
纳塔栎和柳叶栎对铅锌矿区污染土壤的修复潜力分析:田间试验
2020-11-14 07:07曲豪杰张涵丹谭志超陈光才
生态学报 2020年19期
曲豪杰,张涵丹,谭志超,陈光才,*
1 中国林业科学研究院亚热带林业研究所, 杭州 311400 2 南京林业大学, 南京 210000 3 临武县众诚林木种植专业合作社, 郴州 423000
我国是全球矿产资源消耗大国,对矿产资源的需求和实际开采力度也在逐年增加。开采矿产资源过程中会直接对周围环境造成水土流失、植被减少和土壤地质结构被破坏等不利影响[1]。金属冶炼及采矿废水、废渣的随意排放等也会致使大量重金属进入到土壤,造成矿区周边土壤重金属超标[2-3]。根据2014年《全国土壤污染状况调查公报》,与1980年代末相比,25年间我国土壤重金属浓度明显增加,以Cd为例,中国西部和北部的平均浓度增加了10%—30%,沿海地区和西南地区土壤Cd增加了50%以上[2]。土壤重金属污染会导致土壤质量下降[3]、农作物减产[4]、农产品品质降低[5],甚至威胁到人类健康和生态安全,给社会的可持续发展造成严重威胁[6]。
植物修复是利用植物对污染物的积累能力进行环境修复的方法,具有新颖、经济、高效、环保、原位修复等特点,被认为是一种绿色的重金属污染修复方式[7-8],适合大面积污染土地的修复。利用超积累植物对重金属的高积累及转运能力将重金属积累在地上部分的植物萃取技术、以及利用高耐性植物将重金属积累在根系从而降低其移动性的植物稳定技术,是植物修复重金属污染土壤的主要方式[9-10]。植物稳定对土壤的修复作用主要表现为两个方面:一方面是植物根部对重金属离子的积累、吸附和转化,从而固定土壤中的重金属;另一方面保护污染土壤不受或减少侵蚀,通过减少水土流失减少重金属在地表的流失、及在土壤垂直方向的下渗和迁移[11]。
近年来,有研究发现与草本的超积累植物相比,木本植物根系生物量、覆盖面积、养分利用率和生长周期均高于草本植物[12]。木本植物寿命长,一般不进入食物链、可以持续对重金属土壤进行修复[13],在重金属重度污染土壤修复中具有较大潜力。例如,研究发现柳树(Salixspp.)、墨西哥落羽杉(TaxodiummucronatumTenore)、水紫树(Nyssaaquaticre)、枫香(LiquidambarformosanaHance)、紫穗槐(AmorphafruticosaLinn)等树种对重污染土壤中Cd、Pb和Cu的耐性强,能够维持较强的生长势头[14-15]。针对重金属污染程度超过风险管控值的严格管控用途类土地,国家要求强制退耕还林、还草。《土壤污染防治行动计划》要求,到2020年全国重度污染耕地种植结构调整或退耕还林还草面积力争达到133万hm2。然而,由于重金属对植物的毒害作用较强,并且矿区土层破坏严重,多为砂石、石砾等质地,保水保肥能力差,土壤极度贫瘠,使得植物修复较难在尾矿地区实践[16]。因此,筛选、利用对重金属具有较强耐性、景观效果好、木材价值高的景观或用材树种,在实现重金属污染修复的同时,可以形成较好的森林景观,实现重度污染土壤的安全利用目标,潜力巨大,需求迫切。
橡树,又称为栎树,其树体雄伟,冠大荫浓,材质优良,生态适应性强,在园林绿化、生态建设与木材生产中具有重要潜力。二十世纪初期,我国从国外引进了一批优良栎树资源如纳塔栎(Quercusnuttallii)、柳叶栎(Quercusphellos)、水栎(Quercus.nigra)、舒玛栎(Quercus.shumardii)等,在长江中下游地区表现出适应性强、生长快、景观效益好的优良性状,应用范围逐步扩大[17-19]。Evangelou 等[20]发现在Cu、Zn、Cd和Pb污染的土壤中,栎树(Quercusrobur)比柳树(SalixviminalisL.)、杨树(Populusmonviso)、桦树(Betulapendula)有更高的重金属耐性。Shi等通过盆栽实验,发现柳叶栎、舒马栎、弗栎(Quercus.Virginiana)在Pb、Zn尾矿矿砂及污染土壤中生长较好,显示出较强的重金属耐性[21]。那么,这些栎树应用于矿区重金属污染土壤的植被恢复及生态修复实际表现如何?目前对此尚无相关研究,一定程度上限制了栎树在重金属污染地生态修复工程的实际应用。针对以上重金属污染生态修复的实际需求,本研究旨在(1)分析评价纳塔栎和柳叶栎在矿区污染土壤中的生长表现及重金属植物修复潜力;(2)筛选矿区污染土壤植被恢复的耐性乔木树种,为金属矿区污染地的植被恢复和生态修复提供科学依据。
本研究选择湖南某Pb、Zn矿区重金属污染的废弃地,利用树型优美、叶色在秋末分别变为红色和黄色的纳塔栎和柳叶栎进行了植被恢复试验,生长1年后对纳塔栎和柳叶栎开展生长调查,测定树苗的树高和地径,采集植物样品和矿区土壤,分析树木组织、根际土壤中的重金属含量,为评价纳塔栎和柳叶栎在矿区污染土壤中植被恢复及生态修复潜力提供田间试验依据。
1 材料与方法
1.1 试验区概况
试验地位于湖南郴州市临武县(北纬25°31′3.60″ 东经112°33′7.66″),地处湖南省最南部,与蓝山县、嘉禾县、桂阳县、北湖区相接;属中低纬度区,气候温和,雨量充沛,光热充足,年平均气温为17.9°C,年降雨量为1022.3—1917.4 mm。临武矿产资源丰富,探明的矿物有9类32种,大型矿床9处,中型矿床12处。
1.2 试验设计及采样调查
该研究主要在临武典型的Pb、Zn矿区废弃地开展,通过人工引种造林开展植被恢复。于春季在A区(尾矿库)、B区(苗圃)、E区(山地)种植1年生纳塔栎,在C区(苗圃)、D区(山地)种植1年生柳叶栎(图1)。自然生长至2018年6月树龄达2年。苗圃(B、C区)种植面积为33.33 hm2,各植株株行距为1 m×0.5 m左右;山地(D、E区)的种植面积为66.67 hm2,各植株间距为2 m×2 m;尾矿库(A区)种植面积为50 m2,各植株间距为2 m×2 m。分别在各区域随机选取长势相近植株4株,用卷尺垂直测量其地上部最大高度,用游标卡尺测量植株地径(测量点距地面10 cm处)。之后树木整株挖出,并尽量保持根部完整性。抖动植物根部采集各植物根部土壤1份,铲除尾矿库表层土层挖取深层矿砂3份,每份土壤及矿砂样品约500 g。将采集的植物样品分成根茎叶,装入塑封袋,用带有冰袋的保温箱保存,土壤样品放入塑封袋保存。
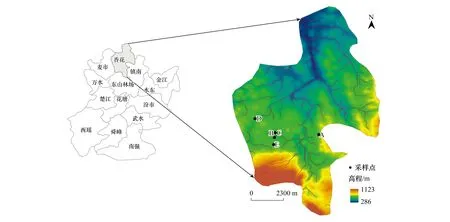
图1 采样点分布图Fig.1 Sampling point distributionA,A区种植纳塔栎;B,B区种植纳塔栎;C,C区种植柳叶栎;D,D区种植柳叶栎;E,E区种植纳塔栎
1.3 样品处理与测定
将样品带回实验室后,首先用自来水冲洗以去除粘附于植物样品上的泥土和污物,然后用纯水冲洗,沥干并于105 ℃杀青之后,在70 ℃下烘干至恒重。植物样品剪碎后,用球磨机(Retsch MM400,GER)磨碎,过0.149 mm筛,用来测定重金属含量;土壤样品自然风干,拣出砖头、石块、杂草等,用木棒碾碎分别过2、0.149 mm筛,前者用于土壤pH和重金属(Cd、Ni、Pb、Cu、Zn、Cr、As)有效态的测定,后者用于重金属全量的测定。
土壤样品采用王水(盐酸∶硝酸为3∶1)进行消解,植物样品用硝酸-双氧水(硝酸∶双氧水=5∶1)消解,经消解后土壤和植物样品用电感耦合等离子体质谱仪(ICP-MS,Perkin elemer NexION300D,USA)和电感耦合等离子体发射光谱仪(ICP-AES,ThermoFisher CAP 7400,Germany)测定重金属(Cd、Ni、Pb、Cu、Zn、Cr、As)含量。
土壤重金属有效态浸提:采用DTPA浸提(其成分为0.005 mol/L DTPA-0.01 mol/L CaCl2-0.1 mol/L TEA),浸提后用电感耦合等离子体质谱仪(ICP-MS,Perkin elemer NexION300D,USA)和电感耦合等离子体发射光谱仪(ICP-AES,ThermoFisher CAP 7400,Germany)测定重金属(Cd、Ni、Pb、Cu、Zn、Cr、As)有效态含量。
采用2.5∶1水土比浸提 — pH计法测定土壤样品的pH值。
1.4 土壤污染评价
1.4.1单项污染指数法
单项污染指数法是评价土壤污染程度的无量纲指数,可反应污染物超标倍数和污染程度[22]。其计算公式:
式中,Pi为土壤中重金属i的单项污染指数;Ci为土壤中重金属i的实测值;Si为依据《食用农产品产地土壤环境质量标准限值》(HJ 332-2006),当Pi≤0.7 时,污染等级为1,表示土壤清洁;0. 7
1.4.2综合污染指数法
内梅罗指数法(Nemerow index) 是常用的综合污染指数法之一, 能够较全面地评价整个区域土壤重金属的污染程度[22]。其计算公式如下:
式中,PN为某样点(或某区域)所有重金属的内梅罗综合污染指数;Pmax为土壤重金属最大单项污染指数;Pave为所有重金属单项污染指数平均值。根据综合污染指数等级标准,当PN≤0. 7时,污染等级为1,表示土壤清洁; 0.7
1.5 植物修复能力
通过计算生物富集系数(BCF)和转移系数(TF)可以定量评价植物修复能力。生物富集系数表明植物从周围环境中积累金属到组织中的效率[7],公式如下:
式中,Charvestedtissue为目标部位重金属含量;Csoil为土壤重金属含量;
转移系数是指植物将积累的金属从根转移到地上部的效率[7],计算方法如下:
式中,Cshoot为地上部重金属含量;Croot为根部重金属含量。
1.6 数据处理
采用IBM SPSS Statistics 19进行数据处理及单因素方差分析,采用ArcMap 10.6、OriginPro 9.1和R 3.6.1 进行作图。
2 结果与分析
2.1 试验区土壤污染评价
由表1所示,矿区土壤偏酸性,土壤pH值的范围在4.39—6.13之间,矿砂的pH值为2.78。不同区域土壤重金属含量具有一定的差别,矿区中Ni、Cu、Cr含量基本处于农用地土壤污染筛选值(《农用地土壤污染风险管控标准》GB 15618—2018)范围内,Cd、Pb、Zn、As是区域的主要污染物。其中矿砂、A区、B区、C区、D区土壤中Cd、Pb、Zn、As均超出污染筛选值。矿砂中Cd、Pb、Zn和As的含量极高,分别达到了10.82、3336.91、876.08 mg/kg和34498.14 mg/kg,超出筛选值36.07倍、47.67倍、4.38倍、862.45倍。A区土壤是直接受到矿砂污染的其Cd、Pb、Zn和As分别为3.61、317.77、498.37 mg/kg和1101.11 mg/kg。B区、C区、D区主要污染物浓度低于矿砂和A区。E区污染相对较轻,仅Pb、As浓度较高,超出土壤污染筛选值1.19倍和1.3倍。
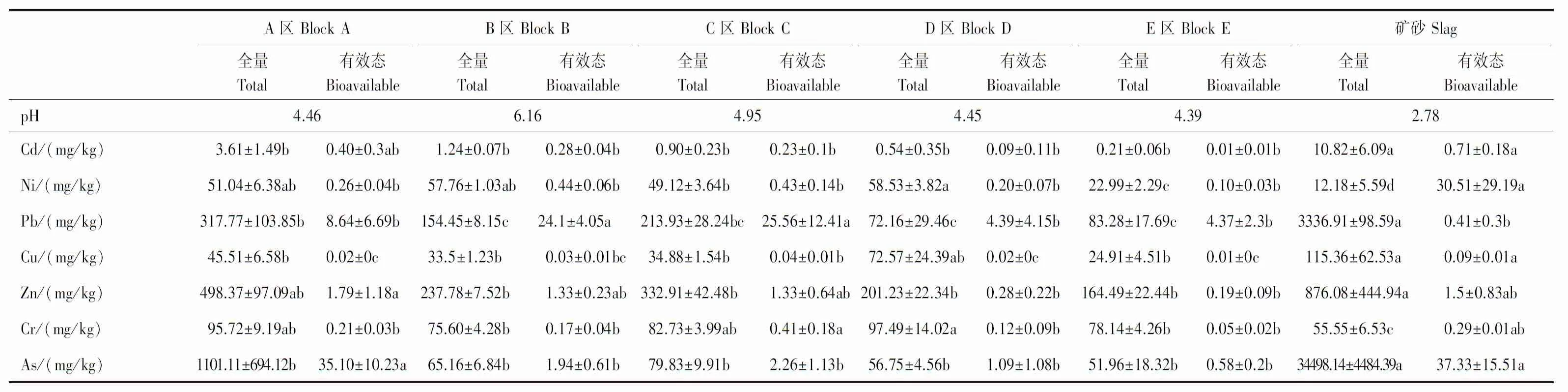
表1 土壤重金属含量/(mg/kg)Table 1 Heavy metals level in soil
单项污染指数(表2)表明,矿砂中Cd、Pb、Zn、As的Pi值均远超过3,达到重度污染水平;A区域中Cd、Pb、As的Pi值> 3为严重污染,Zn的Pi值在2—3之间,表现为中度污染,Ni的Pi值在1—2之间表现为轻度污染;B区Cd的Pi值为4.13,达到了重度污染水平,Ni、Pb、Zn、As的Pi值在1—2之间为轻度污染;C区的Cd、Pb的Pi值在2—3之间,达到中度污染水平,Ni、Zn、As的Pi值在1—2之间为轻度污染;D区的Cd、Ni、Cu、Zn、As的Pi值在1—2之间均表现为轻度污染;E区Pb和As表现为轻度污染。单项污染指数说明不同区域单一重金属的污染程度差异较大,难以评判复合污染的程度。因此,我们进一步计算了综合污染指数,表明矿砂存在严重的复合污染,数值PN为617.31远超重度污染参照值3,A区和B区也达到了重度污染水平,PN值分别为20.08和3.14,C区PN值为2.43处于中度污染水平,D区和E区PN值分别为1.55和1.07,达到轻度污染水平,污染程度由重到轻依次为:矿砂>A区(尾矿库)>B区> C区>D区>E区。
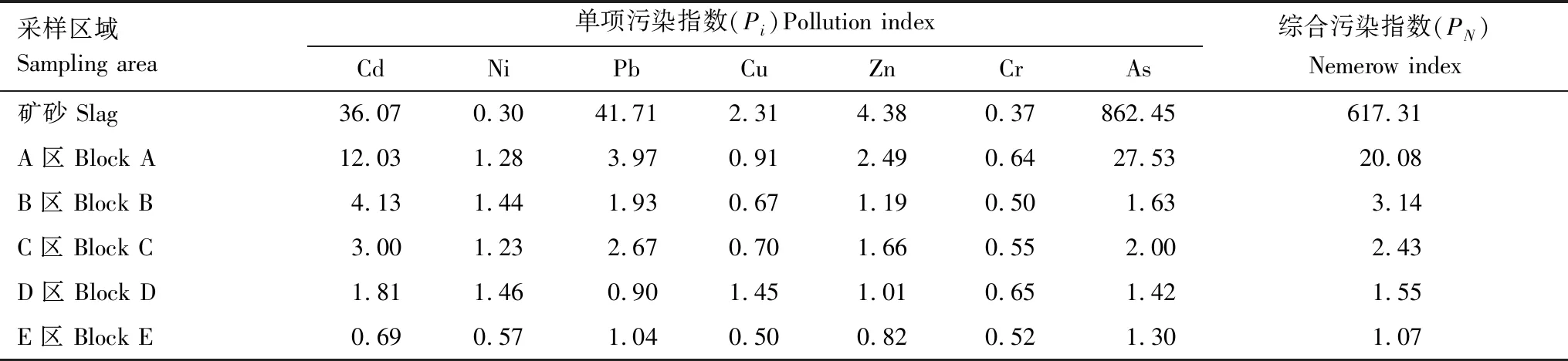
表2 土壤重金属综合污染指数Table 2 Comprehensive pollution index of heavy metals in soil of mining area
2.2 树木生长分析
分别统计各区域植株的成活率,测量采集植株的树高、地径和不同部位的生物量分析各区域栎树的生长情况。各区域内植株能很好存活,成活率接近100%,并且生长较为正常,表现出较强的重金属耐性。不同区域植株的树高、地径和不同部位的生物量(表3)不同,植株的地径、树高以及生物量均随污染的加重呈减少的趋势。五个区域树木地径在12.28—21.32 cm之间,树高在0.93—1.61 m之间,生物量由大到小的顺序为E区纳塔栎、D区柳叶栎、C区柳叶栎、A区纳塔栎、B区纳塔栎,依次为503.7、336.94、239.67、132.95、124.67 g。除A区纳塔栎和B区纳塔栎,其余各区域植物间生物量有显著的差异。根、茎、叶生物量占比表明,A区纳塔栎叶子的生物量占比仅有其整株植株生物量的5%,其他不同区域的叶子生物量占比在15%—35%之间。
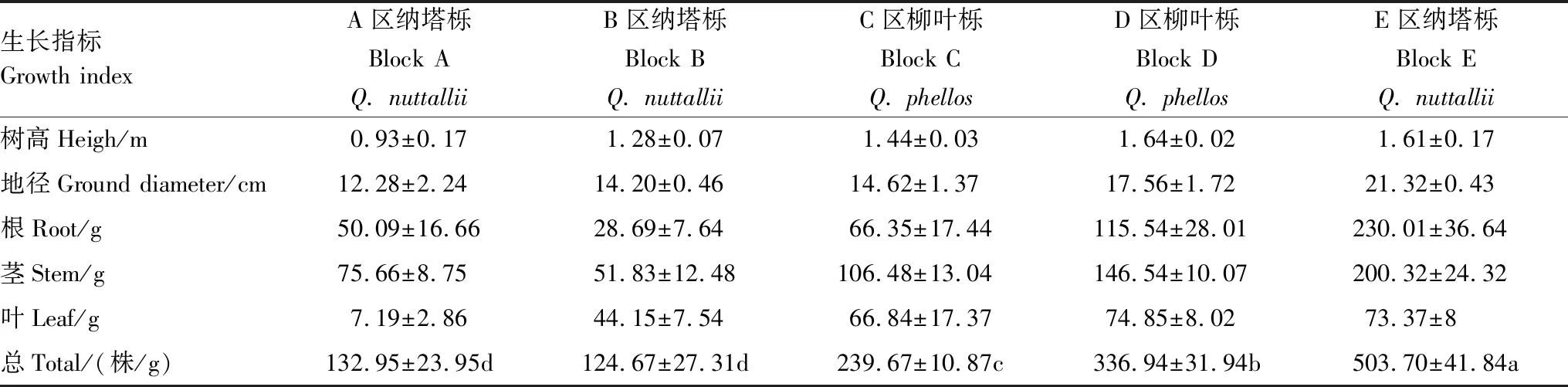
表3 不同区域植物树高、地径及生物量Table 3 Tree height、ground diameter (DG) and plant biomass in different areas
2.3 树木的重金属含量和单株积累量
树木组织内重金属浓度、分布模式因立地土壤的重金属水平和种类而不同。图2表明,纳塔栎和柳叶栎体内Zn的含量高于其他重金属;且在Zn轻度污染水平下(B—E区),柳叶栎和纳塔栎均表现出叶片Zn浓度(39.18—59.95 mg/kg)显著高于茎和根的浓度(10.04—23.39 mg/kg)(P<0.05),但在Zn中度污染的尾矿库中(A区),根中Zn的浓度最高为300.92 mg/kg,且与茎和叶中没有显著性差异。A、B、E区纳塔栎体内Cd的含量在4.36—26.91 mg/kg,C、D区柳叶栎体内Cd含量为0.74—1.29 mg/kg,其在根茎叶中的分布没有显著性差异。Pb在A区纳塔栎中含量最高,其在根中浓度达到了47.44 mg/kg,其余区域的纳塔栎和柳叶栎Pb含量在2.18—19.17 mg/kg之间,各区域Pb在根中的浓度均高于茎和叶。A区As为严重污染,其纳塔栎的根中浓度达到了64.38 mg/kg,显著高于茎(5.16 mg/kg)和叶(10.29 mg/kg)(P<0.05)。其余轻度污染区域As含量则在0.52—1.55 mg/kg之间。
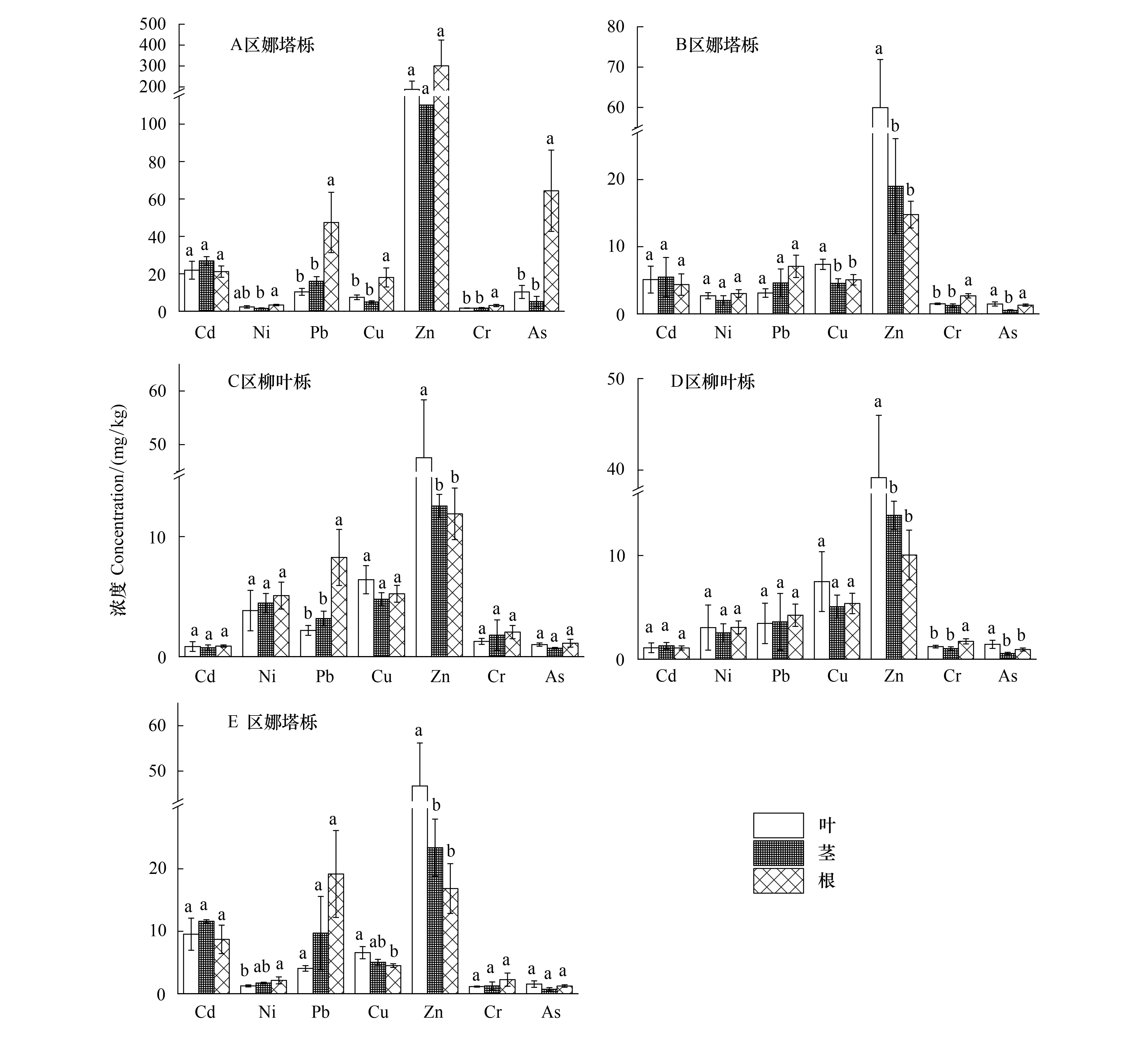
图2 植物体内重金属浓度Fig.2 Concentrations of heavy metals in plants不同小写字母代表同种金属不同部位差异显著(P<0.05)
通过综合树木的干生物量和重金属含量,进一步计算纳塔栎和柳叶栎单株的金属积累量(表4)。纳塔栎在复合污染程度最低的E区,具有最高的生物量,且对Cd、Pb、Cu、Cr的积累量最高,分别为5.01、6.48、2.53、0.86 mg/株;在污染程度最高的A区,Cd、Pb、Cu、Zn、As的积累量分别为3.24、3.77、1.39、3.6 mg/株,其中Zn和As的积累量高于其他区域,Cd、Pb、Cu积累量仅次于B区纳塔栎。同样,复合污染程度较低的D区柳叶栎对不同重金属的积累量均超过C区。从不同重金属元素间的积累量看,柳叶栎和纳塔栎对Zn的积累量最高。不同污染程度下纳塔栎对Cd积累量要高于柳叶栎。
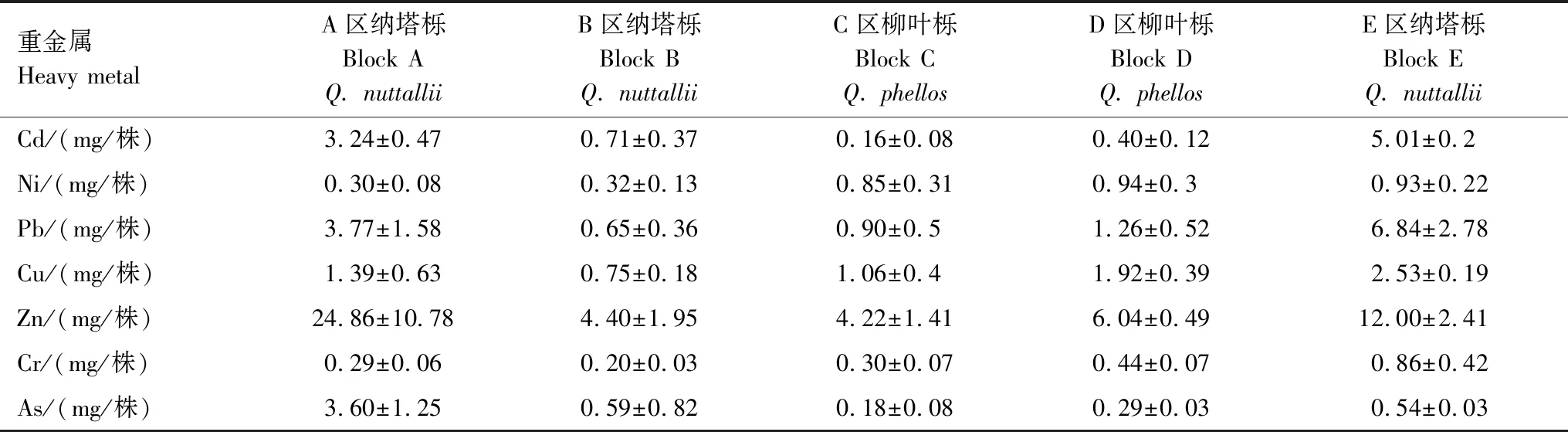
表4 不同区域植物重金属积累量Table 4 Accumulation of heavy metals in plants in different regions
2.4 树木对重金属的富集和转运
纳塔栎和柳叶栎对不同重金属的生物富集系数(BCF)(图3)和转运系数(TF)(表5)不同,并受到土壤重金属浓度的影响。纳塔栎和柳叶栎对Cd有较强的富集能力,其富集系数显著高于其他重金属(P<0.05)。A、B、E区纳塔栎Cd的BCF值分别为6.27—8.37、3.67—4.38、42.93—52.75,显著高于C、D区柳叶栎的值1.79—2.15、0.89—1.07(P<0.05)。值得注意的是E区纳塔栎在土壤Cd浓度为0.21 mg/kg时,植物体内不同部位Cd浓度为8.7—11.6 mg/kg,其BCF值也达到42.93—52.75。各区域的纳塔栎和柳叶栎对金属Ni、Pb、Cu、Zn、Cr的BCF值在0.01—0.59之间各不相同,其中最高值为A区纳塔栎叶对Zn的BCF值,达0.59,而对Ni、Cr、As的富集系数(BCF)均不足0.1,对Pb、Cu的富集系数(BCF)也均低于0.3。
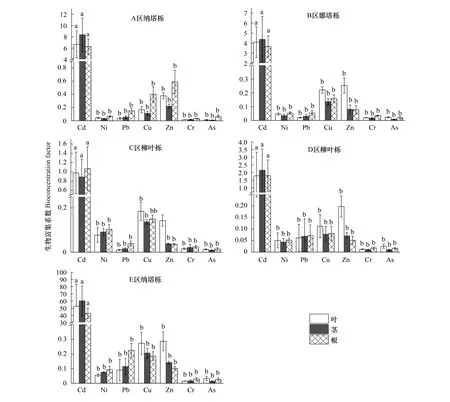
图3 植物不同部位生物富集系数Fig.3 Bioconcentration factor of different parts of plants不同小写字母代同一区域不同重金属BCF差异显著(P<0.05)
结合转运系数TF值(表5)可以发现,在Zn轻度污染区域(B—E区),Zn的转运系数较高,在1.79—2.28之间,但是在Zn中度污染以及复合污染严重的A区Zn的转运系数仅有0.43。植株对Cd的转运系数值在0.89—1.34之间,其中最高值出现在E区的纳塔栎中(1.34),C区柳叶栎最低为0.89,在复合污染严重的A区,Cd的转运系数为1.28。各区域植株对Pb的转运系数在0.35—0.8之间。A区纳塔栎Ni的转运系数为0.5,B—E区中两个树种Ni的转运系数在0.78—0.85之间。
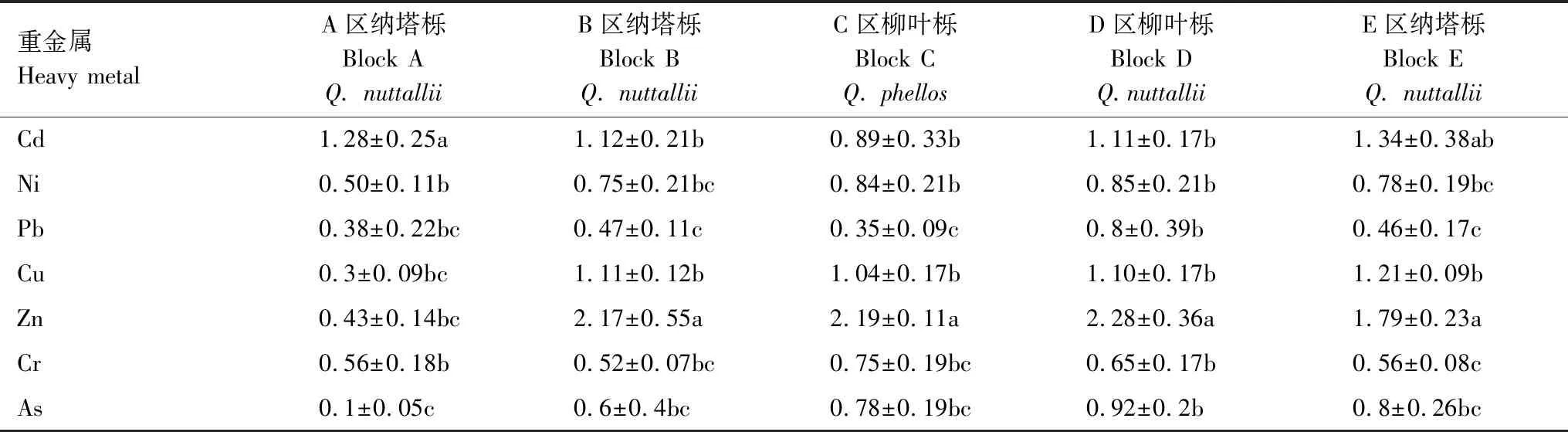
表5 不同区域树木对重金属的转运系数Table 5 Translocation factor of heavy metals in trees grown in different regions
3 讨论
3.1 树木对重金属胁迫的适应性
本研究区域是Pb、Zn矿区,存在多种金属复合污染,由图4可知重金属Cd、Pb、Zn、As的浓度与PN有很强的正相关性,除Pb外均达到显著性水平(P<0.05),相关系数分别为0.98、0.86、0.91和1。同时各重金属的单项污染指数Pi值(表2)和土壤重金属含量(表1)均表明主要污染为Cd、Pb、Zn、As四种重金属。纳塔栎和柳叶栎在轻微污染、中度污染的土壤都生长较好,但是随着污染加重,两种栎树的生长受到了一定程度的抑制。不同区域的生物量大小为B区纳塔栎
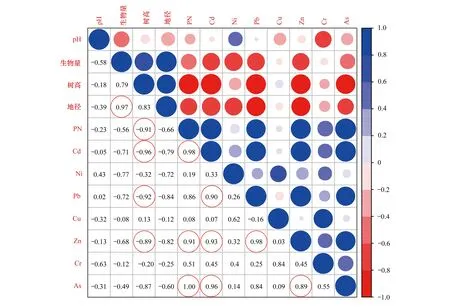
图4 pH、土壤重金属、综合污染指数(PN)与植物生长的关系Fig.4 Correlations between pH,metal(loid)s,PN and Plant growth state红色圆圈标记代表P<0.05
近年来,研究者在不同的土壤条件下开展了树木的适应性研究,施翔等人发现紫穗槐、桤木(Alnuscremastogyne)和黄连木(Pistaciachinensis)能在Pb、Zn矿砂和Cu矿砂中生长,三种植物生物富集系数(BCF)值和 转移系数(TF)值都小于1[16]。在Cu、Pb和Zn含量分别为467.45、49.69 mg/kg和656.63 mg/kg的复合污染土壤中,白蜡树(Fraxinuschinensis) 、白棠子树(Callicarpadichotoma) 、紫薇(Lagerstroemiaindica) 、盐肤木(Rhuschinensis) 、接骨木(Sambucuswilliamsii) 、刺槐(Robiniapseudoacacia)、枫香树、构树(Broussonetiapapyrifera) 、山桐子(Idesiapolycarpa) 和珊瑚树(Viburnumodoratissimum) 表现出较好的耐性[13]。同样,本研究各区域中纳塔栎和柳叶栎也维持了较为正常的生长,保持了较高的生物量,表现出较强的重金属耐性。表明纳塔栎和柳叶栎对Cd、Pb、Zn、As复合污染土壤表现出较强的适应性。利用两种栎树对复合污染的高耐性对重污染矿区进行植被恢复,可以起到恢复矿区植被破坏,减少裸露矿区水土流失的效果,进而起到降低重金属随水分迁移的能力,从而减少其对生物和环境的危害,同时满足植物稳定和美化景观的效果。
3.2 重金属在树木中的累积
李俊凯等[26]认为植株体内重金属浓度与土壤重金属浓度具有一致性,比较各区域土壤重金属(表1)、植株体内重金属的浓度(图2)以及积累量(表4)可以发现:A区纳塔栎土壤重金属浓度最高,同样A区纳塔栎体内的重金属(Cd、Pb、Zn、As)含量最高,对Zn、As的积累量也最高,C、D区柳叶栎体内的Pb、Zn、As含量具有相同的规律。但是随着土壤污染水平的降低E区和B区纳塔栎体内重金属(Cd、Pb、Zn、As)浓度却与土壤污染趋势相反;同样柳叶栎中Cd也表现与土壤污染程度相反的趋势。其中Cd、Pb、Cu的积累量均为污染程度最低的E区纳塔栎最高且污染较轻的D区柳叶栎对各种重金属的积累量均大于C区柳叶栎。表明在污染加重情况下植物体中重金属浓度会相应增加,但这同时也存在抑制植株的生长从而反过来影响植物对污染物的吸收的情况。有研究发现杞柳微山湖(S.integra‘Weishanhu′)在Cd胁迫试验中,随着Cd浓度的增加根系对Cd的积累量达到最大值后便开始下降[27]。土壤污染会一定程度提高植株体内含量,但随着污染的加重,会影响植株的生长,降低植株的生物量,最终影响植株对重金属的总积累量,因此复合污染低可能有利于部分重金属的积累。另外,植物对重金属的富集能力除了受土壤重金属含量影响,其他因素如植物特性、土壤理化性质、土壤中重金属的浓度、形态和毒性大小等也会影响重金属的积累[14]。
3.3 纳塔栎和柳叶栎对重金属的富集和转运
纳塔栎和柳叶栎两个树种对Cd的生物富集能力(BCF)显著高于其他重金属(P<0.05)(图3),且不同区域纳塔栎和柳叶栎对Cd的转运系数(TF)(表5)在0.89—1.34间。比较两个树种可以发现纳塔栎对Cd生物富集能力和转运能力要高于柳叶栎。在Cd污染区域和不含Cd污染区域,纳塔栎对 Cd的生物富集系数均高于其他重金属,转运系数(TF)在1.21—1.34间。原因可能是Cd作为植物生长发育非必需的金属元素,由于它存在着强毒性与可迁移性,所以被植物的根系所吸收的Cd很容易迁移至植物的地上部[28]。也有研究发现低浓度的Cd胁迫处理植物能够促进其生长,且促进作用会因植物物种的不同而有所差异[29-30],因此在轻污染区域纳塔栎和柳叶栎对Cd仍具有较高的富集和转运能力。复合污染严重的A区纳塔栎对Zn的生物富集系数高于其他区域,但Zn的转运系数TF却明显低于其他区域的植株。原因是随着土壤Zn浓度的增加植物体内也相应的增加,因此植物体的富集量增多,Zn轻微污染的情况下柳叶栎和纳塔栎会主动向地上部转运Zn,但当土壤中Zn浓度达到中度污染以上时,Zn在植株根部的浓度不断增加,最终影响植株生长且转运系统受到抑制。这与齐笑笑等的研究相一致,当Zn含量低时,根系可优先满足地上部需要而向上转运,而当Zn供应充足时,多余的Zn大多富集在根部[31]。各区域树木对Pb的富集系数在0.02—0.22之间且根部高于茎和叶除D区柳叶栎外Pb的转运系数均低于0.5,这也与Pb在植株体内的浓度相一致。对大多数非耐性或非超积累植物而言, 根系所吸收的Pb大部分被局限于根系组织(比例大约为95% 或更高),仅有少部分铅可借助共质体途径向地上部输送并累积[32]。各区域As的生物富集系数均少于0.07,As与Fe、Mn、Zn和Cu等植物正常生长及代谢所必需的元素不同,对植物而言是一种非必需元素且没有生理功能。植物吸收As首先进入细胞间隙以及细胞壁间的间隙(即质外体中)进行运输, 但由于根部内皮层上凯氏带(casparian strip)的阻隔,As不能通过质外体途径直接到达木质部导管,而必须经过共质体途径进行跨质膜转运和木质部装载且不同形态的砷在跨质膜转运过程中需要不同的质膜转运系统和转运蛋白,这可能限制了非超积累植物对As的吸收[33]。Ni、Cu、Cr的生物富集系数均在0.3以下,三者在土壤中,即三者在土壤中基本不存在污染,因此植物富集较少。Ni、Cu、As的转运系数与Zn有相似的规律,A区显著低于其他区域,不同的是A区Cu、Ni污染并不严重,因此可能是复合污染影响植物的生长发育进而影响到对其他重金属的转运能力。过量的重金属可以降低植物根系活力,影响植物生长,影响土壤微生物活性、土壤酶活性等进而影响植物对其他重金属的吸收转运[34]。
4 结论
纳塔栎和柳叶栎对Cd、Pb、Zn、As复合污染土壤均具有一定的耐性,两个树种对Cd具有较高的生物富集能力,能够在组织中富集比土壤含量更高的Cd;对Zn具有较高的转运能力,能够从地下部向地上部转更多的Zn。在Cd、Pb、Zn、As不同污染水平下,纳塔栎相较于柳叶栎表现出更优的耐性和重金属富集能力,对Cd、Pb、Zn、As的积累量较高,说明纳塔栎对矿区Cd、Pb、Zn、As复合污染的适应能力较强,可作为以亚热带地区铅锌矿区Cd、Pb、Zn、As复合污染土壤的植被恢复及生态修复候选树种。
猜你喜欢
矿产综合利用(2020年1期)2020-07-24
中学生数理化·七年级数学人教版(2018年11期)2019-01-31
娃娃乐园·综合智能(2018年23期)2018-12-26
中成药(2018年8期)2018-08-29
娃娃乐园·综合智能(2018年3期)2018-03-22
中成药(2017年6期)2017-06-13
上海农业学报(2017年4期)2017-04-10
中国照明(2016年6期)2016-06-15
中国资源综合利用(2016年3期)2016-01-22
中国果业信息(2015年12期)2015-01-24
