
太行鸡VIPR-1基因表达特点及其功能分析
2020-11-12 07:09:52周祖阳张辉张清阳白莹杨俊琦刘荐男李凯扬刘玉芳
畜牧与兽医 2020年11期
周祖阳,张辉,张清阳,白莹,杨俊琦,刘荐男,李凯扬,刘玉芳*
(1.河北工程大学生命科学与食品工程学院,河北 邯郸 056001;2.河北工程大学园林与生态工程学院,河北 邯郸 056001;3.北京市畜牧总站,北京 100107)
地方鸡品种与商业化蛋鸡品种相比,尽管鸡蛋具有蛋黄颜色深、口感好等特点,但因其产蛋量低而严重制约了地方鸡品种的推广和发展[1]。蛋鸡的产蛋性能受到多个基因的调控,查阅前人研究发现,血管活性肠肽受体-1(vasoactive intestinal peptide receptor-1,VIPR-1)是VIP蛋白受体家族成员之一,属于Ⅱ类亚家族。有文献报道,VIPR-1基因在下丘脑和垂体中均有表达,但主要在垂体中表达,并且只有垂体中VIPR-1基因的mRNA差异表达与生殖功能改变相关,VIPR-1基因已被证实参与禽类的育雏调节[2]。与其他家族成员结构相似,VIPR-1是一种糖蛋白,具有大的亲水性细胞外N端和7个高度保守的疏水性跨膜螺旋以及细胞质C端[3]。辛清武等[4-5]研究发现VIPR-1基因C902T突变和A988G突变与黑番鸭的就巢性状显著相关,能够一定程度上提高黑番鸭的产蛋性能。在雌性鹌鹑的研究中发现,VIPR-1基因的G373T和A313G两个突变位点可用于选择产蛋量高或蛋重大的鹌鹑品系,此外,VIPR-1基因还可以作为分子标记辅助鹌鹑生长性状选择的遗传标记[6]。在宁都三黄鸡的研究中也发现,VIPR-1基因中的C1704887T和C175301T两个突变位点可能是家禽高产蛋量的重要分子标记[7]。位于VIPR-1基因内含子6的C1704887T与鸡300日龄的产蛋数有显著相关,基于VIPR-1基因C1704887T和C175301T的单倍型分析也证实了这一结果[8]。另一研究分析了VIPR-1基因的突变位点A1661691G与蛋鸡初产日龄相关[9]。这些研究表明,VIPR-1基因的分子生物学功能与家禽产蛋性状密切相关,而太行鸡VIPR-1基因与产蛋性能的研究较少。
本研究选择高产组和低产组太行鸡卵巢组织作为研究对象,经过PCR扩增、克隆VIPR-1基因序列并进行测序和结构分析,利用半定量RT-PCR和实时荧光定量PCR技术探讨VIPR-1在太行鸡260日龄产蛋阶段的表达情况,为后期进一步探究VIPR-1基因的分子生物学功能及深入研究VIPR-1基因与蛋鸡产蛋性能的关系奠定基础。
1 材料与方法
1.1 材料
选取260日龄健康、产蛋稳定、羽色良好、体重相近的太行鸡母鸡250只进行标号,从蛋鸡开产日开始,每日下午3时记录每只母鸡的产蛋情况,统计产蛋鸡260日龄内的产蛋量,将鸡群分为高产(H)(产蛋量≥40枚,平均产蛋量(48.33±0.40)枚)和低产(L)(产蛋量<40枚,平均产蛋量(28.12±0.40)枚两组,随后进行翅静脉采集血液样本并注明标号,-20 ℃保存。从高、低产蛋组中选取体态均匀、健康母鸡各10只,屠宰后快速采集心、肝、脾、肺、肾、卵巢组织等样品,液氮速冻后-80 ℃保存。
1.2 方法
1.2.1 太行鸡血样DNA提取
取20 μL血样,使用血液DNA提取试剂盒(北京天根生物科技有限公司)进行DNA提取。取出2 μL DNA与上样Buffer混匀,用1.0%琼脂糖凝胶电泳检测DNA的完整性。利用NanoDrop 2 000检测DNA的浓度,随后放入低温-20 ℃保存。
1.2.2 各组织RNA提取与cDNA合成
分别取约100 mg 心脏、肝脏、脾脏、肺脏、肾脏和卵巢组织,按照RNA提取试剂盒说明书进行总RNA提取,随后进行琼脂糖凝胶电泳和RNA浓度检测。检测合格的RNA样品,根据Fast Quant cDNA 第一链合成试剂盒(北京天根生物科技有限公司)说明书将太行鸡各组织RNA反转录合成cDNA,-20 ℃保存备用。
1.2.3 太行鸡VIPR-1基因克隆、扩增与测序
在GenBank数据库中找到鸡参考基因组中VIPR-1基因的CDS区序列(GenBank登录号:NM_001097523.1),利用Primer premier 5软件设计目的基因(VIPR-1-E1、VIPR-1-E2、VIPR-1-E3、VIPR-1-E4、VIPR-1-E5-6、VIPR-1-E7-8、VIPR-1-E9、VIPR-1-E10、VIPR-1-E11、VIPR-1-E12、VIPR-1-E13)的CDS区引物,引物信息见表1。引物序列由上海生工生物工程股份有限公司合成。PCR反应体系20 μL:2×TaqPCR Mix 10 μL,ddH2O 8 μL,上、下游引物各0.5 μL,cDNA模板1 μL。PCR反应条件:95 ℃预变性5 min;95 ℃变性30 s,退火温度见下表30 s,72 ℃延伸1 min,共34个循环,72 ℃延伸7 min,4 ℃保存。取PCR产物进行琼脂糖凝胶电泳检测,并送至生工生物工程股份有限公司进行测序。

表1 扩增VIPR-1基因CDS区及实时荧光定量PCR引物
1.2.4 太行鸡VIPR-1序列结构分析
测得VIPR-1基因CDS区结果利用 NCBI 网站(http://blast.ncbi.nlm.nih.gov/Blast.cgi)上BLASTP软件进行同源基因的氨基酸序列搜索;开放阅读框(open reading frame,ORF)氨基酸序列分析用NCBI中的ORF Finder(http://www.ncbi.nlm.nih.gov/gorf/gorf.html)工具完成。利用ExPASy网站(http://www.expasy.ch/tools/protparam.html)分析蛋白的理化性质包括相对分子质量、等电点、消光系数、亲水性和疏水性、氨基酸组成等;SOPMA(http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)预测蛋白质的二级结构。
1.2.5 系统进化分析
将获得的序列在DNAMAN 5.0软件上进行分析,并与GenBank上获取的其它物种序列进行同源性比较。利用 MEGA X软件以距离矩阵邻近归并法(NeighborJoining,NJ)建种间系统发生树,通过 bootstrap 法获得系统分支的置信度(重复次数为1 000)。
1.2.6VIPR-1组织表达谱分析
以心脏、肝脏、脾脏、肺脏、肾脏、卵巢组织的cDNA为模板,VIPR-1-Q1-F/R(见表1)为引物,GAPDH为内参基因进行半定量RT-PCR。PCR反应体系为20 μL:2×TaqPCR Mix 10 μL,ddH2O 8 μL,上、下游引物各0.5 μL,cDNA模板1 μL。PCR反应条件:95 ℃预变性5 min;95 ℃变性30 s,56 ℃退火30 s,72 ℃延伸30 s,共31个循环,72 ℃延伸7 min。取PCR产物进行琼脂糖凝胶电泳检测,分析半定量RT-PCR结果。
1.2.7 实时荧光定量PCR分析
我想,我们的暗恋大多无果而终,是因为自己不够优秀吧。“我不想继续开小卖部了,我要奋斗!”沐子她从小卖部撤了资,藏起浮躁的女汉子性格,不再成天把八卦和大话挂在嘴边,虔诚地皱着眉头听老师讲课,活得像一个“出家人”,灵台一片澄澈空明。
以高、低产太行鸡卵巢组织cDNA为模板(每组10只),VIPR-1-Q1-F/R(见表1)为引物,GAPDH为内参基因,采用SYBR Green Ⅰ染料法,进行实时荧光定量PCR,对VIPR-1基因在太行鸡高、低产蛋量个体卵巢组织中的表达情况进行分析。反应体系:Real MasterMix(SYBR Green I)10 μL,ddH2O 8 μL,上、下游引物各0.5 μL,cDNA模板1 μL。反应条件:95 ℃预变性30 s;95 ℃变性30 s,56 ℃退火30 s,72 ℃延伸30 s,共40个循环,72 ℃延伸10 min。采用2-ΔΔCt法计算相对表达量,利用SPSS 18.0统计软件的t检验进行显著性差异分析。
2 结果
2.1 太行鸡VIPR-1基因PCR扩增
DNA提取后,经核酸检测仪检测和琼脂糖电泳合格后,进行PCR扩增。用VIPR-1-E1、VIPR-1-E2、VIPR-1-E3、VIPR-1-E4、VIPR-1-E5-6、VIPR-1-E7-8、VIPR-1-E9、VIPR-1-E10、VIPR-1-E11、VIPR-1-E12和VIPR-1-E13共11对引物进行PCR扩增(每对引物4个梯度),将产物在1.5%的琼脂糖凝胶中进电泳,分别可见目的片段大小的特异性条带(图1),随后送去测序,获得序列进行比对拼接,用于后续基因结构分析。
2.2 生物信息学分析
2.2.1 太行鸡VIPR-1基因序列分析
克隆测序后获得5 942 bp全长的太行鸡VIPR-1基因cDNA序列,此序列包含了长度为1 341 bp的ORF(开放阅读框),编码446个氨基酸。将该序列与参考基因组的VIPR-1基因完整CDS序列进行同源性比较,发现其同源性为100%,推定该序列即为太行鸡VIPR-1基因的完整CDS序列。
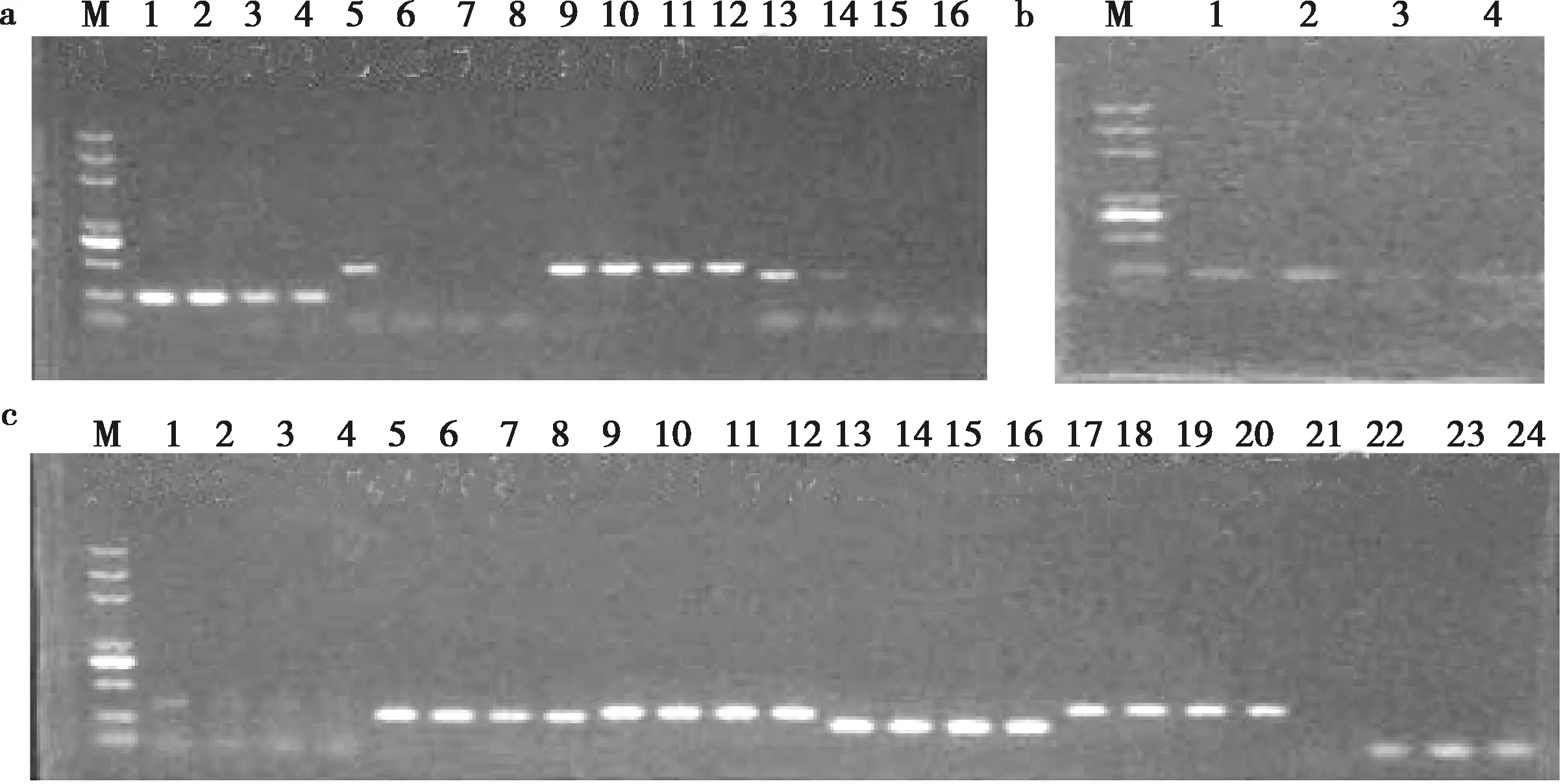
M.DL5000 DNA marker;a.泳道1~4、5~8、9~12、13~16分别代表VIPR-1-E1、VIPR-1-E2、VIPR-1-E3、VIPR-1-E4在4个梯度下的PCR产物电泳;b.泳道1~4代表VIPR-1-E5-6在4个梯度下的PCR产物电泳;c.泳道1~4、5~8、9~12、13~16、17~20、21~24分别代表VIPR-1-E7-8、VIPR-1-E9、VIPR-1-E10、VIPR-1-E11、VIPR-1-E12和VIPR-1-E13在4个梯度下的PCR产物电泳图1 VIPR-1基因梯度PCR产物电泳
2.2.2 分子进化分析
用DNAMAN 5.0 软件进行同源性比较分析,太行鸡VIPR-1基因编码区序列和其他已经报道的禽类VIPR-1基因的同源性较高,其中与野鸭(XM_005023238)的同源性最高为 92.24%;其次为火鸡(U31991)90.11%;与绵羊(XM_027958056)、猪(NM_214036)、牛(NM_001081607)、人(NM_004624)、小鼠(NM_011703)、兔(XM_002713061)、非洲爪蟾(XM_002941864)、沼泽蛙(AF100644)等哺乳物种、两栖类的同源性较低,在64.51%~67.18%。氨基酸序列比较发现,太行鸡的VIPR-1氨基酸序列与野鸭氨基酸一致性高达93.72%,与火鸡的一致性也达到82.93%,与人、犬、牛、猪等哺乳物种的同源性在62.95%~65.50%,与非洲爪蟾、两栖类沼泽蛙的同源性比其他哺乳物种高,分别为74.22%和76.34%。见表2。
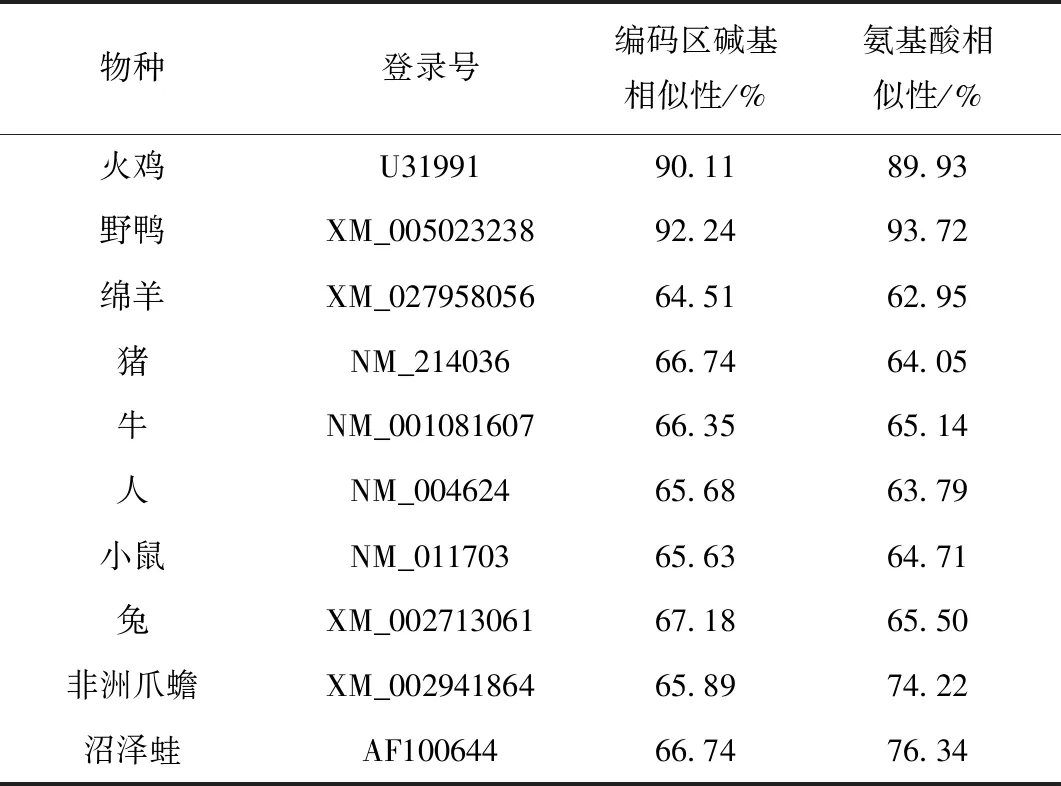
表2 VIPR-1不同物种间同源性比较
2.2.3 N-J系统进化树

图2 太行鸡VIPR-1基因系统发生树
2.2.4 太行鸡VIPR-1蛋白结构预测
利用ORF Finder预测太行鸡VIPR-1基因编码446个氨基酸。太行鸡VIPR-1蛋白特征分析显示,该蛋白分子质量为112 620.03 Da,理论等电点为4.98,280 nm的摩尔消光系数为172 375 moL中含量-1;该蛋白质包含了20 种常见的氨基酸,其氨基酸含量中色氨酸最低为1.9%,亮氨酸和丝氨酸含量高分别为10.9%和10.6%,其他氨基酸的含量在2.0~8.0之间,酸性氨基酸和碱性氨基酸含量较低,分别为7.1%和16.6%,带电荷的氨基酸和疏水氨基酸的含量分别为21%和36%;精氨酸(Arg)疏水性最强(最高分值为4.5),异亮氨酸(Ile)亲水性最强(最低分值为-4.5);疏水性预测显示太行鸡 VIPR-1大多数氨基酸为亲水性氨基酸,总平均疏水性为-0.250(图3)。蛋白质二级结构分析发现太行鸡VIPR-1蛋白由α-螺旋、延伸、β-旋折叠及无规则卷曲4种结构组成,所占比例分别为30.1%、21.12%、7.85%和40.94%。
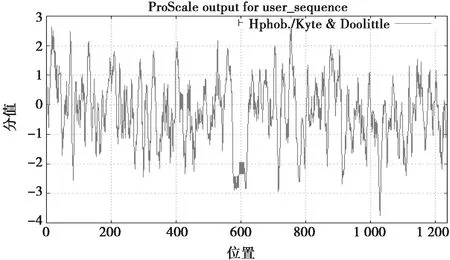
图3 太行鸡VIPR-1蛋白疏水性分析
2.3 组织半定量RT-PCR表达谱分析
VIPR-1基因在太行鸡心脏、肝脏、脾脏、肺脏、肾脏和卵巢中均表达,但在卵巢中表达量最高(图4)。
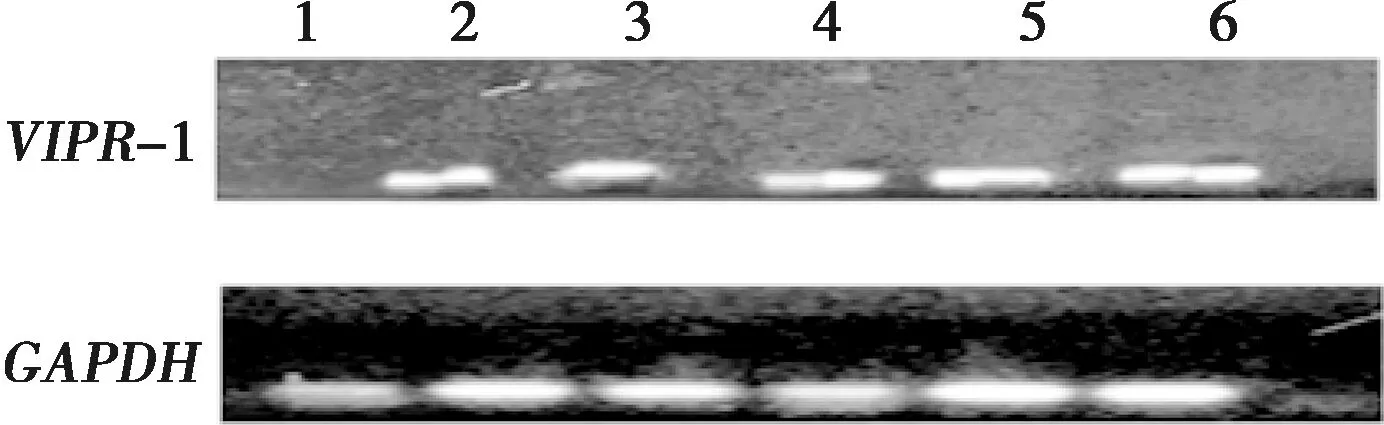
1.心脏;2.肝脏;3.脾脏;4.肺脏;5.肾脏;6.卵巢图4 VIPR-1基因在太行鸡各组织中表达
2.4 荧光定量PCR分析卵巢中VIPR-1基因表达水平
VIPR-1基因在高产母鸡和低产母鸡卵巢中相对表达量见图5。荧光定量PCR结果显示,VIPR-1基因在高产太行鸡卵巢组织中的表达量显著高于低产蛋鸡中的表达量(P<0.05)。
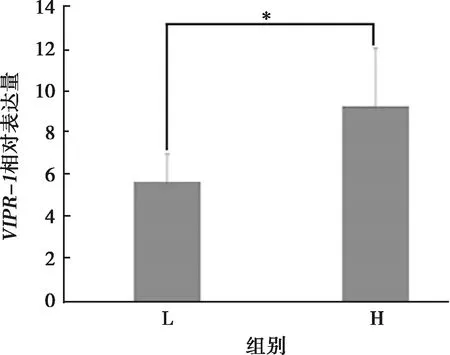
注:*表示差异显著(P<0.05);H.太行鸡高产组;L.太行鸡低产组图5 VIPR-1基因在太行鸡高产和低产母鸡卵巢中的表达量
3 讨论
太行鸡又称河北柴鸡,主要分布于河北省境内太行山区及周边地区,中心产区为沙河、赞皇、涞源等地,以产蛋为主,兼作肉用。太行鸡具有体型小、耐粗饲、抗逆性强、适应性强、觅食力强、蛋肉品质好,营养价值高等优点,但该地方鸡品种同时也存在着产蛋期产蛋量低的缺点[10]。本研究分析了对太行鸡血液中VIPR-1基因编码区的结构特征,进一步对不同物种的VIPR-1基因CDS序列及氨基酸序列进行同源性比较,利用定量和半定量PCR技术对VIPR-1基因在太行鸡中的表达模式进行研究,为深入探究VIPR-1基因的功能奠定了基础。
近年来,大量研究对禽类VIPR-1基因进行了研究,发现了VIPR-1基因在不同繁殖时期mRNA的表达量不同,证明了VIPR-1基因对产蛋性能有一定影响[11-12]。在洪军等[13]的研究中,以如皋黄鸡为素材,发现VIPR-1基因内含子2的2个多态位点与如皋黄鸡产蛋性能和蛋品质之间的关系,发现VIPR-1基因对如皋黄鸡的产蛋性能和鸡蛋品质有一定影响。目前,在家禽的研究中,鸡、黑番鸭、鹌鹑等物种的VIPR-1基因CDS序列已被克隆[2,4,6],然而在太行鸡中对于该基因的研究目前少有报道,本试验获得了太行鸡VIPR-1基因CDS全长,并对该基因的核酸序列和蛋白序列的分子特性进行了分析,发现太行鸡与野鸭VIPR-1基因一致性高达92.24%,其次是火鸡90.11%,表明该基因的功能在禽类中具有一定的保守性,推测其在功能上可能也具有一致性。
前人研究发现,VIPR-1基因在家禽驯化和饲养期间的改良中经受了较强的选择压力,其DNA序列突变与鸡的繁殖有关[14]。VIPR-1基因在家禽行为中起着重要的作用,VIPR-1基因的mRNA表达在不同的时期不尽相同[15]。Karacay等[16]研究发现,VIPR-1基因主要是通过与VIP蛋白特异性结合,调节PRL合成与分泌,进而影响畜禽的繁殖性能。朱志明等[17]研究发现VIPR-1基因的多个多态位点对蛋鸡产蛋性状存在显著影响,可以作为影响产蛋性状的重要分子标记。本试验发现VIPR-1基因在不同的组织均有表达,在卵巢组织中表达量最高;进一步发现VIPR-1基因在太行鸡高产组卵巢组织中的表达量显著高于低产组,该结论验证了前人结果,同时也表明该基因能够影响太行鸡的产蛋性能。
4 结论
太行鸡VIPR-1基因作为影响蛋鸡产蛋性能的重要候选基因,在其他家禽如火鸡、野鸭中具有较高的相似性;该基因在卵巢组织中高表达且在高产组中的表达量显著高于低产组,因此,该基因转录水平上的差异与太行鸡产蛋性能密切相关。本研究为进一步探究VIPR-1基因的分子作用机制和在太行鸡产蛋性能中的调控作用提供了依据,为后期深入研究蛋鸡产蛋性能的分子生物学机理奠定基础。
猜你喜欢
今日农业(2022年2期)2022-11-16 12:29:47
今日农业(2021年7期)2021-11-27 13:44:48
今日农业(2021年16期)2021-11-26 06:05:34
21世纪(2019年12期)2019-12-18 07:22:30
小哥白尼(野生动物)(2018年9期)2018-12-08 08:17:54
现代园艺(2017年19期)2018-01-19 02:49:58
湖南农业(2017年1期)2017-03-20 14:04:42
山东林业科技(2016年5期)2016-07-05 00:43:08
少儿科学周刊·少年版(2015年11期)2015-12-17 23:47:17
少儿科学周刊·少年版(2015年11期)2015-12-17 23:44:23
