
整合性学习观的动物行为模型探索*
2020-11-13 05:31武晓睿
心理学报 2020年11期
尹 彬 武晓睿 连 榕
整合性学习观的动物行为模型探索*
尹 彬 武晓睿 连 榕
(福建师范大学心理学院, 福州 350108)
我国当下的学习观以渐进性学习为主。研究尝试提出“整合性学习”的学习观, 以40只SD大鼠为被试, 采用2(学习方式:整合/渐进) × 2(性别:雄/雌)组间设计, 运用14单元组合T迷宫进行五阶段的动物行为建模。结果显示, 1)学习错误次数上, 整合少于渐进、雄性少于雌性; 2)整合较渐进学习效果的迁移性更好; 3)各组皆出现对第一段原正确路径的固着。由此得出:整合性学习更长效, 习得的知识具有整体性、组块化、范畴化的可迁移的 特征。
整合性学习观, 动物行为模型, 迷宫测试
1 前言
学习观, 是指学生个体对知识、学习现象和经验的直观认识(刘儒德, 2002)。人类已经进入了信息社会、智能时代(后工业时代、后现代社会), 传统的学习观已经不能很好地解释新时代学习现象的变化, 急需我们立足新的发展、以新的研究去发现学生学习心理的新成分(施良方, 2000)。自伴随工业革命出现的学校教育, 学生的学习一直强调的是循序渐进、从部分到整体的学习方式, 在知识相对贫乏、获取途径单一的时代, 这种学习方式符合其情境的需要。随着知识的极大丰富、获取途径的多样化的时代的到来, 这种学习方式的固有问题越来越显现。其中一个突出现象就是学生的学习负担、学习压力不断在加大、加重。虽然各种教学改革都以此为改善目标, 但学生的学习负担却是只加不减, 问题的根源就在于我们对已质变的学习现象缺乏新的认识。
基于此, 我们尝试提出整合性学习观:“整合性学习”是指在元认知的作用下认知积极统整学习材料, 实现高效且深入地对知识的理解和掌握的过程,是元认知与认知高度合一的学习心理过程; 与此对应的非整合性学习(如渐进式的), 是强调在认知层面上积极渐进地从部分到整体的对知识的理解掌握过程。两种学习观的主要区别在于, “整合性学习”是强调元认知与认知高度合一的学习心理机制, 非整合性学习强调的是认知的学习心理机制。
那么这两种学习观的差异是否存在生物心理学基础呢?动物行为的教学和研究有助于我们理解行为背后的基本原则, 并且为人类提供了方便和高度控制的模型(朱滢, 2019)。因此, 想要了解整合性学习观对于学习效果的影响, 并进一步找到对应的神经生理证据去证实, 需要像当年提出联结主义学习理论的桑代克(张厚粲, 2003)、提出操作性条件反射和强化理论的斯金纳(Skinner, 1945; 另见周正怀, 2005a, 2005b)以及提出“认知地图”理论并首次在心理学研究中引入中介变量的托尔曼(Tolman, 1948)一样, 首先应进行的是动物行为建模研究。通过研究大鼠对迷宫路径信息的整合过程(王彦, 苏彦捷, 2001), 我们可以从中获得基础的理论启示, 为阐明整合性学习观的特征、效果和内在机制提供进化发展上的证据。
我们利用和改造了Tolman (1948)的14个单元的组合T迷宫, 将整合性学习方式操作性定义为“大鼠在对迷宫路径整体把握的基础上, 积极整合路径信息与内部认知, 高效地习得可迁移的路径信息的学习方式”; 渐进性学习方式的操作性定义为“根据迷宫路径的可迁移规律, 迷宫在路段和时程上进行硬性规划, 逐步增加大鼠学习材料容量和可迁移信息的学习方式”。这样定义的原因在于, 整合性学习的本质特征是学生掌握知识的过程实现元认知与认知的高度合一。Schraw (1998)提出元认知是对于认知过程的认知, 具有认知和监控两个层面。因此, 在整合性学习观指导下的整合性学习方式中, 学习者是在元认知监控下进行“整体−部分−整体”的学习; 而渐进性学习方式由于缺乏考虑“元认知”这个认知成分, 学习者只进行“部分−整体”的学习。在动物行为建模中, 我们将整合性迷宫路径设计为“整段路径开放”。大鼠可以在一开始就有机会感知“整体”路径, 并在学习的过程中不断的进行“整体−部分”的关联, 最终掌握“整体”迷宫路径; 而我们将渐进性迷宫路径设计为“三段路径依据规律进行逐步开放”, 大鼠依据“部分”路径随时间逐步开放的方式进行“部分”学习, 最终掌握“整体”的迷宫路径。
另一方面, “学”是“教”的基础。Marshall (1992)认为“以学定教”是教育变革与研究的主要途径, 从学生视角探究学生如何学习已经成为教育变革的起点。我们不仅需要关注“教学者”设计合理的“教学方式”这一外部过程, 也需要关注“学习者”如何学习这一内部过程。而动物迷宫的相关研究已经发现雄性在空间导航的策略和效果上不同于雌性(Hawley et al., 2012; Keeley et al., 2013; Perrot-Sinal et al., 1996; Roof, 1993; Saucier et al., 2008), 以人类为被试的研究也发现了类似的现象(Munion et al., 2019; Sneider et al., 2015), 这些研究暗示着性别变量影响着学习者在空间学习任务中的内部学习过程。由于学习者(大鼠)在整合性学习条件下自发的学习过程是本研究的重点, 因此为了探究不同性别的大鼠是否都具有自发的“整合”行为, 以了解整合性学习方式的最适受众特征和使用其的边界(支持)条件, 我们同时设置了“性别”因素进行探究。
最终, 我们以14个单元的组合T迷宫为实验装置, 采用2(学习方式:整合; 渐进) × 2(性别:雄; 雌)的两因素组间设计, 比较4组大鼠迷宫学习的效果, 并设计了一系列后续实验任务, 以探究学习效果差异的行为学层面上的内在机制。
2 方法
2.1 被试
被试为40只一月龄SD大鼠(20只雄性, 20只雌性), 分为整合雄(Integrative Learning-Male, IL- Male)、整合雌(Integrative Learning-Female, IL- Female)、渐进雄(Progressive Learning-Male, PL- Male)、渐进雌(Progressive Learning-Female, PL- Female) 4组, 每组10只。所有被试被随机分配到8个饲养笼中, 每笼5只, 饲养在恒温22.5℃、光照自动化控制(8:00关灯, 20:00开灯)、空气净化器24小时运作的清洁级动物房中。
正式实验前1周, 每天对大鼠进行15分钟的触摸互动。正式实验时, 在单笼实验结束后依次喂食, 每笼25 g, 控制大鼠为半饥饿状态; 实验间歇期, 喂食时间固定, 每笼50 g, 控制大鼠为饱食状态; 实验期间, 大鼠体重保持在自由喂食体重的85%以上。学习阶段, 雄性(89.88 ± 2.04 g)和雌性(89.27 ± 2.04 g)的体重差异不显著,(1, 36) = 0.042,= 0.840; 整合组(87.16 ± 2.04 g)和渐进组(91.99 ± 2.04 g)的体重差异不显著,(1, 36) = 2.83,= 0.100。在第四个测验任务中, PL-Male-6被试由于体重过轻死亡, 因此其后3次的概括分析任务和4次的阶段固化测验数据空缺。
实验全过程经福建师范大学实验动物伦理审查委员会认证通过(IACUC-20180019)。
2.2 工具和材料
14单元组合T迷宫(图1a)、实验动物日常养护用具、清洁防护用具、视频监控用具、视频处理与数据分析软件等, 详见附录支持材料(Supporting Materials)的S1.2。
2.3 实验设计
研究采用2(学习方式: 整合/渐进; IL/PL) × 2(性别: 雌/雄; Female/Male)两因素组间设计, 以学习次数、错误次数(错误探测次数和错误进入次数之和)、行进路程、完成时间为量化指标, 以动物特征行为及行径图为质化指标。为探究不同学习方式的效果及内在机制, 设置学习任务、一周后复测任务、格式塔(逆向)迁移任务、概括分析任务和阶段固着任务5个试验任务(图1c-f)。详细的任务设计说明见网络版附录S1.3。
2.4 实验程序
(1)适应阶段, 对大鼠进行为期一周的触摸互动, 每次每笼15分钟。投放足量食物(50克每笼), 记录其体重、饮水量和剩余食物量。
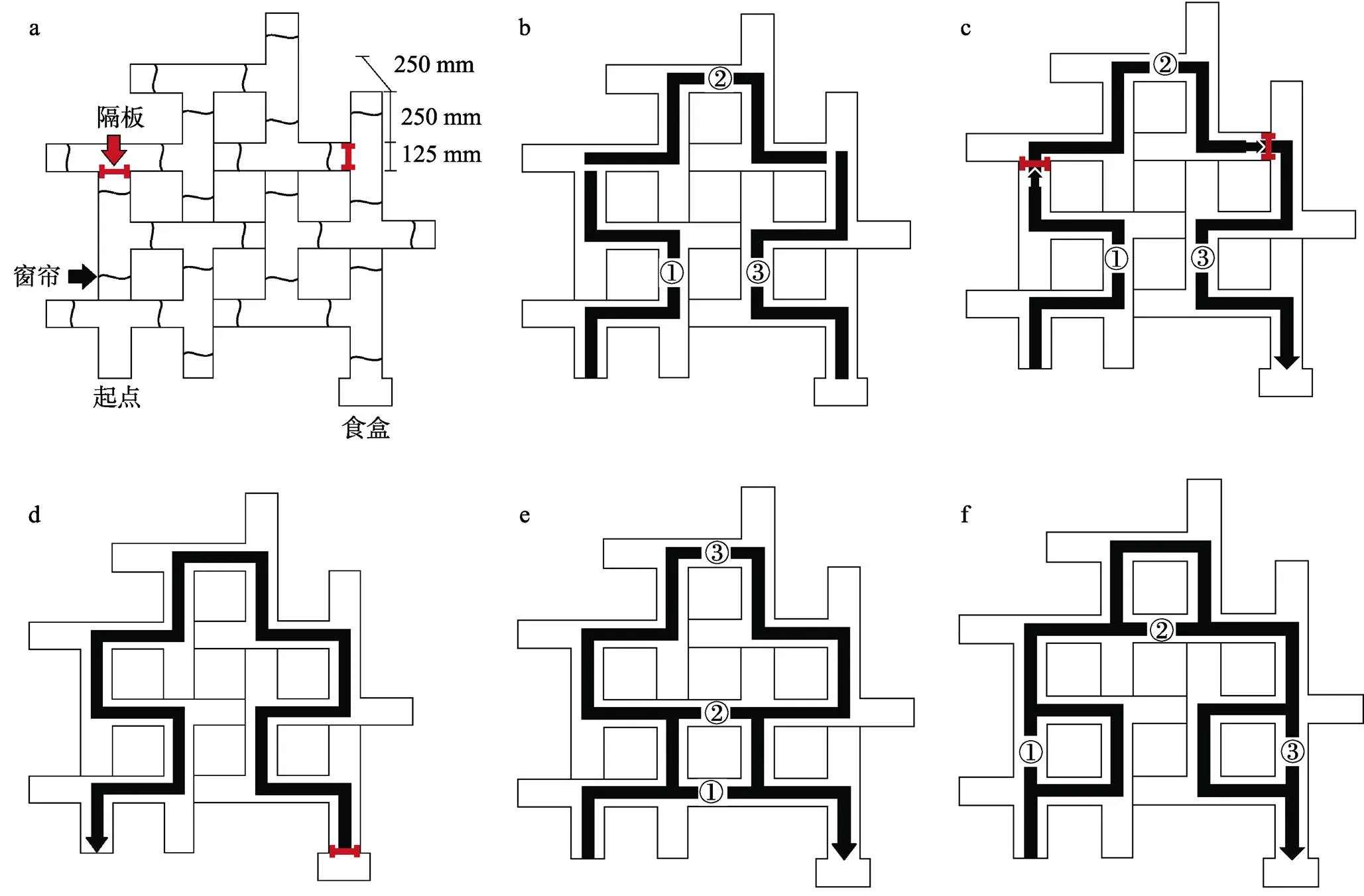
图1 测验任务设计图
注:a)基本实验装置, 改造自Tolman (1948)。b)将基础实验装置的正确路径划分为具有形状相似性的三段, ①、②、③路径依次为第一路径、第二路径和第三路径, 以方便结果报告时描述大鼠行为特征。c)学习和复测任务设计图。IL组不设置分段隔板全程开放, PL组依据迷宫的长度和规律用隔板分为3段(隔板放置处有凹槽, 可抽插)。① (第一路径)、② (第一、二路径)、③ (全路径)路线分别是PL组1~3天、4~7天、8~12天的正确行径路线, ③ (全路径)路线也是IL组1~12天的正确行径路线。复测时, 保持全路径开放。d)格式塔(逆向)迁移任务设计图。食盒入口被隔板密封, 变更为起点, 原起点的窗帘移至终点岔路口处。e)概括分析任务设计图。图中①、②、③分别为最短路线、隐蔽路线和原路线。f)阶段固着任务设计图。①、②、③分别为第一路径、第二路径和第三路径的最短替代路径, 原路径仍保持通畅。

图2 研究流程图
(2)正式实验包括5个任务(见图2)。首先是为期12天的学习任务(每天一个试次), 第1天各组皆限时15分钟, 2~12天在大鼠进食结束后即取出(15分钟内)。一周后, 进行3天的一周后复测任务; 接着是3天的格式塔迁移任务; 再按学习阶段路径重新学习巩固一天, 然后进行5天的概括分析任务; 再按学习阶段路径重新学习巩固一天, 然后进行4天的阶段固着任务。所有任务皆为大鼠15分钟内进食结束后取出。
(3)实验期间填写观察记录表, 使用SuperMaze v4.0动物行为视频分析系统(上海欣软)对实验录像视频进行动物运动轨迹分析和数据导出(包括轨迹图和热图), 并使用Microsoft Excel、IBM SPSS 18.0、GraphPad Prism 8、3D画图等软件进行数据分析和图表制作。实验细节详见网络版附录S1.1及S1.4。
2.5 统计方法
在统计分析中, 以学习方式(IL/PL)和性别(Male/Female)为组间变量, 学习次数和路径(第一路径/第二路径/第三路径)为组内变量, 错误次数、学习成功天数、是否出现某种特定行为以及出现某种特定行为的比例为因变量, 进行了不同变量组合下的方差分析, 原始数据以及图表对应分析数据详见网络版附录S2。
3 结果
3.1 学习记忆效果
学习任务中, 如图3a所示, IL组学习曲线呈“指数下降型”, PL组则呈“波浪型”; 以学习方式和性别为组间变量, 学习次数为组内变量进行三因素重复测量方差分析, 结果显示, 学习方式、性别和学习次数的主效应均显著, 且两两之间存在交互作用(详见网络版附表1, 位于附录支持材料中的S2实验结果统计分析表格, 下同)。在学习成功天数上(图3b), 以学习方式和性别为组间变量进行两因素完全随机方差分析, 结果显示(网络版附表2), 主效应(性别:(1, 36) = 0.72,= 0.551, η20.42, 95% CI = [−2.63, 0.33]; 学习方式:(1, 36) = 3.29,= 0.321, η20.77, 95% CI = [−3.93, −0.98])和交互作用((1, 36) = 3.45,= 0.072, η20.09, IL和Male: 95% CI = [3.23, 6.18]; PL和Male: 95% CI = [7.03, 9.98]; IL和Female: 95% CI = [5.73, 8.68]; PL和Female: 95% CI = [6.83, 9.78])虽不显著, 但效应量较大; 关于学习方式的事后多重比较显示(网络版附表3), IL短于PL ((1, 36) = 11.36,= 0.002, η20.24, 95% CI = [−3.93, −0.98]); 控制性别因素, 对学习方式进行简单效应比较显示(网络版附表4), IL-Male组和PL-Male组的差异显著((1, 18) = 4.12,= 0.000, Cohen’s= −1.84,= −0.67, 95% CI = [−6.21, −1.40])。
一周后复测任务中(图3c), 三因素重复测量方差分析显示(网络版附表5), 不同组别错误次数无显著差异,(1, 36) = 4.03,= 0.052。
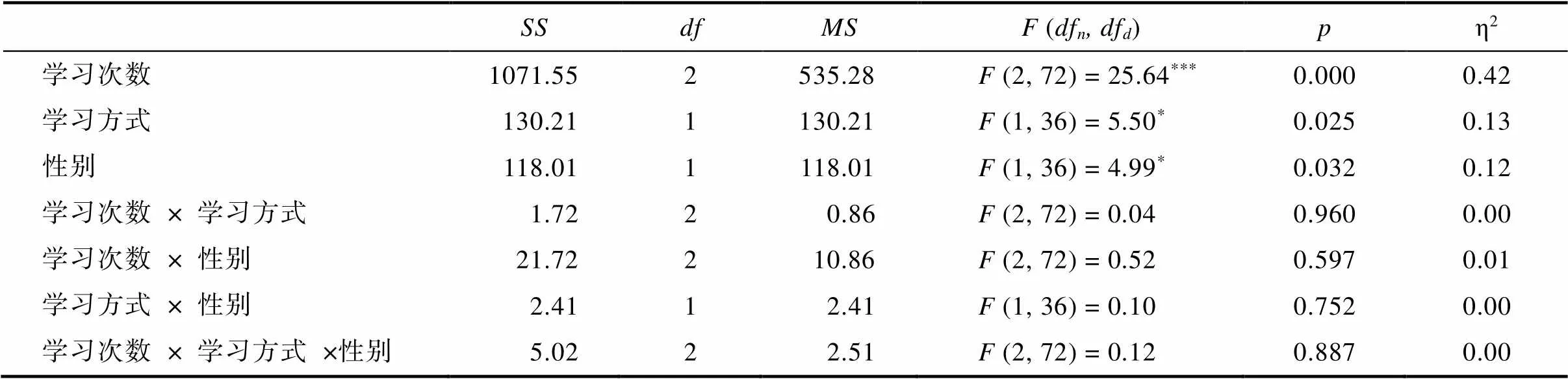
格式塔迁移任务中(图3d), 三因素重复测量方差分析显示(网络版附表6), IL组错误次数显著少于PL组,(1, 36) = 5.50,= 0.025, η20.13, 95% CI = [0.28, 3.89]; 雄性错误次数少于雌性,(1, 36) = 4.98,= 0.032, η20.12, 95% CI = [−3.79, −0.18]。
在概括分析任务中(图4b), 以学习方式、性别和学习次数为三因素的重复测量方差分析显示(附表7), IL组5天的错误次数少于PL组,(1, 35) = 4.66,= 0.038, η20.12, 95% CI = [−0.99, −0.03]。通过观察首次测验的分组热图(图4a)可知, IL组几乎全部直接选择原路线或最短路线, 而PL组出现了路线选择上的混杂(三条路线都选), 表现出全盘重新搜索的特征。

图3 学习记忆效果对比图
注:a)学习曲线:学习阶段中的错误次数结果汇总图。其中, “错误次数” = “错误进入次数”+“错误探测次数”, 从起点开始计算, 到达终点食盒为止。错误进入次数指大鼠身体的头部和重心进入错误区域(一般需穿过窗帘), 而错误探测次数指大鼠头部进入错误区域(一般没有穿过窗帘)。b)学习阶段中的学习成功天数图。学习成功天数定义为从第一次学习开始到进入“开放路段”出现零错误次数之前的天数总和。其中, IL组的“开放路段”是迷宫全路径, 因此出现零错误次数之前的天数总和记为学习成功天数; PL组学习过程中的“开放路段”是分三个阶段逐步开放的, 因此每个阶段出现零错误次数之前的天数总和记为学习阶段成功的天数。c)一周后复测任务的错误次数结果图。d)格式塔迁移任务的错误次数结果图。各统计图中的圆圈、三角符号的位置或柱的高度显示的是每个试次中各组的平均值, 误差线显示的是标准误。**< 0.01。
在阶段固着任务的最后一次测试中(图4c-d), 在第一路径中各组仍一致的选择原路径, 第二路径中选原路径和新路径的二者兼有, 第三路径中各组一致的选择新的最短路径。在各路径中选择原路径编码为1, 选择新路径编码为0, 以学习方式、性别和路径为三因素进行重复测量方差分析显示(网络版附表8), 学习方式和性别主效应不显著, 路径主效应显著,(2, 70) = 40.83,< 0.001, η20.54, 95% CI = [−0.07, 0.24], 表现出各组大鼠均一致的对第一路径的原路线的固着、对第二路线的新路线的尝试和对第三路径的新路线的偏好。
3.2 IL组学习阶段行为特征
为进一步探究IL组在学习过程中的内在特征, 我们对IL组在学习过程中的行进路线进行了追踪分析(图5)。
如图5a所示, IL组学习阶段的轨迹图分布大致有3类, 其中a左图(先反复学习左半段而后迅速完成右半段)和a中图(先反复学习第一路径和第二路径后迅速完成第三路径)出现在IL-Male组学习的第1天和IL-Female组学习的前3天; a右图(仅仅在第一路径上有自发的折返学习而后迅速完成第二路径和第三路径)普遍出现在IL-Male组第2天至学习成功天数之间, 且IL-Female组的9只在12天中至少出现1次, 3只IL-Female组在前三天出现15分钟内未到达终点的情况。
以性别、分段路径为两因素进行重复测量方差分析(图5b、c, 网络版附表9、10), 结果显示, 路程指标上, 分段路径的主效应显著((2, 36) = 38.01,< 0.001, η20.68, 第一路径和第二路径:95% CI = [5.52, 15.63], 第一路径和第三路径:95% CI = [10.81, 23.45], 第二路径和第三路径:95% CI = [2.48, 10.63]); 时间指标上, 分段路径的主效应也显著((2, 36) = 39.10,< 0.001, η20.69, 第一路径和第二路径:95% CI = [45.57, 111.60], 第一路径和第三路径:95% CI = [62.08, 138.43], 第二路径和第三路径:95% CI = [1.02, 42.33]), 呈逐渐递减趋势; 性别的主效应显著, 雄性比雌性的行进路程((1, 18) = 22.51,< 0.001, η20.56, 95% CI = [−21.10, −8.15])和行进时间((1, 18) = 22.10,< 0.001, η20.55, 95% CI = [−96.70, −36.97])皆更短; 性别和分段路径存在交互作用(行进路程:(2, 36) = 6.26,= 0.005, η20.26; 行进时间:(2, 36) = 9.10,= 0.001, η20.34)。

图4 概括分析任务和阶段固着任务分段路径对比图
注:a)概括分析任务首次测试分组热图对比。b)概括分析任务5天错误次数对比图。c)阶段固着任务末次测试分组热图对比。d)阶段固着任务分段路径固着比例对比图。热图中亮度越高的地方表示探索密度较高。统计图中的圆圈符号的位置或柱的高度显示的是每个试次中各组的平均值或占比的多少, 误差线显示的是标准误。*< 0.05。
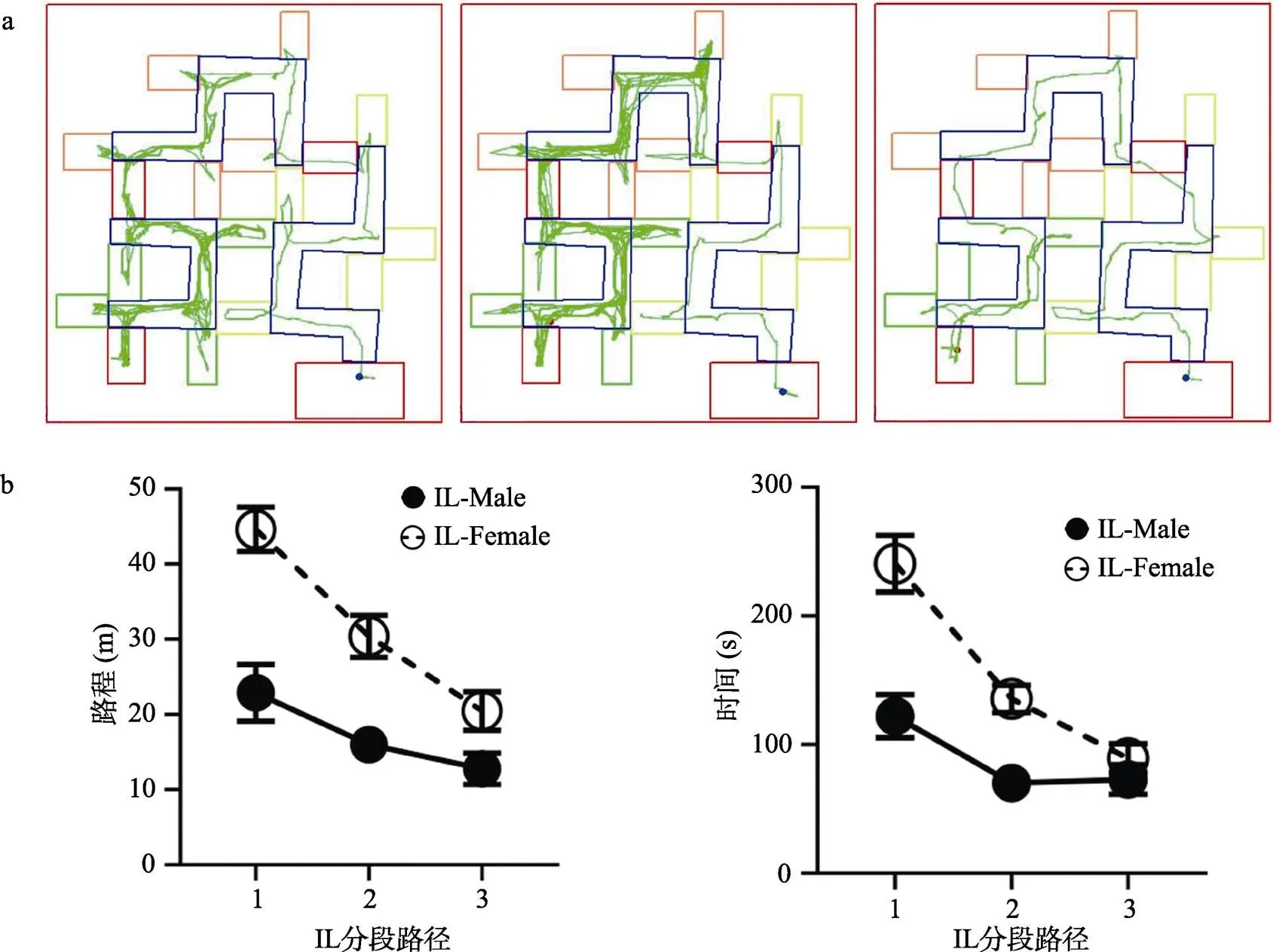
图5 IL组学习过程分段路径分析结果图
注:a)IL组典型轨迹图个案, 从左到右分别为IL-Female-2-1, IL-Female-6-1, IL-Female-9-1。b)和c)IL组的行进路程和时间分段对比图。分段时间的计算方式为“分段路径的行进时间 = 持续行进时间 − 起点静止时间”。其中, ①减除“起点静止时间”是为了排除大鼠在第一段路径产生起点固着而造成第一段时间显著较长的系统误差; ②在路程或时间指标上, 每只IL组大鼠的“第一路径”、“第二路径”、“第三路径”三个数据单元皆为大鼠“学习成功”前的n天的总路程或总时间; ③“学习成功”是指大鼠从开始学习至无错误次数的走完全程为止, 是因个体而异的; 只在“学习成功”的天数范围内分析分段路程和分段时间是为了使数据分析聚焦大鼠对迷宫正确路径“从不会到会”的学习过程, 而不受会了以后的行为表现的影响。统计图中的圆圈符号的位置显示的是每个试次中各组的平均值, 误差线显示的是标准误。
总体来看, IL组的行进轨迹呈现出大鼠对第一段正确路径更密集的学习, 即区域分布不均匀的现象, 其中雄性比雌性更典型。
3.3 PL组学习阶段行为特征
同时, 为了探究PL组在学习阶段的行为特征, 我们着重探究了PL组在新学习路径开放时第一天的表现(第二路径和第三路径的开放分别为总学习过程的第4天和第8天)。如图6及网络版附图7、8所示, PL-Male组在第二路径开放后仍保持对第一路径的高密度探索, 而PL-Female组则对新开放的第二路径进行了高密度探索。以性别、分段路径和学习次数为三因素进行重复测量方差分析, 结果显示(网络版附表11、12), 在行进路程上, 分段路径和性别存在交互作用,(1, 18) = 5.35,= 0.033, η20.23; 且分段路径、学习次数和性别间存在交互作用,(3, 54) = 3.13,= 0.033, η20.15。学习第三阶段(8~12天)的行径路程和时间上未出现性别上的显著差异(网络版附表13、14及附图9、10)。

图6 PL组学习过程中开放新路径后的探索密度图
注:学习阶段PL组第4天和第8天的分组热图, 颜色亮度越高表示探索该区域的密度越高。
总之, 在新路段开放后, 不同性别的PL组大鼠的探索密集路段不同, 其中, PL-Male组集中探索原路径, PL-Female组集中探索新开放路径, 推测PL-Male组注重新旧路段的联结, 而PL-Female将新开放路段当作全新的路段进行学习。
3.4 学习阶段的个体适应性
为了探究整合性学习方式是否适用于每个个体, 我们对于学习阶段中, 被试在起点的静止时间、到达终点时是否在原地吃食、是否吃巧克力(作为一种更有诱惑力但被试之前未接触过的新异奖赏刺激)和体重等指标进行观测, 作为学习方式适应性的指标。以学习方式、性别和学习次数为三因素的重复测量方差分析显示(图7, 网络版附表15, 附图11、12), IL组比PL组更少在原地进食(0.51 ± 0.05 vs 0.78 ± 0.05,(1, 34) = 15.93,< 0.001, η20.32, 95% CI = [−0.42, −0.14]), 更少的进食巧克力(0.83 ± 0.03 vs 0.92 ± 0.03,(1, 36) = 4.44,= 0.031, η20.12, 95% CI = [−0.16, −0.01]), 但学习方式和性别之间的交互作用不显著(原地进食:(1, 34) = 2.92,0.097; 吃巧克力:(1, 36) = 4.11,0.050); 体重、起点静止时间等因素主效应不显著且无交互作用。

图7 特定行为观察指标
注:比例是指被试在学习阶段到达终点在原地吃或吃掉巧克力的次数占总统计次数的比例。图中的柱高的位置显示的是的平均值, 误差线显示的是标准误。*< 0.05, ***< 0.001。
4 讨论
4.1 不同学习方式的的学习效果差异
总体来看, IL组的学习效果好于PL组, 体现在以下几个方面:1) IL组学习成功天数更短; 2)在一周后复测任务中, 不同学习方式的大鼠在正确路段记忆层面上无显著差异, 但在考察大鼠对迷宫整体把握程度的格式塔(逆向)迁移任务中, 又出现了分离, 表明IL组学习阶段形成的对迷宫路径的整体认知更牢固且可迁移; 3)在概括分析任务中, IL组错误次数更少, 第一天测试的热图显示, 其更明确的选择走最短路线和原路线, 而PL组则出现了全盘重新探索的行为, 表明相对于PL组, IL组在记忆迷宫路径的基础上, 对起点和终点的绝对方位有更明确的把握, 可能存在对记忆路线的多层面的范畴化加工。
在学习过程中, IL组的轨迹图(图5)以及相应指标显示, IL组对第一、第二和第三路径存在探索密度的递减趋势; 结合阶段固着测验中各组对第一路径(原路径)的持续性固着的结果, 我们认为IL组在整体迷宫范围内, 对第一路径进行了重点记忆加工。研究发现, 海马可以编码空间记忆, 同时也可以进行“元空间”的信息加工并且这一类信息是可迁移的(Dusek & Eichenbaum, 1997); 通过调控海马细胞的突触可塑性可以增强“元空间”的信息加工以及空间方位预判的能力(Bannerman et al., 1995; Saucier & Cain, 1995)。由于第一路径和第二、三路径的正确路径形状一致, 因此我们推测IL组在记忆第二、三路径时迁移了第一路径的记忆信息。此外, 阶段固着任务中, 第二、三路径各组均出现选择最短路径的行为, 我们推测大鼠使用迁移方式记忆的信息固化程度可能更小, 在面对新的任务时, 该记忆信息更易被优化。而IL组自发的选择第一路径进行重点探索, 然后较为快速的完成对第二路径和第三路径的学习, 暗示其在不同路径间建立了关系和联结, 并进行了组块化的划分, 而这种行为具有自发性, 可能是一种“自适应”的学习现象。
但IL组的部分个体在学习过程中, 尤其是在学习阶段初期, 会出现显著的焦虑和适应不良(图6), 表现为更少在终点原地进食, 以及更少的去吃作为新颖刺激出现的巧克力(一种Novelty-suppressed feeding现象, 可参见(Bechtholt et al., 2007; Dulawa & Hen, 2005; Dulawa, 2009), 可能需要更多的支持。
4.2 性别差异
在特定的迷宫学习任务中, 雄性的学习行为明显异于雌性, 这与以往研究中雄性和雌性在空间方位任务上的行为表现的差别相一致(e.g. Hawley et al., 2012; Keeley et al., 2013; Perrot-Sinal et al., 1996; Roof, 1993; Saucier et al., 2008)。从学习阶段的轨迹图和热图分析来看, IL-Male组对三段路径的组块划分更明确, 表现为轨迹图中, 着重探索第一路径的行为更频繁和普遍; IL-Female组则显得更为谨慎, 出现与IL-Male组相似的规律性探索行为较晚, 且到了终点以后更加不敢在原地进食或吃掉巧克力。PL-Male组有同样的行为倾向, 表现为热图中, 第二路径开放后更多的折回去探索第一路径, 第三路径开放后对三个路径的探索较为平均; 而PL-Female组在每次新路径开放后, 都更高密度的探索新路径。因此, 我们推测PL-Male组同样倾向于基于对第一路径的深度加工, 建立新旧路径之间的记忆联结, 将对原路径的认知迁移到新路径上, 只是其潜力被人为的阶段划分限制住了; 而PL-Female组则更倾向于直接探索未知的新路径。
4.3 对学习的启示
尽管“整合性学习”这一名词概念已有学者提出过(陈琦,张建伟, 2003; 方华梁, 2018), 但本研究是首次基于动物行为建模对这一概念的内涵和特性进行了诠释, 提示了整合性学习观在自然规律上的可能性。Yeager和Walton (2011)曾强调以心理学手段对学生的学习行为进行干预时, 需要注意情境的作用。本研究暗示着教养者对于情境的理解可以更加丰富和深层, 例如一种基于学生学习“整体−部分−整体”的“自适应”过程的整合性学习观可能对于长期的知识掌握和迁移利用而言是更加有效的, 只要教养者能够根据学习者的个体属性(如性别、主动性人格等, 参见Zhu et al., 2017)以及学习任务的性质、学习环境的支持度等情境因素(Walton & Yeager, 2020), 在初期提供足够相应的支持以帮助他们度过适应期。
5 结论
(1)对于大鼠迷宫学习而言, 整合性学习方式更高效, 习得的信息更多层, 具有长期性和辅助迁移的优势;
(2)在大鼠形成对迷宫路径整体认知的过程中, 记忆信息具有整体性、组块化、范畴化和可迁移的特征;
(3)在迷宫学习任务中, 雄性大鼠和雌性大鼠的学习行为存在明显差异;
(4)整合性学习过程中, 部分大鼠会在初期出现更多的焦虑或适应不良。
Bannerman, D. M., Good, M. A., Butcher, S. P., Ramsay, M., & Morris, R. G. M. (1995). Distinct components of spatial learning revealed by prior training and NMDA receptor blockade.(6553), 182−186.
Bechtholt, A. J., Hill, T. E., & Lucki, I. (2007). Anxiolytic effect of serotonin depletion in the novelty-induced hypophagia test.,(4), 531−540.
Chen, Q., & Zhang, J. W. (2003). A model of integrated learningin the information era., (3), 90−96.
[陈琦, 张建伟. (2003). 信息时代的整合性学习模型——信息技术整合于教学的生态观诠释., (3), 90-96.]
Dulawa, S. C., & Hen, R. (2005). Recent advances in animal models of chronic antidepressant effects: The novelty-induced hypophagia test.,(4-5), 771−783.
Dulawa, S. C. (2009). Novelty-induced hypophagia.,, 247−259.
Dusek, J. A., & Eichenbaum, H. (1997). The hippocampus and memory for orderly stimulus relations.(13), 7109−7114.
Fang, H. L. (2018). Integrative learning in general education curriculum: The case of a great book reading course.,(2), 61+67+62−66.
[方华梁. (2018). 通识教育课程的整合性学习——对一门名著阅读课程的个案研究.,(2), 61+67+62−66.]
Hawley, W. R., Grissom, E. M., Barratt, H. E., Conrad, T. S., & Dohanich, G. P. (2012). The effects of biological sex and gonadal hormones on learning strategy in adult rats.,(4), 1014−1020.
Keeley, R. J., Tyndall, A. V., Scott, G. A., & Saucier, D. M. (2013). Sex difference in cue strategy in a modified version of the Morris water task: Correlations between brain and behaviour.,, e69727.
Liu, R. (2002). On conception of learning of college students., (4), 77−81.
[刘儒德. (2002). 大学生的学习观., (4), 77−81.]
Marshall, H. H. (1992)..Ablex Pub. Corp.
Munion, A. K., Stefanucci, J. K., Rovira, E., Squire, P., & Hendricks, M. (2019). Gender differences in spatial navigation:Characterizing wayfinding behaviors.,(6), 1933−1940.
Perrot-Sinal, T. S., Kostenuik, M. A., Ossenkopp, K. P., & Kavaliers, M. (1996). Sex differences in performance in the Morris water maze and the effects of initial nonstationary hidden platform training.,(6), 1309−1320.
Roof, R. L. (1993). Neonatal exogenous testosterone modifies sex difference in radial arm and Morris water maze performance in prepubescent and adult rats.,(1-2), 1−10.
Saucier, D., & Cain, D. P. (1995). Spatial learning without NMDA receptor-dependent long-term potentiation.(6553), 186−189.
Saucier, D. M., Shultz, S. R., Keller, A. J., Cook, C. M., & Binsted, G. (2008). Sex differences in object location memory and spatial navigation in Long-Evans rats.,(1), 129−137.
Schraw, G. (1998). Promoting general metacognitive awareness.,(1/2), 113−125.
[Shi, L. F. (2000).Beijing, China: People’s Educational Press.
[施良方. (2000).. 北京: 人民教育出版社.]
Skinner, B. F. (1945). The operational analysis of psychological terms.(4), 547−553.
Sneider, J. T., Hamilton, D. A., Cohen-Gilbert, J. E., Crowley, D. J., Rosso, I. M., & Silveri, M. M. (2015). Sex differences in spatial navigation and perception in human adolescents and emerging adults.,, 42−50.
Tolman, E. C. (1948). Cognitive maps in rats and men.(4), 189−208.
Walton, G. M., & Yeager, D. S. (2020) Seed and soil: Psychological affordances in contexts help to explain where wise interventions succeed or fail.,(3), 219−226.
Wang, Y., & Su, Y. J. (2001). Mazes and animal behavioral research.(3), 264−269.
[王彦, 苏彦捷. (2001). 迷宫与动物行为研究.(3), 264−269.]
Yeager, D. S., & Walton, G. M. (2011). Social-psychological interventions in education: they’re not magic.(2), 267−301.
Zhang, H. (2003).Hangzhou, China: Zhejiang Educational Press.[张厚粲. (2003).. 杭州: 浙江教育出版社.]
Zhou, Z. H. (2005a). On the common characters between E. L. Thorndike and B. F. Skinner in learning theory.(6), 157−159.
[周正怀. (2005a). 桑代克和斯金纳在学习理论上的共通性.,(6), 157−159.]
Zhou, Z. H. (2005b). The difference in learning theory between Thorndike and Skinner.(3), 34−37.
[周正怀. (2005b). 桑代克和斯金纳在学习理论上的分歧.(3), 34−37.]
Zhu, Y. (2019). Ethics in psychological research.(02), 99−108.
[朱滢. (2019). 心理学研究中的伦理.,(02), 99−108.]
Zhu, Y., He, W., & Wang, Y. F. (2017). Challenge-hindrance stress and academic achievement: Proactive personality as moderator., 4(3), 441−452.
An animal behavioral model for the concept of “Integrative Learning”
YIN Bin, WU Xiaorui, LIAN Rong
(School of Psychology, Fujian Normal University, Fuzhou 350108, China)
The dominant paradigm for learning in China today is “gradual learning”, that is, learners acquire knowledge gradually from a lower to a higher level with the help of teachers. Based on theories of adaptive learning and “meta-learning self”, we advanced the alternative of “integrative learning”, that is, “under the role of ‘meta-learning self’, learners actively integrate learning materials to achieve rapid and in-depth understanding of knowledge.” Furthermore, we designed an animal behavioral model to explore the effects of integrative learning versus progressive learning.
Forty SD rats were selected as subjects, a two (Learning mode: Integrative Learning-IL, Progressive Learning - PL) by two (Sex: Male, Female) factorial design was employed, and a fourteen-unit integrative T-maze was constructed for the study. Five task stages were conceived to test the phenomenon and mechanisms of integrative learning: a learning stage, a retest stage after one week, a Gestalt transfer learning stage, a generalization/analysis test stage, and a segment fixation test stage.
The results showed that: 1. During the learning stage, the number of errors in each trial in the IL group decreased exponentially over time, while that curve in the PL group was wavy; males exhibited significantly fewer errors in total than females; and the number of days to learning success in the IL-male group was significantly less than in the PL-male group, though the difference between female groups was not significant. 2. During both Gestalt transfer learning and generalization/analysis test stages, the IL group performed better than the PL group overall; during the segment fixation test stage, all groups appeared fixed more on the first segment of the original correct path. 3. To identify mechanisms for the IL groups’ better performance, a dynamic heat-map path analysis was employed, showing that the IL group (especially males) appeared to consolidate the first key segment of the correct path repeatedly before quickly apprehending the rest of it, which had elements similar to the first one. Males in the PL group, however, were more likely to return to explore the earlier segment than females when allowed to enter a new segment of the maze. 4. The IL group as a whole either ate less of the chocolate reward at the finish of the correct path or moved the pellet elsewhere to eat, a pattern that was much more obvious in females.
We arrived at the following conclusions: 1) Integrative learning is more efficient than progressive learning, and is characterized by the acquisition of more layered knowledge which can better assist long-term migration learning. 2) During the process of forming a “cognitive map”, information stored in memory has the characteristics of entirety, chunking, and categorization. 3) In a maze learning task, performance among males is more consistent than among females. 4) Some individuals may appear anxious or maladjusted during integrative learning.
integrative learning, animal behavioral model, maze test
附录:
S1 有关研究细节
S1.1 实验控制处理
正式实验前, 主试以笼为单位将大鼠取出, 进行15分钟的触摸互动, 持续一周。其目的在于帮助大鼠减轻压力和紧张, 与笼内物理环境、同笼大鼠以及主试建立安全的互动关系。实验前一天控制喂食量为每笼25 g (平均每只5 g, 处于半饥饿状态); 正式实验时, 在单笼实验结束后依次喂食, 保证当天较早完成实验的大鼠在次日实验时间段已处于半饥饿状态; 实验间歇期, 喂食时间固定, 每笼50 g (平均每只10 g, 处于近饱食状态); 实验期间, 大鼠体重保持在自由喂食体重的85%以上; 对个别体重增长缓慢或者下降较为严重的大鼠, 待当天实验结束后, 适当增加喂食量, 并分单笼喂食避免进食竞争, 并在笼下放置温控为33~40℃的专用发热毯进行供暖, 保证其体重达到正常增长范围。在概括分析测验第3天“PL-Male-6号鼠”由于体重过轻进行了单笼喂养, 进食缓慢, 无力, 次日死亡, 妥善处理后移出后续分析。学习阶段, 雄性(89.88 ± 2.04 g)和雌性(89.27 ± 2.04 g)的体重差异不显著((1, 36) = 0.04,= 0.840, η2= 0.00), 整合组(87.16 ± 2.04 g)和渐进组(91.99 ± 2.04 g)的体重差异不显著((1, 36) = 2.834,= 0.100, η2= 0.072)。
实验过程中, 主试每日记录每只大鼠的体重和饮水量。每3~4日使用酒精和无纺布清洁饲养箱、更换垫料、更换大鼠饮用水(净化器过滤的净化纯净水)、清洗引用水瓶。
参与实验的大鼠饲养在福建师范大学动物心理与行为实验室(EVC环境), 室内恒温22.5℃。每日控制12:12的光照循环(使用定时开关和遮光帘, 隔绝自然光照, 控制光照循环)保证大鼠生理节律正常(大鼠有昼伏夜出的习性, 而实验在白天进行。为了保证大鼠在实验时处于清醒状态, 实验室人工控制了12:12的昼夜光照循环)。同时, 空气净化机调节室内空气质量、吊顶监控摄像头记录实验室24小时的内部状况。
S1.2 实验材料
S1.2.1 实验动物的日常养护
9个专用饮水瓶、8个中号饲养笼(笼盖设置有喂食和放置饮水瓶功能、箱体上标注学习方式和性别信息以及饲养负责人的姓名和联系方式)、2个小号饲养笼(用来更换垫料时暂时安放大鼠、对体重轻的大鼠进行单独喂食)、垫料(木屑、玉米芯)、大鼠专用饲料、四层高的置物架、记号笔、称重器(以克为单位)、3个不锈钢器皿(2小, 1大, 称量大鼠和食物)、平板车(运送供应垫料和饲料)、立式台灯和开关定时器、加热垫和温控调节器、黄色标准医用垃圾箱、拖把、扫帚、簸箕、标识有医用垃圾的垃圾袋若干。
S1.2.3 实验装置材料
迷宫制作的原材料包括:4.7 mm厚单面磨砂的亚克力板若干(250 × 250 mm2:31块、125 × 250 mm2:27块、625 × 250 mm2:14块、510 × 135 mm2:1块、500 × 250 mm2:1块、130 × 635 mm2:14块、135 × 260 mm2:1块、250 × 10 mm2:55块)、50 mm宽的白色亚克力无痕接缝胶条(用于接合迷宫拼接缝隙)、500 g亚克力粘合剂(附带注射器)、2卷45 mm宽的黑色强力布基胶带(用于拼装14个单元的组合T迷宫)、1.5 × 1.6 m2亚克力黑色摇粒绒布料(自主剪裁为250 × 140 mm2不透光且双面材质相同的长布条, 控制大鼠记忆窗帘的特殊信息)、19 mm宽的黑色小号长尾夹(用于固定迷宫中的窗帘)、0.3 mm粗优质镀锌铁丝(用于悬挂迷宫中的窗帘)、3 × 3 m2的深灰色地垫。
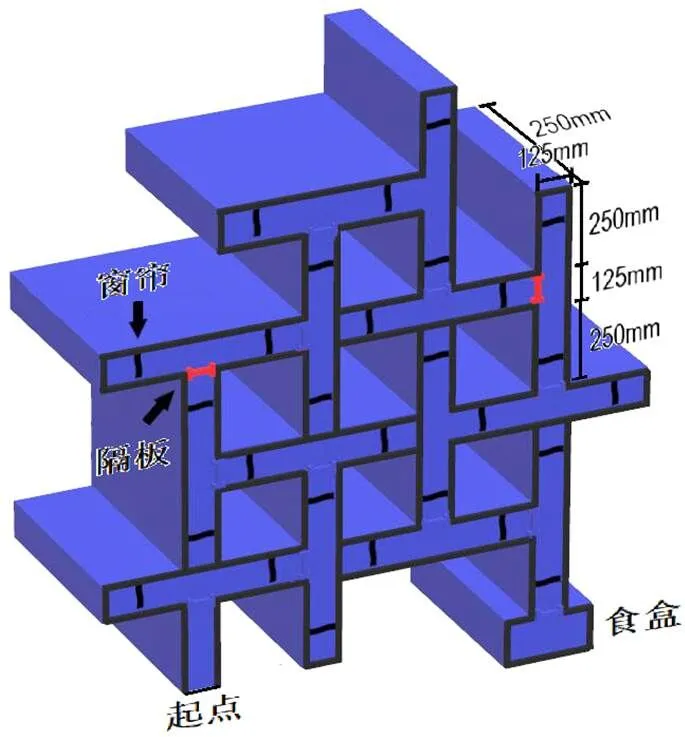
附图1 整合性迷宫实验装置设计图
注:包括窗帘, 隔板, 食盒(食盒内左侧末端配套有一个长方形的黑色系瓷碗)3种5个实验环节皆会用到的基本实验装置。
S1.2.4 清洁防护
主试实验用具:一次性头套、脚套、口罩、手套、实验服; 酒精洗手液、碘伏、棉球、棉签、镊子等擦刮伤急救包;
被试实验用具:无纺布(也称除尘纸)、75%酒精喷剂(除味)。
S1.2.5 其他软硬件材料
动物饲养常规设备(EVC级)、延时供电设备(应急保障)、海康威视(HikVision)硬盘录像机监控设备(4路DS-7104N-F1/4P)、海康威视(HikVision)摄像头(DS-IPC-T12-I/POE)、适配U盘、移动硬盘、立式台灯和开关定时器、加热垫和温控调节器等。
软件包括格式工厂、Camtasia 9(TechSmith)、Supermaze动物行为视频分析系统(上海欣软)、GraphPad Prism 8、Microsoft Excel 2017、SPSS 18.0、3D画图软件等。
S1.3 实验范式设计
整合性学习实验的动物行为迷宫设计共有5个模块。迷宫的轮廓尺寸为标准尺寸, 分段隔板、卡槽设计以及窗帘制作都围绕整合性学习的研究需要进行了自主设计。
学习阶段中, 渐进组的分段依据路径的长度(三段路径的长度一致)和路径的规律迁移特性(三段路径的形状相似)分割为3段; 整合组全段开放。实验装置以整合性学习和渐进性学习方式的操作性定义为指导进行设计(见附图2)。一周后复测任务测试大鼠对于迷宫路径形成的长时记忆水平, 考察不同学习方式在长时记忆上的差异, 即记忆信息的牢固程度(见附图2); 格式塔迁移任务是基于大鼠对迷宫正确路径记忆的“整体性”设计的测试, 考察大鼠对路段记忆的联结(附图3); 概括分析测验是对大鼠绝对方位认知的测试, 考察大鼠对路段的加工层次和深度(附图4); 阶段固着测验是对渐进组大鼠是否由于分段提示信息的产生相对方位认知上的优势的测验, 同时, 也可以显化大鼠对路段间迁移信息的使用效果和使用方式(附图5)。
S1.4 研究程序
(1)设计和制作合适尺寸的14个单元的组合T迷宫;

附图2 整合性学习实验学习阶段和复测阶段的动物行为迷宫设计图
注:红色隔板只供渐进组学习阶段使用(隔板放置处有凹槽, 可抽插)。黄、红、绿路线分别是渐进组1~3天、4~7天、8~12天的正确行径路线, 绿路线也是整合组的正确行径路线。复测时, 整合组和渐进组饥皆开放迷宫的隔板施测

附图3 整合性学习实验格式塔迁移任务的动物行为迷宫设计图
注:“食盒”入口被隔板密封, 变更为“起点”, 起点的窗帘移至“终点”岔路口处。

附图4 整合性学习实验概括分析测验的动物行为迷宫设计图
注:左图黄、红、绿分别为最短路线, 原路线和隐蔽路线(隔板拆卸在错误区域的窗帘后)是1~5天的概括与分析测试路线。

附图5 整合性学习实验阶段固化的动物行为迷宫设计图
注:黄、绿、蓝分别为第一段、第二段、第三段路径的最短路径, 是1~4天的阶段固化测试路线
(2)使用12只大鼠进行了预实验, 制定实验步骤、观察记录表、量化分析指标、实验操作要领、排除系统干扰条件;
(3)采购40只1月大的大鼠后分笼, 实验前连续7天, 对大鼠进行每日15分钟的触摸互动。投放足量食物(50 g每笼), 记录其体重、饮水量和剩余食物量, 作为正式实验时的喂食量参考;
(4)正式实验按照整合雄1、渐进雄1、整合雌1、渐进雌2、整合雄2、渐进雄2、整合雌2、渐进雌2的笼号顺序进行, 每笼实验结束后喂食该笼25 g食物, 保证次日实验中笼间的半饥饿水平一致;
(5)正式实验前, 在终点处放置装有食物(大鼠更喜欢牛奶巧克力豆, 但由于大鼠之前未食用过巧克力豆, 因此碗中开始时放置1小粒日常食用的饲料和1颗巧克力豆。在所有大鼠都开始食用巧克力豆后, 只放置1粒巧克力豆)的方形黑色瓷碗(瓷碗有一定程度的提示功能, 提示大鼠食物位置的改变, 以及预防因忘记投食而影响大鼠后续的探索行为, 因为瓷碗作为强化物仍有一定的强化功能)。实验前, 先对大鼠称重, 然后单手将手臂伸长将大鼠放入起点, 身体靠后远离迷宫, 避免身体对摄像镜头的遮挡而无法收集有效的初始数据; 实验时, 人工记录大鼠的特殊行为, 例如:是否在原地吃食物, 是否吃了巧克力;
(6)实验后使用酒精喷洒迷宫壁并用无纺布酌情擦拭, 同时, 调试窗帘, 避免前一只大鼠的气味(屎、尿、毛发)和造成窗帘下垂纹路的变化对后一只大鼠的影响;
(7)每日实验后备份当日原始视频, 裁剪成可分析的标准格式, 存入移动硬盘;
(8)学习阶段进行12日。1~3日渐进组在第一段终点放置隔板, 学习3日; 4~7日渐进组第一段隔板移除, 在第二段终点放置隔板, 学习4日; 8~12日渐进组第二段隔板移除学习5日; 整合组12日全段不放置隔板。其中, 学习阶段的第一日, 每只大鼠定时学习15分钟(保证绝大多数大鼠对迷宫产生适应后的探索行为并吃到食物, 也平衡渐进组因为路段短, 偶然到达终点而造成的探索时间不足), 2~12日以大鼠到达终点吃完食物(等待大鼠吃完再取出, 避免对其走迷宫行为动机产生影响)为取出迷宫中大鼠;
(9)停测1周, 每笼每日喂食50 g, 记录体重和饮水数据。期间, 处理12天的学习阶段的视频。将视频放入SuperMaze系统进行批量分析, 第1天虽然每只大鼠都探索了15分钟, 但视频分析时, 触发器会自动截取其第一次到达终点前的视频数据进行分析, 一周后复测阶段中, 整合组和渐进组完成相同的整段迷宫的测试;
(10)格式塔迁移测试时, 将食槽口用隔板密封, 终点与起点互换, 终点前的窗帘移动至起点前, 将盛有食物的食碟放在新的终点(原起点), 施测3天;
(11)按学习阶段路径重新学习一天(减少逆向迁移测验对概括分析测验的影响), 然后进行概括分析测验5天;
(12)按学习阶段路径重新学习一天(减少概括分析测验对阶段固化测验的影响), 然后进行阶段固化测验4天。
(13)使用SuperMaze系统进行批量分析, 使用SuperMaze中的轨迹图复查数据, 对部分由于视频片段不完整或缀余的视频进行了二次备份和裁剪, 保证数据的质量。使用Excel、SPSS 18.0对数据进行整理, 使用GraphPad Prism 8进行数据分析和图表制作。
S1.5 数据处理详细步骤
首先, 使用连接吊顶固定摄像头的监控设备对1113次(其中“PL-Male-6”的后三次的概括分析任务和全部四次的阶段固化测验空缺)大鼠跑迷宫的试次进行视频记录和缓存, 将有效视频片段以AVI格式导出。随后, 使用格式工厂和TechSmith Camtasia Studio 9进行原始视频的裁剪和格式化, 生成裁剪掉无关信息的640 : 480的屏幕大小, Xvid格式的单只大鼠视频。
然后, 使用《SuperMaze动物行为视频分析系统》(上海欣软信息科技有限公司)创建分析平台(见附图6)。其中, 设置动态背景算法、3点(头部、重心、尾部)跟踪位置、大鼠亮度和背景亮度对比为较亮、工作模式为先腐蚀后膨胀(参数:2像素, 主要用于消除小物体、在纤细点处分离物体、平滑较大物体的边界)、面积阈值启用头部、重心判断(只有大鼠将头部和重心皆进入指定区域才记录进入)、头部探索时间阈值为5×100 ms、测试参数中延时结束为900 s、触发器设置为“首次进入区域Foodbox (IL组一直设置为Foodbox; PL组设置1~3天为First segment end、4~7天为Second segment end、8~12天为Foodbox)并停留1 s”。后4个任务的测验只改变分析区域设置。设置后在“采集分析”中使用“批量处理录像”功能对1113个视频进行动态分析。结束分析后在“数据管理”中导出原始数据。

附图6 Supermaze动物行为视频分析系统中的实验设计——区域设置
最后, 使用SuperMaze分析系统中的轨迹图复查数据, 对部分由于视频片段不完整或缀余的视频进行了二次备份和裁剪, 保证数据的质量。使用Excel、SPSS18.0对数据进行整理, 使用GraphPad Prism8进行数据分析和图表制作。
S2 实验结果统计分析表格及附图
研究原始数据已上传至百度云盘可供查用。
链接:https://pan.baidu.com/s/1fWtfjPH1Tnekj2OxzDcz_Q
提取码:geag
附表1 学习阶段的错误次数三因素重复测量方差分析表

SSdfMSF (dfn, dfd)pη2 学习次数3090.7211280.00F (11, 396) = 29.42***0.0000.45 学习方式141.921141.92F (1, 36) = 15.54***0.0000.30 性别105.471105.47F (1, 36) = 11.55**0.0020.24 学习次数 × 学习方式2940.8111267.35F (11, 396) = 27.99***0.0000.44 学习次数 × 性别257.461123.41F (11, 396) = 2.45**0.0060.06 学习方式 × 性别79.22179.22F(1, 36) = 8.67**0.0060.19 学习次数 × 学习方式 × 性别1911117.36F (11, 396) = 1.82*0.0480.05
注:*< 0.05, **< 0.01, ***< 0.001。
附表2 学习成功天数的两因素完全随机方差分析结果
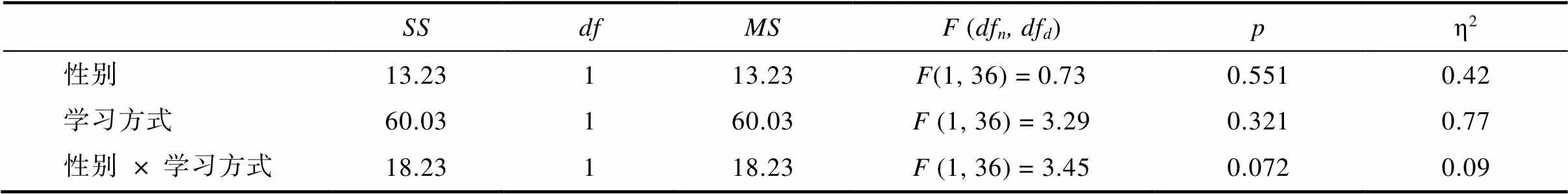
SSdfMSF (dfn, dfd)pη2 性别13.23113.23F(1, 36) = 0.730.5510.42 学习方式60.03160.03F (1, 36) = 3.290.3210.77 性别× 学习方式18.23118.23F (1, 36) = 3.450.0720.09
附表3 学习成功天数的事后多重比较结果

SSdfMSF (dfn, dfd)pη2 性别13.23113.23F(1, 36) = 2.500.1220.07 学习方式60.03160.03F (1, 36) = 11.36**0.0020.24
注:**< 0.01。
附表4 性别水平上学习方式的简单效应比较

tpCohen’s d 雄性4.12***0.000−1.84 雌性0.980.341−0.44
注:***< 0.001。
附表5 一周后复测任务错误次数的三因素重复测量方差分析表

SSdfMSF (dfn, dfd)pη2 学习次数3.8221.91F (2, 72) = 2.190.1200.06 学习方式10.21110.21F(1, 36) = 4.030.0520.10 性别3.0113.01F(1, 36) = 1.190.2830.03 学习次数 × 学习方式3.0221.51F(2, 72) = 1.730.1850.05 学习次数 × 性别0.5220.26F(2, 72) = 0.300.7450.01 学习方式 × 性别0.0110.01F (1, 36) = 0.000.9550.00 学习次数 × 学习方式 × 性别0.5220.26F (2, 72) = 0.300.7450.01
附表6 格式塔迁移任务错误次数的三因素重复测量方差分析表
SSdfMSF (dfn, dfd)pη2 学习次数1071.552535.28F (2, 72) = 25.64***0.0000.42 学习方式130.211130.21F (1, 36) = 5.50*0.0250.13 性别118.011118.01F (1, 36) = 4.99*0.0320.12 学习次数 × 学习方式1.7220.86F (2, 72) = 0.040.9600.00 学习次数 × 性别21.72210.86F (2, 72) = 0.520.5970.01 学习方式 × 性别2.4112.41F (1, 36) = 0.100.7520.00 学习次数 × 学习方式 ×性别5.0222.51F (2, 72) = 0.120.8870.00
注:*< 0.05, ***< 0.001。
附表7 五天概括分析任务的三因素重复测量方差分析

SSdfMSF (dfn, dfd)pη2 测试次数24.5446.13F(4, 140) = 3.00*0.0200.08 学习方式12.60112.60F(1, 35) = 4.66*0.0380.12 性别3.5813.58F(1, 35) = 1.320.2580.04 测试次数× 学习方式9.1042.28F(4, 140) = 1.110.3520.03 测试次数× 性别2.2340.56F(4, 140) = 0.270.8950.01 学习方式× 性别4.1214.12F(1, 35) = 1.530.2250.04 测试次数× 组别× 性别6.8841.72F(4, 140) = 0.840.5000.02
注:*< 0.05。
附表8 阶段固着末次测验依然选择原路线大鼠比例的三因素重复测量方差分析

SSdfMSF (dfn, dfd)pη2 分段路径9.7024.85F (2, 70) = 40.83***0.0000.54 学习方式0.2010.20F (1, 35) = 1.180.2860.03 性别0.0710.07F (1, 35) = 0.420.5200.01 学习方式× 性别0.0110.01F (1, 35) = 0.050.8300.00 分段路径× 学习方式0.5520.28F (2, 70) = 2.320.1060.06 分段路径× 性别0.2920.15F (2, 70) = 1.220.3000.03 分段路径× 学习方式× 性别0.3620.18F (2, 70) = 1.500.2310.04
注:***< 0.001。
附表9 IL组分段路径的行进路程的两因素重复测量方差分析

SSdfMSF (dfn, dfd)pη2 分段路径2987.8721493.94F (2, 36) = 38.01***0.0000.68 性别3208.6313208.63F (1, 18) = 22.51***0.0000.56 分段路径× 性别492.362246.18F(2, 36) = 6.26**0.0050.26
注:**< 0.01, ***< 0.001。
附表10 IL组分段路径的行进时间的两因素重复测量方差分析

SSdfMSF (dfn, dfd)pη2 分段路径111308.10255654.05F (2, 36) = 39.10***0.0000.69 性别67002.42167002.42F(1, 18) = 22.10***0.0000.55 分段路径×性别25910.57212955.29F (2, 36) = 9.10**0.0010.34
注: **< 0.01, ***< 0.001。
附表11 PL组第二阶段路程的三因素重复测量方差分析
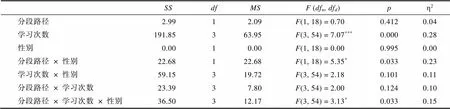
SSdfMSF (dfn, dfd)pη2 分段路径2.9912.09F(1, 18) = 0.700.4120.04 学习次数191.85363.95F(3, 54) = 7.07***0.0000.28 性别0.0010.00F(1, 18) = 0.000.9950.00 分段路径× 性别22.68122.68F(1, 18) = 5.35*0.0330.23 学习次数× 性别59.15319.72F(3, 54) = 2.180.1010.11 分段路径× 学习次数23.3937.80F(3, 54) = 2.000.1240.10 分段路径× 学习次数× 性别36.50312.17F(3, 54) = 3.13*0.0330.15
注: **< 0.01, ***< 0.001。
附表12 PL组第二阶段时间的三因素重复测量方差分析

SSdfMSF (dfn, dfd)pη2 分段路径137.601137.60F(1, 18) = 0.130.7220.01 学习次数34073.62311357.87F(3, 54) = 8.00***0.0000.31 性别549.601549.60F(1, 18) = 0.300.5890.02 分段路径× 性别4361.6214361.62F(1, 18) = 4.140.0570.19 学习次数× 性别3598.0131199.34F(3, 54) = 0.850.4750.05 分段路径 × 学习次数2163.393721.13F(3, 54) = 0.810.4960.04 分段路径× 学习次数× 性别2471.073823.69F(3, 54) = 0.920.4370.05
注:***< 0.001。
附表13 PL组第三阶段路程的三因素重复测量方差分析

SSdfMSF (dfn, dfd)pη2 分段路径29.19214.60F(2, 36) = 15.86***0.0000.47 学习次数48.08412.02F(4, 72) = 5.53**0.0010.24 性别0.6910.69F(1, 18) = 0.430.5230.02 分段路径× 性别4.7422.37F(2, 36) = 2.570.0900.13 学习次数× 性别5.5341.38F(4, 72) = 0.640.6380.03 分段路径× 学习次数15.8581.98F(8, 144) = 2.33*0.0220.11 分段路径× 学习次数× 性别2.1080.26F(8, 144) = 0.310.9620.02
注:*< 0.05, **< 0.01, ***< 0.001。
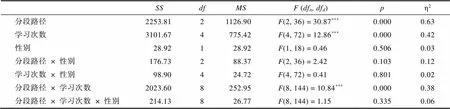
附表14 PL组第三阶段路程的三因素重复测量方差分析
SSdfMSF (dfn, dfd)pη2 分段路径2253.8121126.90F(2, 36) = 30.87***0.0000.63 学习次数3101.674775.42F(4, 72) = 12.86***0.0000.42 性别28.92128.92F(1, 18) = 0.460.5060.03 分段路径× 性别176.73288.37F(2, 36) = 2.420.1030.12 学习次数× 性别98.90424.72F(4, 72) = 0.410.8010.02 分段路径× 学习次数2023.608252.95F(8, 144) = 10.84***0.0000.38 分段路径× 学习次数× 性别214.13826.77F(8, 144) = 1.150.3350.06
注:***< 0.001。
附表15 关于学习阶段中学习方式适应性的三因素重复测量方差分析中的被试间结果

SSdfMSF (dfn, dfd)pη2 体重学习方式2793.6812793.68F (1, 36) = 2.800.1030.07 性别44.41144.41F (1, 36) = 0.040.8340.00 学习方式×性别1387.2011387.20F (1, 36) = 1.390.2460.37 起点静止时间学习方式7.3117.31F (1, 36) = 0.640.4290.02 性别25.60125.60F (1, 36) = 2.240.1430.06 学习方式× 性别28.10128.10F(1, 36) = 2.460.1260.06 原地进食学习方式8.0118.01F (1, 34) = 15.93***0.0000.32 性别0.7810.78F (1, 34) = 1.550.2220.04 学习方式× 性别1.4711.47F (1, 34) = 2.920.0970.08 进食巧克力学习方式0.8310.83F (1, 36) = 5.07*0.0310.12 性别0.4110.41F (1, 36) = 2.490.1240.07 学习方式× 性别0.6810.68F (1, 36) = 4.110.0500.10
注:三因素包括学习方式、性别和学习次数, 各组在终点原地进食或进食巧克力的比例随学习次数变化的情况可见附图11&12; 是否原地进食的原始数据为学习阶段2~12次的数据, 因为学习阶段第1次学习为限时15分钟的探索, 未记录是否在原地进食的行为, 其余指标皆为学习阶段1~12次的数据。各指标统计了学习阶段的总比例或均值。*< 0.05, ***< 0.001。
附图7 PL组第4~7天第一路径和第二路径行进时间的性别对比图

附图8 PL组第4~7天第一路径和第二路径行进路程的性别对比图
附图9 PL组第8~12天第一、二、三路径行进时间的性别对比图

附图10 PL组第8~12天第一、二、三路径行进路程的性别对比图
附图11 各组在终点原地进食比例随学习次数增加的变化
附图12 各组在终点吃掉巧克力比例随学习次数增加的变化
S3 实验视频示例
研究中比较有代表性的7个实验视频下载链接如下:
链接:https://pan.baidu.com/s/1fWtfjPH1Tnekj2OxzDcz_Q
提取码:geag
视频1:学习任务完成示例(IL-Male-2学习任务第12天)
说明:学习任务第12天(最后一天), 大部分被试均可零错误的完成迷宫任务, 少量被试有1至4次错误, 组间错误次数分布呈显著差异(F = 5.128, p = 0.03)。其中IL-Male组所有被试均无错误; IL-Female组有1只错了4次, 其余均无错误; PL-Male组有3只错了1次, 1只错了3次, 其余均无错误; PL-Female组有4只错了1次, 1只错了2次, 2只错了3次, 其余均无错误。
视频2:格式塔迁移任务完成示例(IL-Male-1格式塔迁移任务第3天)
说明:格式塔迁移任务第3天(最后一天), 大部分被试均可以较少错误或零错误完成任务, 少部分被试错误较多, 组间错误次数分布呈显著差异(表5)。其中IL-Male组有5只错了1次, 其余均无错误; IL-Female组有3只错了1次, 1只错了2次, 1只错了4次, 1只错了5次, 1只错了11次, 其余均无错误; PL-Male组有4只错了1次, 1只错了2次, 1只错了3次, 1只错了4次, 其余均无错误; PL-Female组有1只错了1次, 1只错了2次, 1只错了3次, 2只错了6次, 1只错了7次, 1只错了8次, 1只错了11次, 其余均无错误。
视频3:学习过程中IL组重点学习第一路径示例(IL-Male-5学习阶段第2天)
说明:IL-Male-5在第一路径多次折回探索, 而在第二、三路径错误次数较少的通过。迷宫路径的划分如下图所示。数据结果如正文中图5所示。
注:将基础实验装置的正确路径划分为具有形状相似性的三段, 红、绿、蓝依次为第一路径、第二路径和第三路径。
视频4:学习过程中PL-Male组在新路段开放后着重探索旧路径示例(PL-Male-9学习阶段第4天)
视频5:学习过程中PL-Female组在新路段开放后着重探索新路径示例(PL-Female-9学习阶段第4天)
视频4&5说明:在新路段开放后, PL-Male组在原路段终点处折回, 重新探索第一路径, 后进入第二路径到达终点; PL-Female组则直接选择进入第二路径, 而后在第二路径进行了折回探索并到达终点。PL组三段路径逐步开放示意图如下。数据结果如正文中图6所示。
视频6:概括分析任务各组5次测验所有轨迹图的快速呈现。
视频6说明:
·我们将概括分析任务5次测验(学习)的全部被试的轨迹图的导出过程用录频软件进行了录屏, 并区分出了不同组别进行连续播放。
·视频说明:播放顺序为先IL, 后PL; 先雄, 后雌; 并按单只大鼠1~5天的顺序进行循环(共40个被试)。视频左下角的标注“IL-Male1-1”即“整合雄组1号大鼠第一次概括分析任务测试”;
·规律说明:IL组多数选择最短路线或原路线; PL组多数出现无明显规律的全盘搜索。
·概括分析任务示意图如下。数据结果如正文中图4a, 4b所示。
注:红色隔板只供PL组学习阶段使用。黄、红、绿路线分别是PL组1~3天、4~7天、8~12天的正确行径路线, PL组的学习路线依据迷宫的长度和规律用隔板分为3段(隔板放置处有凹槽, 可插拔)。
注:概括分析任务中黄、红、绿分别为最短路线, 原路线和隐蔽路线(隔板拆卸在错误区域的窗帘后), 是1~5天的概括与分析测试路线。
视频7:阶段固着测验中被试对第一原路径惯性固着示例(IL-Male-10第四次测试)
视频7说明:在阶段固着测验的第四次测试中, 各组仍一致的选择在第一路径选择原路线, 而在其他两个路径更多选择短路线。阶段固着任务示意图如下。数据结果如正文中图4c, 4d所示。
注:阶段固着任务中, 黄、绿、蓝分别为第一、第二、第三路径的最短路径, 原路径仍保持通畅。
分类号 B845
收稿日期: 2020-05-05
* 教育部人文社会科学重点研究重大项目《学生心理健康促进体系与服务平台建设》(16JJD190004); 福建师范大学人事处“海外引进人才−青年英才”科研启动项目(Z0210509)。
通信作者: 连榕, E-mail: lianrong1122@126.com
猜你喜欢
商用汽车(2021年4期)2021-10-13
作文周刊·小学一年级版(2021年36期)2021-01-14
阅读与作文(小学高年级版)(2020年8期)2020-09-12
语数外学习·高中版上旬(2020年5期)2020-09-10
小学生学习指导(低年级)(2018年11期)2018-12-03
中学生数理化(高中版.高一使用)(2018年1期)2018-02-10
文理导航·科普童话(2016年7期)2017-02-04
理科考试研究·高中(2016年10期)2017-01-17
文理导航·科普童话(2016年4期)2016-05-31
儿童故事画报·智力大王(2015年12期)2016-01-23
