的14个种与参试材料可被分为5类。陇油6号、白菜、甘蓝型油菜、天油2号及甘蓝的NCED3在同一分类中。可见NCED3蛋白比较保守,从而也可推断NCED3在白菜型冬油菜中的功能与甘蓝型油菜相似。
2.3 启动子序列分析
2个品种的启动子序列相似性为99.38%。启动子序列预测结果显示,启动子具有ABRE、TGA-element、CGTCA-motif、TGACG-element等激素响应元件(表2)。此外,还鉴定出低温响应元件(LTR)、防御和应激响应元件(TC-rich repeat)、生物过程中常见的顺式作用元件如CAAT-box等、分生组织表达CAT-box相关的顺式作用元件及- 30 TATA box附近的核心启动子元件等。可见,NCED3基因对不同的非生物胁迫有不同的响应。2个品种中只有1个不同的元件(circadian),该元件与昼夜节律控制有关。
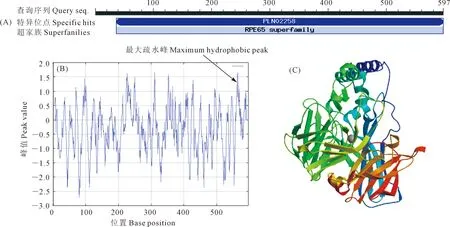
图3 天油2号NCED3的结构域(A)、疏水性(B)及三级结构模型(C)Fig.3 Analysis of protein domain (A), hydrophobicity (B), and tertiary structure model (C) of NCED3 in Tianyou 2

表2 启动子DNA顺式作用元件预测
2.4 NCED3表达分析
由图6可见,胁迫处理后NCED3在叶片和根中的表达均高于CK,且均呈上升趋势,于8 h达到峰值后略有下降,说明ABA、低温和干旱均可诱导NCED3基因表达。ABA处理后(图6A),NCED3在2个品种叶片和根中的表达均上调,其中天油2号的增长幅度高于陇油6号。低温处理后,基因表达与ABA处理呈现相似的表达趋势(图6B);陇油6号叶片在4 h时的表达水平低于CK,可能是胁迫下的强烈应激反应所致。PEG处理后,基因表达随处理时间延长呈逐渐增加的趋势,于8 h显著增高后降低(图6 C)。与根系的表达特性相比,3种处理后叶片的基因表达均在2 h显著升高,天油2号的升高幅度均大于陇油6号,可推断天油2号的叶片比陇油6号对胁迫的反应更快,胁迫后基因表达快速升高,合成积累大量的ABA来应对胁迫。
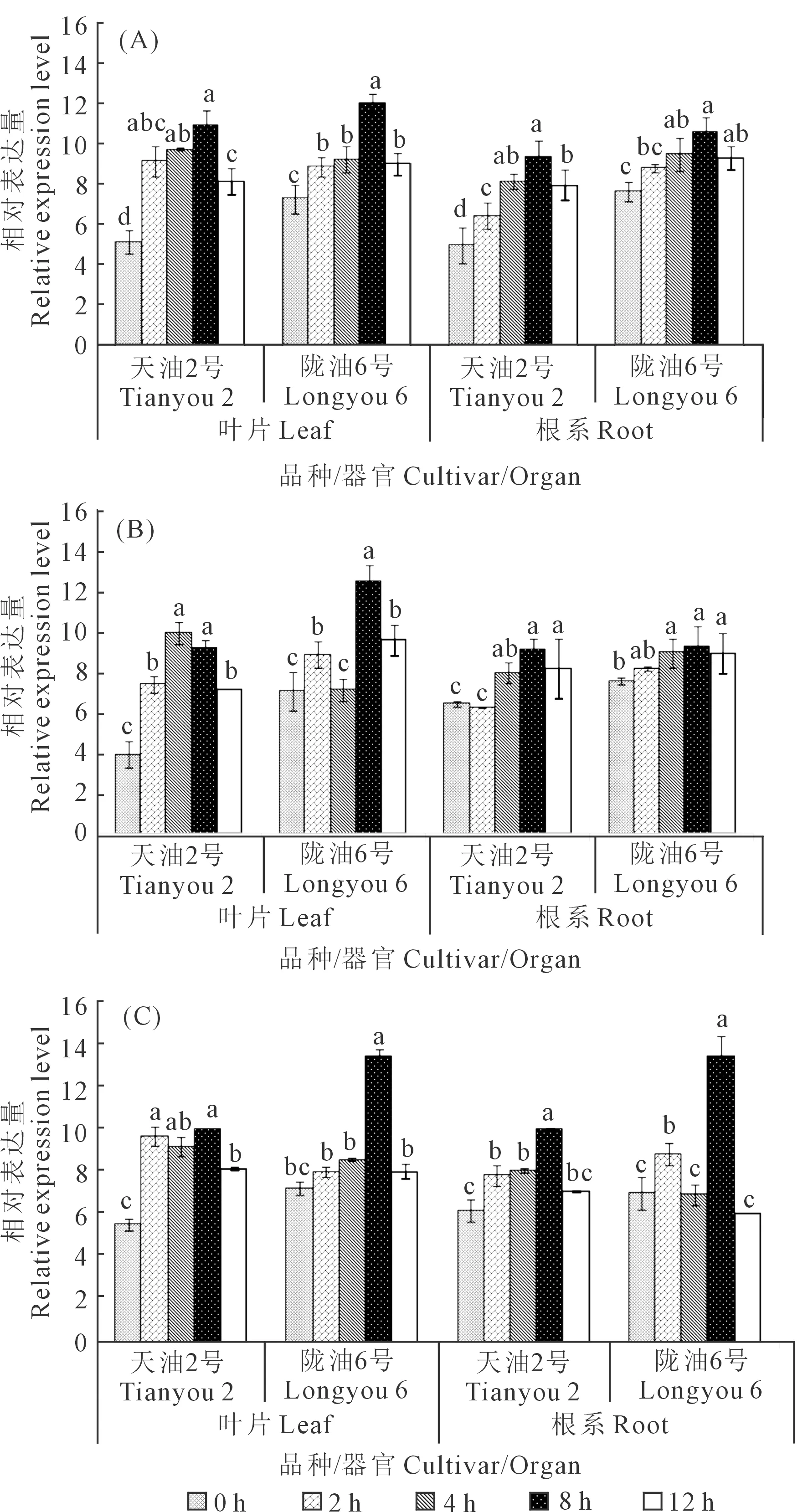
注:(A) ABA处理后的NCED3基因表达;(B) 低温(4℃)处理后NCED3基因表达;(C) PEG处理后NCED3基因表达。小写字母表示差异显著(P≤0.05)。Note: (A) NCED3 gene expression under ABA treatment; (B) NCED3 gene expression under low temperature (4℃) treatment; (C) NCED3 gene expression under PEG treatment. Different lowercase letters indicate significant differences (P≤0.05).图6 不同处理条件下NCED3表达分析Fig.6 The effects on NCED3 expression by different treatments
3 讨论与结论
基因结构的改变会导致基因表达的改变,拟南芥中AtNCED3的274和327位氨基酸的变化可能导致其表达的改变[19-20]。在本研究中,陇油6号和白菜的NCED3同源性较高,而天油2号与甘蓝型油菜的NCED3同源性较高;2个抗寒性不同的冬油菜NCED3相似度很高,且亲缘关系很近。可见NCED3蛋白比较保守,从而也可推断NCED3在白菜型冬油菜中的功能与甘蓝型油菜相似。
基因启动子中所含的顺式作用元件在基因表达调控中起着重要作用,基因的不同启动子特征决定了基因的不同表达特征[21-22]。根据预测结果2个品种的启动子相似性高,具有常见的与激素有关的元件如:ABRE、TGA-element、CGTCA-motif、TGACG-elemen等,可见NCED3基因的表达可能受到生长素、脱落酸、茉莉酸甲酯等的调控,本研究结果也表明ABA处理可诱导NCED3基因的表达。光可以参与植物的光合作用,也是一种非常重要的信号物质,从而调节基因转录[23-24]。本研究的启动子有3个与光相关的元件:G-box、TCT-motif和chs-CMA2a,它们通常存在于光诱导基因的启动子中;由此推断,NCED3的表达可能会受到光照的调控。在本研究结果中,白菜型冬油菜启动子中有1个特殊的元件,即低温响应元件(low-temperature responsiveness, LTR),该元件在其他被报道植物的NCED启动子中不存在[25-28]。因此推测,该基因可能受低温调控,本研究的基因表达分析结果也证实了该推测。本试验首次分析了低温(4℃)处理对NCED3表达的影响,低温可诱导白菜型冬油菜NCED3的表达。2个品种中有1个不同的DNA元件,即昼夜节律控制元件,它与季节反应的调节、从营养生长到生殖生长的转变以及休眠的开始等相关[29]。种子萌发、下胚轴伸长、叶片运动、气孔导度、开花、衰老等发育过程均通过昼夜节律钟进行调节[30]。根据形态学观察,天油2号的下胚轴长度确实比陇油6号的长。因此,如果与昼夜节律控制因素有关,这种差异是否与2个品种的抗寒性有关,可以作为我们今后研究的方向。
本研究中,NCED3基因可以在冬油菜的根和叶中表达,而没有组织特异性,这与拟南芥的研究结果相似[31]。经过低温、PEG模拟干旱和ABA处理后,NCED3在陇油6号和天油2号中的表达量均增加。已有研究表明,NCED3通过调控ABA的合成,在植物耐旱性方面发挥重要作用[32]。本研究中干旱胁迫可诱导NCED3的表达,12 h前表达量急剧增加,12 h后下降,这与Xian等[32]和Zhang等[11]的研究结果基本一致。本研究中,在低温、模拟干旱及喷施ABA处理中,随着处理时间延长,天油2号的NCED3基因表达增加幅度均高于陇油6号(包括叶片和根),由此可见,逆境胁迫后弱抗寒品种比强抗寒品种更敏感,胁迫初期快速做出反应,调控基因表达来应对逆境,该结论与Zeng等[33]的结果相似。
猜你喜欢
今日农业(2022年1期)2022-11-16
电脑迷(2022年10期)2022-11-08
农业工程学报(2022年5期)2022-06-22
趣味(作文与阅读)(2022年3期)2022-06-10
今日农业(2022年4期)2022-06-01
小读者(2021年6期)2021-11-23
小猕猴智力画刊(2021年9期)2021-10-11
农技服务(2021年6期)2021-09-22
中学生数理化·中考版(2016年7期)2016-12-07
山东工业技术(2016年15期)2016-12-01
