
基于叶绿体DNA片段的槭属鸡爪槭组系统进化和生物地理学研究
2020-11-13 01:09:44李俊清
生态学报 2020年17期
高 健,于 涛,李俊清,*
1 内蒙古科技大学包头师范学院资源与环境学院,包头 014030 2 北京林业大学森林资源生态系统过程北京市重点实验室,北京 100083
槭属(AcerL.)是北半球地区仅次于栎属(QuercusL.)的第二大树木属,属内物种丰富(约124—200种),且广泛分布于亚洲、欧洲及北美洲等地[1- 2]。东亚,特别是中国长江流域及其以南地区,是槭属植物现代的多样性中心[3]。鸡爪槭组(sectionPalmataPax)是槭属中物种多样性最为丰富的分类组,组内物种为多年生乔木或灌木,落叶或常绿[1]。鸡爪槭组内物种形态特征变化丰富,具有掌状叶片从不裂(例如罗浮槭、滨海槭等)到3—13裂、叶边缘从全缘到锯齿、叶裂片从浅裂到深裂乃至全裂、花序类型多样(圆锥花序、伞房花序等)及果实长度变化较大(1—5 cm)等特征[2]。此外,鸡爪槭组物种集中分布于东亚地区(除了AcercircinatumPursh.分布于北美),是东亚槭属植物的代表类群[1- 2]。
鸡爪槭组物种具有重要的生态价值和科研价值。例如鸡爪槭(A.palmatumThunberg ex Murray)作为北温带广泛分布的园艺观赏树种,其种内人工选育出多种不同叶形及叶片颜色的鸡爪槭变种、变型等,在园林造景、植被恢复等生态建设中发挥着重要的作用[4]。东亚地区作为鸡爪槭组物种主要的分布区,其地貌类型复杂多样,且在末次冰期时未被大面积冰川所覆盖,其生物地理历史相较欧美地区来说更为复杂[5]。所以,鸡爪槭组物种是了解东亚地区植物进化历史的一个代表性材料。因此,本文对鸡爪槭组物种的起源、组内物种的分化时间、物种的扩散路径等进行了分析。该研究有助于了解鸡爪槭组的进化历史,并为东亚地区植物的生物地理历史提供了一个重要的范例。
1 材料与方法
1.1 试验材料
选取鸡爪槭组内18个物种(包含亚种),共27个个体作为研究材料(表1)。物种的鉴定与分类依据《Maples of the world》中的定义[2]。部分物种的序列于Genbank中下载得到(表1)。每个分类群选取1—3株个体,采集到的植物幼嫩叶片放入置有变色硅胶的取样袋中干燥、备用。同时,压制凭证标本,存放于内蒙古科技大学包头师范学院资源与环境学院。
1.2 DNA提取、PCR及测序
每份样品取约0.2 g干燥叶片,全基因组DNA的提取使用植物基因组DNA提取试剂盒(TIANGEN Beijing China)。依据本课题组前期对于鸡爪槭高分辨率DNA条形码的筛选结果,选取叶绿体基因片段psbA-trnH、rpl16和trnL-trnF作为本研究的遗传片段[4]。叶绿体基因片段的PCR扩增均在LabnetMultiGeneTM96-well Gradient Thermal Cycler上完成。PCR反应体系为20 μL,包含DNA模板1 μL、Taq PCR Master Mix(莱枫,上海)10 μL、正反引物各1 μL(表2)、二次水(ddH2O)7 μL。PCR反应程序为:94 ℃预变性3 min;94 ℃变性30 s,52—58 ℃退火30 s,72 ℃延伸60 s,共30个循环;最后72 ℃延伸10 min。PCR扩增产物经1%琼脂糖凝胶电泳检测并纯化后,送往北京生工生物工程股份有限公司在ABI3730XL遗传分析仪上进行双向测序。
1.3 系统发育分析
序列通过ClustalX[11]进行比对,并使用BioEdit 7.1.11[12]进行人工矫正。此外,对3个DNA片段的相容性进行ILD检测[13],1000次重复启发式搜索后显示片段间没有显著的冲突(P= 0.16)。故将3个片段合并后进行后续的联合分析。
金钱槭(DipteroniasinensisOliver)与云南金钱槭(DipteroniadyerianaHenry)在系统发育分析中作为外类群[14- 15]。最大似然法(ML)、最大简约法(MP)以及贝叶斯推理(BI)等分析分别使用PhyML 3.0[16]、PAUP*4.0b10[17]、MrBayes 3.2[18]进行计算。最佳的取代模型通过Modeltest 3.7[19]进行分析获取。
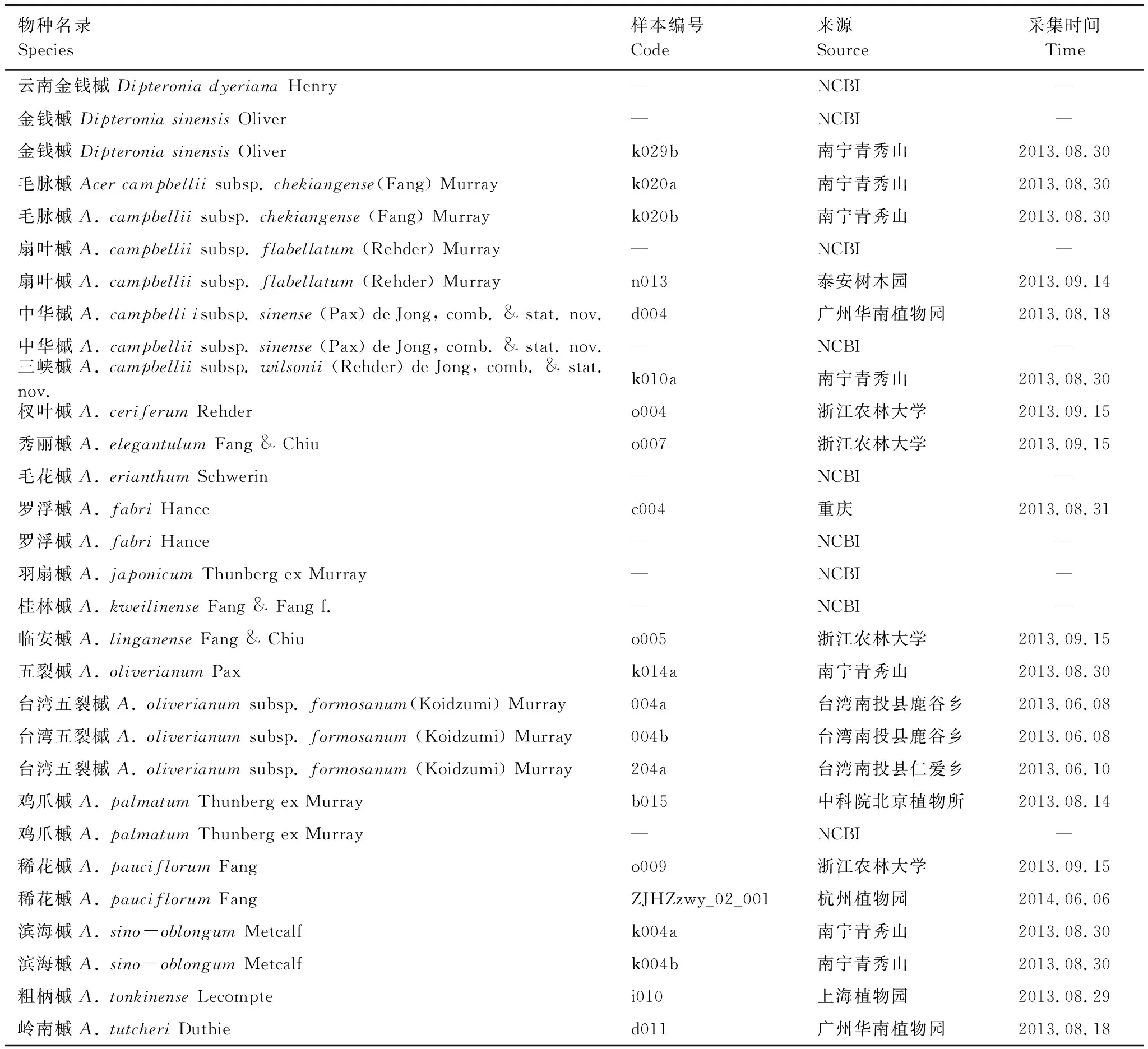
表1 实验物种基本信息

表2 基因片段PCR扩增的引物信息
1.4 分子定年及多样性速率分析
采用BEAST v.1.8.2[20]构建系统发育树,并推测鸡爪槭组物种间的分化时间。采用槭属与金钱槭属的分化时间(62 Mya)[21]和鸡爪槭组的分化时间(27.99 Mya)[22]作为化石校准点进行分子定年。进行1000000000次的MCMC模拟运算,每100000笔运算进行一次取样,总取样结果的前2500次取样不计入后续分析当中。
通过随机分枝过程(stochastic branching process,SBP)[23]绘制支系随时间累积趋势图(lineages through time,LTT),采用reversible-jump马尔可夫链蒙特卡尔(rjMCMC)模拟进行贝叶斯非参数估计(Bayesian non-parametric estimation)[24]来评估多样性速率。利用R语言的package APE[25]绘制LTT,并以APE中的mltt.plot功能,重新绘制以贝叶斯进化分析软件构建的1000次取样,以此来评估支系随时间累积的偏差。rjMCMC分析进行1000000次模拟,其中每1000次模拟提取1笔数据,对前10%的不稳定数据剔除后进行后续分析。γ统计被用来通过时间评估多样性速率的变异规律[26]。
1.5 祖先分布区重建
参考香茶菜属(Isodon)[27]、芭蕉科(Musaceae)[28]等祖先分布区重建研究中,依据地形对亚洲地理区块进行划分的方法,再结合本研究中鸡爪槭组物种主要分布于东亚、东南亚的特点,把鸡爪槭组物种的分布区范围主要划分为4个区块:(A)中国华东、华中、华南、华北、东北部分地区及朝鲜半岛;(B)中国西南及东南亚北沿地区(云贵高原、青藏高原等地);(C)日本地区;(D)中国台湾岛。采用R语言中的BioGeoBEARS软件包进行鸡爪槭组生物地理历史的推测[29]。
2 结果与分析
2.1 DNA扩增片段分析
序列比对后rpl16片段为909bp,含114个(12.5%)变异位点和74个(8.1%)有效信息位点;psbA-trnH片段为421bp,含72个(17.1%)变异位点和48个(11.4%)有效信息位点;trnL-trnF片段为902bp,含155个(17.1%)变异位点和109个(12.1%)有效信息位点。3个叶绿体DNA片段联合后,矩阵长度为2232bp,具有341个(15.2%)变异位点和231个(10.3%)有效信息位点。
2.2 系统发育关系
三种方法(ML,MP和BI)所构建的物种系统发育关系基本一致。18个鸡爪槭组物种(包含亚种)的27个个体形成单系群(99/95/1.0)(节点A)(图1)。稀花槭(A.pauciflorum)与滨海槭(A.sino-oblongum)的亲缘关系较近(99/57/0.64)。在节点B中,藏南槭(A.campbellii)种下的4个亚种之间没有归并为单系群。此外,羽扇槭(A.japonicum)与鸡爪槭(A.palmatum)被高度支持为单系群(96/94/1.0);秀丽槭(A.elegantulum)与罗浮槭(A.fabri)间的亲缘关系比较近(76/70/0.96)(图1)。
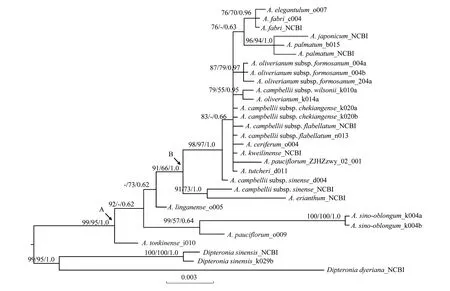
图1 基于rpl16,trnH-psbA以及trnL-F片段合并序列构建的系统发育树Fig.1 The tree combined chloroplast rpl16,trnH-psbA and trnL-F regionsML、MP分析中自展值高于50%及BI分析中后验概率高于50%被依次标于分枝上
2.3 分化时间及多样性速率估计
分子定年得出的物种树(图2)与基因树(图1)系统发育关系基本一致。结果显示:粗柄槭(A.tonkinense)较早从鸡爪槭组分化而出;临安槭(A.linganense)(浙江、安徽南)和飞蛾槭(A.sino-oblongum)(广东沿海)两个现代的局限分布种,分别约在19.92 Mya与15.58 Mya分化而来;12.42 Mya分化出鸡爪槭与羽扇槭的共同祖先,进而在4.18Mya分化出这两个物种;五裂槭(A.oliverianum)与其亚种台湾五裂槭(A.oliverianumsubsp.formosanum)的分化时间约为2.91 Mya;藏南槭在4.71 Mya开始其种内亚种的分化(图2b)。LTT结果显示,鸡爪槭组内物种的多样性速率保持相对稳定(图2a)。
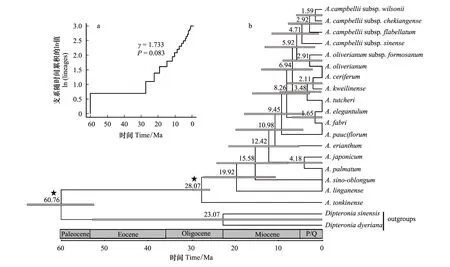
图2 鸡爪槭组物种的支系随时间累积趋势(a)及基于3个叶绿体DNA片段构建的鸡爪槭组植物分子定年结果图(b)Fig.2 The lineage-through-time (LTT) of Sect. Palmata (a) and Chronogram for Sect. Palmata reconstructed by three cpDNA sequences (b)
2.4 生物地理分析
通过BioGeoBears分析得出,进行鸡爪槭组生物地理分析的最佳模型是DEC模型(AIC=96.54,lnL=-45.92)。根据DEC模型的分析得出,A加B区域可能是鸡爪槭组物种的祖先分布区(图3)。此外,可能曾发生物种从亚洲大陆扩散至周边岛屿的情况,从而形成了日本特有种羽扇槭,我国台湾特有种台湾五裂槭以及目前分布于我国华南及台湾地区的岭南槭(A.tutcheri)(图3)。

图3 鸡爪槭组植物祖先分布区重建Fig.3 The reconstruction of ancestral area for Sect. Palmata
3 讨论与结论
3.1 组内种间的系统位置
鸡爪槭组内物种(包含亚种)被高度支持为单系群(99/95/1.0)(图1)[21,30]。在槭属分类书籍《Maples of the world》中记载A.campbellii中有4个亚种,理论上应当形成单系群,但结果显示这4个亚种分散在不同的分支中(图1)。而在相对较新的分类学著作《Flora of China》中,这4个亚种被处理为4个独立的物种,即:毛脉槭(A.campbelliisubsp.chekiangense)对应A.pubinerve;三峡槭(A.campbelliisubsp.wilsonii)对应A.wilsonii;扇叶槭(A.campbelliisubsp.flabellatum)对应A.flabellatum;中华槭(A.campbelliisubsp.sinense)对应A.sinense。此外,依据形态学证据,4个物种也存在较明显的差别。例如三峡槭叶片较其他3个物种(多为5裂,少7裂)来说多为3裂(少5裂),中华槭叶片较扇叶槭、毛脉槭(纸质或膜质)来说近革质,扇叶槭子房与花盘较毛脉槭(有毛)来说无毛等等。从地理分布上分析可以得出,4个物种主要集中分布于长江以南地区,但三峡槭、扇叶槭侧重分布于华南南部及西南地区(甚至延伸至越南、缅甸等地),而毛脉槭、中华槭侧重分布于华南及华东南部等地。所以,基于本研究结果以及形态特征、地理分布等证据支持《Flora of China》的分类结果,即把这4个亚种分别作为独立的物种进行分类处理。此外,研究结果支持鸡爪槭与羽扇槭间亲缘关系较近(图1)。
3.2 鸡爪槭组的起源
鸡爪槭组的起源时间约为28 Mya左右(渐新世中晚期)[22],且物种多样性在中新世时出现大幅度的增加(图2)。在渐新世中晚期至中新世,全球的气候变暖致使南极冰川的覆盖范围缩小[31],且在中新世中期(17—15 Mya)出现了气温的高点[32]。Donoghue和Smith对66个物种间断分布格局的形成进行了总结,得出绝大多数北温带森林物种在亚洲起源及物种多样化,且中新世是物种多样化的活跃期之一[33]。此外,在渐新世晚期至中新世初期,昆虫出现了植食现象(insect herbivory),随后昆虫的物种多样性在中新世得到了大大的提高[34]。总之,气候变化可能是影响鸡爪槭组物种起源及多样性增加的一个主要驱动力。
通过鸡爪槭组的生物地理分析得出,分布于我国广西、贵州南部、西藏东南部、云南东南部以及马来西亚、泰国、越南北部等地区的粗柄槭是组内较为早期分化出的物种(图3),也就是说鸡爪槭组中相对较为原始物种的分布区位于我国西南部及东南亚北部。郑维艳等指出我国西南部是亚热带常绿阔叶林许多科属的保存中心、现代分布中心和分化中心,且亚热带常绿阔叶林植物区系可能起源于我国的西南部[35]。徐廷志曾提出槭树科的起源地可能在我国横断山区附近[3,36],而鸡爪槭组在槭属中又被认为是较原始的类群[3,21]。加上本研究结果支持鸡爪槭组起源于A区域加B区域(图3),所以有理由推断鸡爪槭组的起源地位于我国西南部地区。这也从侧面支持了槭树科起源于横断山区[3,36]及槭属起源于亚洲的结论[22]。
3.3 鸡爪槭组的生物地理历史
鸡爪槭组植物在进化的历程中,存在多次的生物地理事件。例如,鸡爪槭与羽扇槭约在4.18 Mya出现分化(图2)。鸡爪槭的现代分布区域极广,广泛分布于中国、朝鲜半岛及日本地区,而羽扇槭作为鸡爪槭最近缘的物种却仅仅局限分布于日本。与之相类似的还有五裂槭(广布种)与台湾五裂槭(台湾特有种),两者的分化时间约在2.91 Mya(台湾岛形成时间约3—2 Mya)。两组物种的分化可能与第四纪冰期(开始于3—2 Mya)有直接的联系。第四纪冰期气候的变化导致了中国东海海平面的抬升与降低,在海平面下降时,亚洲大陆与日本、中国台湾等岛屿之间形成了东海陆桥,为分布于该地区的生物提供了迁徙的走廊[37,38]。而且,在4—3 Mya左右时,青藏高原的持续隆起[39],太平洋季风增强带来的丰沛降水[40]均对分布于东亚地区物种的扩散和多样化创造了客观条件。所以,对于鸡爪槭和五裂槭来讲,由于两物种较强的适应能力,冰期时可能借由东海陆桥分别扩散至日本岛、中国台湾岛,而冰后期两岛屿与亚洲大陆不再相连,地理隔离导致物种局地适应,而分别形成了现在的羽扇槭和台湾五裂槭(图3)。此外,依据中国植物志中的介绍,岭南槭种下包含分布于我国华南地区的岭南槭(原变种)与仅分布于中国台湾地区的小果岭南槭[1],所以岭南槭也可能曾借由东海陆桥进行物种的迁徙(图3)。
值得一提的是,纵观鸡爪槭组生物地理特征的结果,不难发现B区域物种是由A区域多次独立扩散而来的(图3)。而且,B区域物种同一进化分支中的姐妹种,多分布于A区域南部(亚热带地区),例如:稀花槭,秀丽槭,杈叶槭(A.ceriferum),中华槭等[1]。B区域多为高原地形,海拔较高,气温相对同纬度地区较低。所以,在周期性环境压力的背景下(例如:冰期与间冰期),鸡爪槭组物种为寻求合适生境,可存在多次往B区域扩散的现象(图3)。Qiu等通过对多种落叶阔叶林植物进行研究发现,亚热带地区阔叶林存在局部地区高低海拔迁移的现象[38]。由此而言,鸡爪槭组物种在进化历程中,可能存在方向性的扩散过程。
综上所述,可以推断得出以下结论:(1)我国西南地区可能是鸡爪槭组的起源地;(2)东海陆桥的隐没与抬升可能影响着羽扇槭、台湾五裂槭及岭南槭等鸡爪槭组物种的迁徙与扩散过程;(3)分布于亚热带亚洲大陆的鸡爪槭组物种,可能存在多次往同纬度高海拔地区扩散的现象。
致谢:感谢台湾师范大学生命科学系廖培钧教授、黄秉宏博士在数据处理上的帮助。
猜你喜欢
东坡赤壁诗词(2024年1期)2024-03-12 16:54:37
Natural Products and Bioprospecting(2023年6期)2023-12-29 13:42:30
红蜻蜓·低年级(2023年11期)2023-11-20 07:14:50
热带亚热带植物学报(2022年4期)2022-08-18 00:36:20
——和田盘羊
野生动物学报(2021年4期)2021-11-14 06:13:24
小读者(2021年4期)2021-06-11 05:42:26
中学生天地(C版)(2020年4期)2020-05-08 03:39:48
小学生导刊(2018年20期)2018-11-30 03:05:12
散文诗(2017年18期)2018-01-31 02:43:41
中学生阅读(初中版)(2016年11期)2017-01-13 01:06:40
