
施氮量对花后高温胁迫后小麦同化物积累、转运及产量的影响
2020-11-09 05:20高春华冯波曹芳李升东王宗帅张宾王峥孔令安王法宏
中国农业科学 2020年21期
高春华,冯波,曹芳,李升东,王宗帅,张宾,王峥,孔令安,王法宏
施氮量对花后高温胁迫后小麦同化物积累、转运及产量的影响
高春华1,2,冯波1,曹芳1,李升东1,王宗帅1,张宾1,王峥1,孔令安1,王法宏1
(1山东省农业科学院作物研究所,济南 250100;2山东棉花研究中心,济南 250100)
【】明确不同施氮量对高温胁迫后小麦同化物积累和转运的影响及其生理基础,以期为小麦抗逆稳产栽培提供技术和理论依据。于2018—2019年在济南和济阳两地进行,以济麦44为材料,田间搭建高温棚进行高温胁迫处理,设置2个温度处理(CK:未胁迫,H:花后高温胁迫),3个氮肥水平(低氮N1:180 kg·hm-2,常规氮N2:240 kg·hm-2,高氮N3:300 kg·hm-2)。通过分析小麦花前同化物质的转运、成熟期同化物质的积累与分配、叶片与籽粒中蔗糖合成酶在同化物转运中的作用,阐明了不同施氮量对花后高温胁迫后小麦籽粒产量形成的影响机制。不同施氮量对高温胁迫后小麦的减产率影响不同,N1处理减产率为54.78%(济南)和50.19%(济阳),N2处理为24.05%(济南)和25.29%(济阳),N3处理为54.49%(济南)和44.13%(济阳)。高温胁迫后,与N1和N3处理相比,N2处理成熟期同化物积累量、花前营养器官同化物向籽粒中转运量和转运率、花后同化物积累量和积累率、同化物向籽粒中的分配比例均显著增加;N2处理旗叶SPAD值、蔗糖合成酶SS-Ⅱ合成方向活性和籽粒蔗糖合成酶SS-Ⅰ分解方向活性增加。本试验条件下,施氮量为240 kg·hm-2可以显著减缓高温胁迫后旗叶衰老,维持旗叶中SS-Ⅱ和籽粒中SS-Ⅰ较高的酶活性,保持较高的同化物合成能力和向籽粒中的转运能力,提高同化物向籽粒中的积累量和分配比例,降低高温胁迫后小麦籽粒产量的损失率。
小麦;施氮量;高温胁迫;同化物质积累;同化物质转运;籽粒产量
0 引言
【研究意义】近年来全球平均温度升高,地域性高温经常发生[1-4],高温胁迫已经成为小麦产量形成过程的主要非生物限制因素之一。山东省位于黄淮海冬小麦主产区的东部,小麦在灌浆期经常遭遇3—5 d的高温天气,超过30℃的短时高温会引起旗叶光合速率降低、灌浆时间缩短,千粒重降低,籽粒中淀粉合成受阻,严重的地区和年份减产幅度可达30%以上[5-7]。开展不同施氮量对花后高温胁迫下小麦籽粒产量的影响及其形成生理基础的研究,为小麦抗逆稳产栽培提供技术支持,对保证粮食安全具有重大意义。【前人研究进展】小麦属喜凉作物,小麦籽粒灌浆的适宜温度是20℃—30℃[8],当温度超过35℃时,小麦籽粒产量显著降低[9]。高温胁迫后,叶片叶素绿含量降低,加速叶片衰老,缩短了叶片中光合产物向籽粒中的运输时间[10]。籽粒灌浆终止早于叶片完全衰老,导致叶片光合产物被分配到其他营养器官中[11],从而降低了叶片中同化物质向籽粒中的转运[12],抑制了籽粒中光合产物的累积,最终影响粒重和产量。氮的供应改变热胁迫对小麦籽粒重量影响程度[13-14],小麦在高施氮量下灌浆期高温胁迫效应更为显著,籽粒产量减产幅度增大[15-16]。低施氮量的粒重在高温胁迫后降低幅度小于高施氮量[17],高土壤有效氮供给下,高温胁迫对籽粒产量和产量构成因素损害更大[16]。高温胁迫下,灌浆期缩短,淀粉积累进程提前结束[7]。蔗糖合成酶是蔗糖向淀粉转化过程中的第1个限速酶,其活性高低和活性持续期长短决定着籽粒灌浆速率的大小和持续时间的长短,并与籽粒的体积和重量密切相关[18]。高温胁迫显著降低了小麦籽粒蔗糖合成酶活性[19-20],而施氮量不足或过量降低玉米穗位叶蔗糖合成酶活性[21-22]。前人通过增施穗肥、叶面施镁、选择品种和调整播期等措施减小高温胁迫伤害[23-26],为降低高温胁迫危害提供了理论依据。然而,由于小麦生育后期高温胁迫的突发性,胁迫发生程度、发生时间、持续时间等的不确定性,生产上调控高温胁迫的栽培技术大都具有一定的局限性。【本研究切入点】考虑到高温胁迫的频发性以及氮肥管理的普遍性,在前人研究基础上,研究不同施氮量对小麦花后高温胁迫的影响。前人关于高温胁迫研究多在温室、生长箱和盆栽条件下进行,在大田条件下,不同施氮量对花后高温胁迫后小麦产量影响的报道较少。【拟解决的关键问题】本研究以济麦44为材料,在大田条件下,通过搭建增温棚进行高温胁迫处理,探讨不同施氮量下小麦同化物积累分配转运及相关酶活性对花后高温胁迫的响应,明确高温胁迫对冬小麦产量影响以及不同施氮量对花后高温胁迫后小麦籽粒产量形成影响的生理基础,为冬小麦抗逆和调优栽培提供技术和理论支撑。
1 材料与方法
1.1 试验地概况
试验于2018—2019年小麦生长季,在山东省农业科学院济南试验基地(117.07°E,36.68°N)和山东省农业科学院济阳试验基地(117.22°E,36.98 °N)两地进行。济南试验地为褐土,有机质含量15.1 g·kg-1,全氮1.29 g·kg-1,速效氮60.9 mg·kg-1,速效磷34.6 mg·kg-1,速效钾146.7 mg·kg-1;济阳试验地为潮土,有机质含量10.3 g·kg-1,全氮1.20 g·kg-1,速效氮59.9 mg·kg-1,速效磷32.7 mg·kg-1,速效钾129.8 mg·kg-1。
1.2 试验设计
田间搭建高温棚,于花后10 d至花后17 d进行高温胁迫(H),搭建高温棚期间,无有效降雨。高温棚采用0.07 mm无色透明的聚乙烯塑料膜做成,透光率为92.3%,棚高1.5 m,棚底距地面30 cm,保证棚内通风条件良好,底面积与小区面积相同。每日8:00—18:00进行高温处理,在试验小区冠层位置悬挂温湿度计,每30 min自动记录温湿度,以此计算两地试验各处理增温阶段日均温度(图1)。
试验材料选用山东省地区推广的小麦品种济麦44(JM44),采用二因素随机区组设计,设2个温度处理(CK:未胁迫,H:高温胁迫),3个氮肥水平(低氮N1:180 kg·hm-2,常规N2:240 kg·hm-2,高氮N3:300 kg·hm-2),小区面积12 m2,重复3次。所用肥料为尿素(N 46.4%),磷酸二铵(P2O546%,N 18%),氯化钾(K2O 52%),50%氮肥作为基施,播种时同时施入,50%于拔节期追施。磷钾肥用量为P2O5112.5 kg·hm-2、K2O 112.5 kg·hm-2,全部基施。2018年10月15日播种,基本苗为180株/m2,其他田间管理同正常大田。
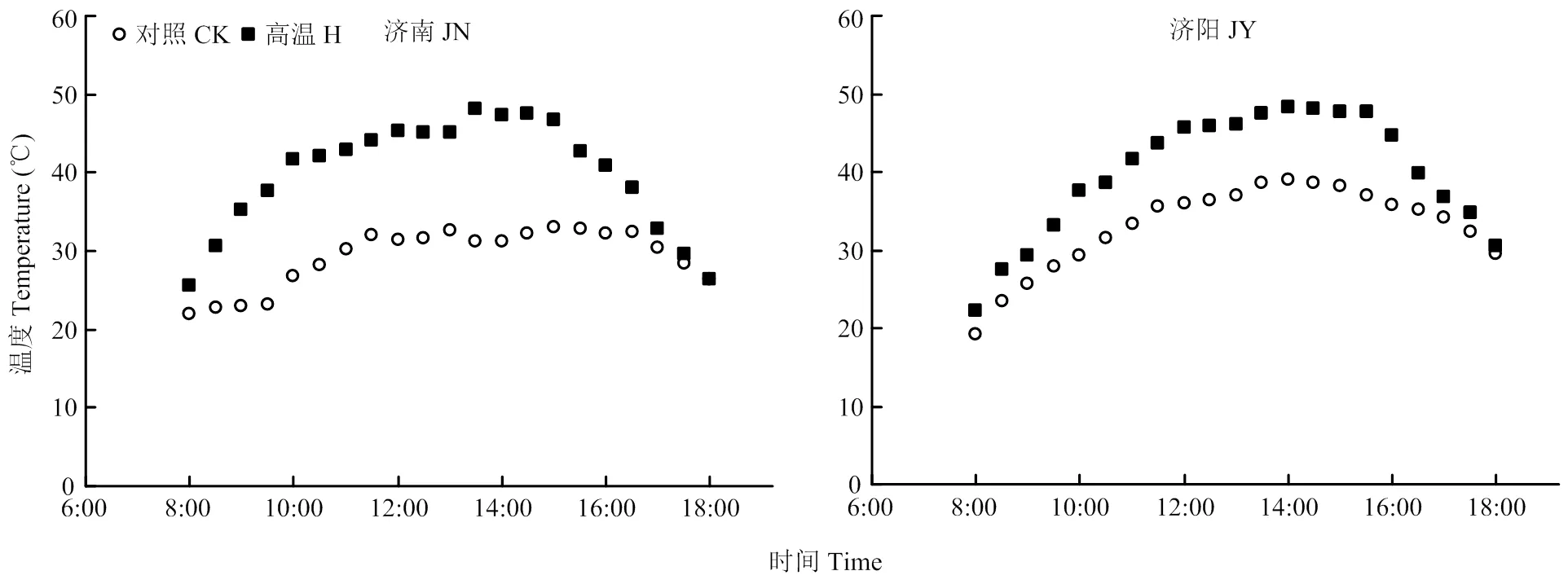
图1 对照和花后高温胁迫处理下的小麦冠层温度
1.3 测定项目与方法
1.3.1 取样 济南试验地开花期为5月2日,成熟期为6月4日;济阳试验地开花期为5月7日,成熟期为6月12日。开花期对长势一致的单茎挂牌标记,开花期和成熟期取标记的单茎,分为穗、旗叶、其余叶、茎+叶鞘,105℃杀青30 min,80℃烘至恒重,测干物质重。于高温胁迫第3天,取标记植株的穗和旗叶,液氮速冻,置-80℃冰箱保存,用于测定蔗糖合成酶活性。成熟后每小区收获2 m2计产。
1.3.2 干物质积累与分配的测定 开花期和成熟期数分蘖成穗数,成熟期按叶、茎+叶鞘、穗轴+颖壳和籽粒4部分分样,称重后用于计算各器官分配比例。同化物质的转运和积累计算公式如下[27]:
营养器官开花前贮藏干物质转运量=开花期营养器官干重-成熟期营养器官干重;
营养器官开花前贮藏干物质转运率(%)=(开花期营养器官干重-成熟期营养器官干重)/开花期营养器官干重×100;
开花后干物质输入籽粒量=成熟期籽粒干重-营养器官花前贮藏干物质转运量;
营养器官花前贮藏干物质对籽粒产量的贡献率(%)=开花前营养器官贮藏干物质转运量/成熟期籽粒干重×100;
开花至成熟期的干物质积累速率=(成熟期的干物质积累量-开花期的干物质积累量)/开花至成熟的天数。
1.3.3 旗叶SPAD值测定 于胁迫后第3天,采用SPAD-502PLUS便携式叶绿素仪(日本托普),选取10个标记的单茎旗叶和顶二叶进行测量,取平均值。
1.3.4 酶活性测定方法 蔗糖合成酶合成方向活性测定参考Rufty等[28]的方法,并加以改进。200 µL酶液加50 µL pH7.5的HEPES-NaOH缓冲液、50 µL 10 mmol·L-1MgCl2、100 µL 50 mmol·L-1UDPG、100 µL 50 mmol·L-16-磷酸果糖,30℃反应30 min后,100℃水浴杀死酶活性,加200 µL 40% NaOH溶液混匀,100℃保温10 min,加 2.0 mL30%盐酸混匀,80℃保温10 min,再加1 mL 1%间苯二酚,混匀后 80℃保温10 min,冷却后于480 nm下比色测生成磷酸蔗糖含量。用蔗糖生成量表示酶活性。
蔗糖合成酶分解方向活性测定参考Sebkova等[29]的方法,取4个1.5 ml的离心管,分别标记对照管、测定管、标准管和空白管。对照管:10 µl酶提取液加40 µl pH7.5的HEPES-NaOH缓冲液;测定管:10 µl酶提取液加40 µl HEPES-NaOH缓冲液(含蔗糖和MgAc2·4H2O),将对照管和测定管30℃水浴30 min后,95℃水浴10 min(盖紧,防止水分散失)。标准管:10 µl的20 mg·ml-1果糖标准液加40 µl pH 7.5的HEPES-NaOH缓冲液;空白管:10 µl的蒸馏水加40 µl pH7.5的HEPES-NaOH缓冲液。在4个离心管中加50 µl DNS试剂终止反应,混匀,沸水浴5 min,冷却后加400 µl蒸馏水,混匀后测定540 nm处的吸光值。对照反应体系中不含UDPG。
1.4 数据分析与方法
用Excel 2010进行数据处理,籽粒产量、同化物的积累、转运、分配及相关酶活性等指标均进行3次重复检测,所有指标进行二因素随机区组的方差分析(DPS18.5数据处理系统),并采用多重比较法中的最小显著性差异法(LSD)进行高温、氮肥及两者交互作用的方差检测(<0.05),用Sigmaplot10.0软件作图。
2 结果
2.1 不同处理籽粒产量及减产率
与对照相比,高温胁迫显著降低2个试验点小麦籽粒产量,但不同施氮量处理减产率不同(图2),N1、N2、N3处理的减产率分别为54.78%、24.05%、54.49%(济南)和50.19%、25.29%、44.13%(济阳)。2个试验地均是N2处理减产率最低,显著低于N1和N3处理。表明N2处理可以缓解花后高温胁迫对小麦籽粒产量的抑制。
2.2 不同处理产量构成因素及同化物积累
高温、施氮量及两者间的交互作用对各性状均有极显著的影响,与对照相比,高温胁迫使单穗重、穗粒数、千粒重、成熟期同化物积累量、积累速率和籽粒收获指数均显著降低,但不同氮处理降低的幅度不同(表1)。
高温胁迫后,千粒重和花后同化物积累速率下降,差异极显著,N1和N3处理千粒重2个试验地降幅为44.28%—60.94%,N2处理为17.53%(济南)和14.59%(济阳),N1和N3处理花后同化物积累速率降幅为32.45%—56.18%,N2处理为24.27%(济南)和29.96%(济阳),说明粒重降低和花后同化物积累速率减缓是花后高温胁迫导致产量降低的主要原因,N2处理在花后高温胁迫后能保持较高的粒重和花后同化物积累速率,从而获得相对较高的籽粒产量。
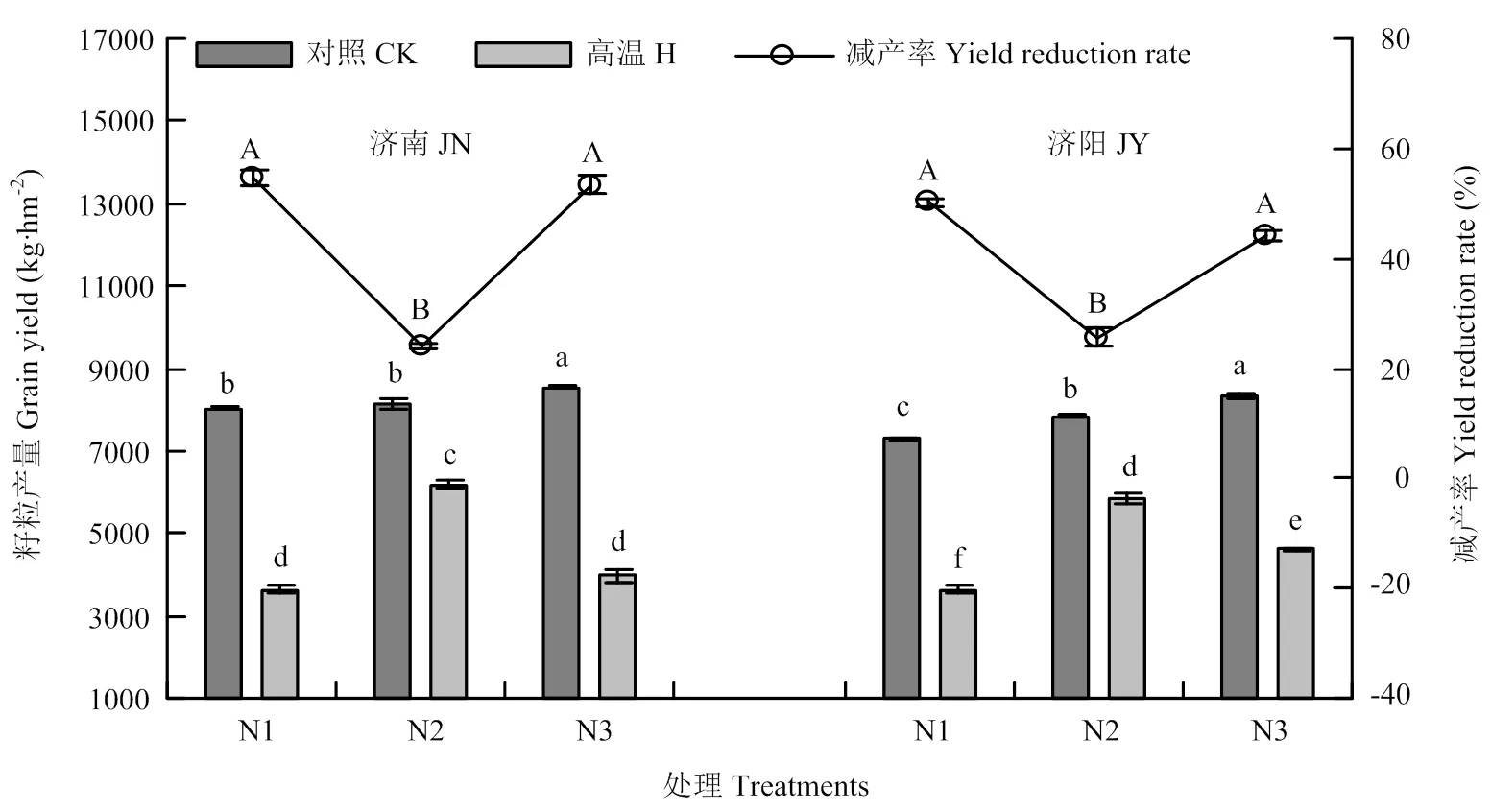
不同小写字母代表示产量处理间差异达5%显著水平,不同的大写字母表示减产率处理间差异达5%显著水平
2.3 成熟期同化物在各器官的分配量及比例
高温胁迫后穗轴+颖壳、叶片和茎鞘的同化物积累量显著升高,粒重显著降低,最终单茎重显著降低(图3)。不同施氮量处理中,N2处理籽粒重最高。
高温胁迫使成熟期同化物在穗轴+颖壳、叶片、茎鞘中的百分比较对照显著增加,籽粒中的百分比显著降低(图4)。高温胁迫使N2处理籽粒中百分比下降30.30%(济南)和24.60%(济阳),显著小于N1(济南67.92%,济阳66.91%)和N3处理(济南52.33%,济阳62.93%)。高温胁迫后N1和N3处理同化物在穗轴+颖壳中分配比例的增幅(31%—42.38%)显著高于叶片(12.38%—27.72%)和茎鞘(12.42%—28.44%)。
2.4 开花后营养器官中同化物向籽粒中的转运
花前营养器官同化物转运量、转运率和对籽粒的贡献率、花后同化物积累量和对籽粒的贡献率在高温、施氮量及两者间的交互作用下均存在显著性差异(表2)。高温胁迫显著降低了不同氮处理花前同化物的转运量、转运率、对籽粒的贡献率以及花后同化物积累量,而花后同化物积累量对籽粒的贡献率升高。高温胁迫后N2处理花前同化物的转运量、转运率以及花后同化物积累量均显著高于N1和N3处理,降幅小于N1和N3处理。高温胁迫下,济南试验地的花前同化物对籽粒的贡献率N1和N2处理间无显著差异,但显著高于N3处理;济阳试验地的N2和N3处理间无显著差异,但是显著高于N1处理;济南试验地的花后同化物对籽粒的贡献率N1和N2处理间无显著差异,但显著低于N3处理,济阳试验地的N2和N3处理间无显著差异,但是显著低于N1处理。说明N2处理在高温胁迫下保持较高的同化物向籽粒中转运的能力。
2.5 不同叶位叶片的SPAD值
与对照相比,高温胁迫显著降低了旗叶和顶二叶的SPAD值,且不同施氮量处理间存在显著差异(图5)。高温胁迫后N1、N2和N3处理旗叶SPAD值的降幅分别为10.24%、1.99%、7.40%(济南)和13.91%、4.36%、9.87%(济阳)。N2处理旗叶SPAD值降幅最低,在高温胁迫后仍具有较高的叶绿素含量。高温胁迫N1、N2和N3处理后顶二叶SPAD值的降幅分别为9.62%、9.29%、7.74%(济南)和10.14%、8.69%、7.60%(济阳),降幅在不同氮肥水平间差异不显著。
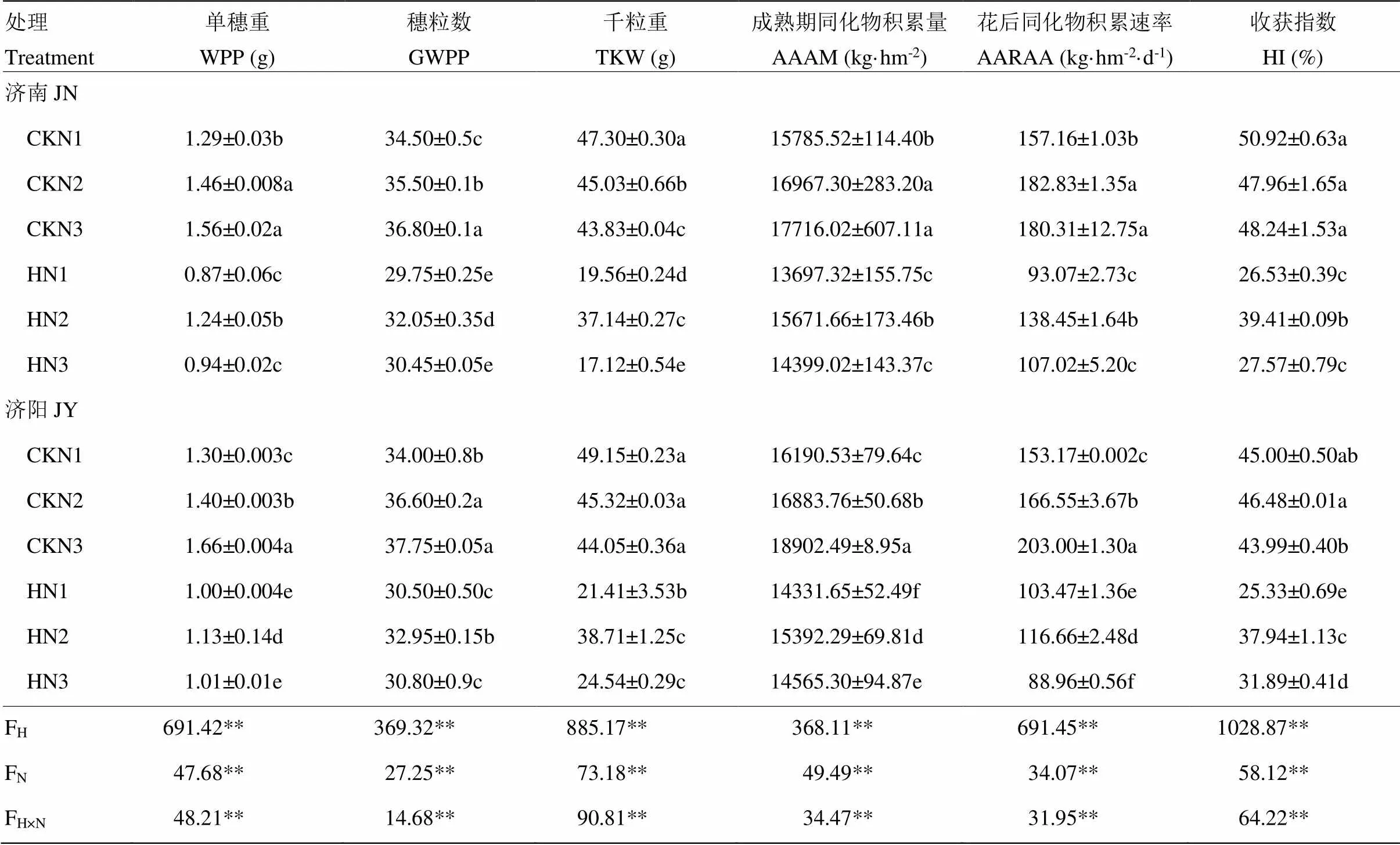
表1 高温胁迫和施氮量对小麦产量构成因素及同化物积累的影响
WPP:单穗重,GWPP:单穗粒数,TKW:千粒重,AAAM:成熟期同化物,AARAA:花后同化物积累速率,HI:收获指数,FH:高温处理的F值,FN:氮肥处理的F值,FH×N:高温和氮肥相互作用的F值。同列中不同小写字母表示处理间差异达5%显著水平,“±”后面的数字代表的是标准误,**表示处理间在0.01水平下差异显著,*表示处理间在0.05水平下差异显著。下同
WPP: Weight per panicle, GWPP: Grain weight per panicle, TKW: Thousand kernel weight, AAAM: Assimilate accumulation at maturity, AARAA: Assimilate accumulation rate after anthesis, HI: Harvest index. FH: F value of high temperature, FN: F value of nitrogen fertilizer, FH×N: F value of high temperature and nitrogen fertilizer. Different small letters indicate significant difference among treatments at 0.05 level, the number after “±” is the standard error, **indicates significant difference among treatments at 0.01 level, * indicates significant difference among treatments at 0.05 level. The same as below
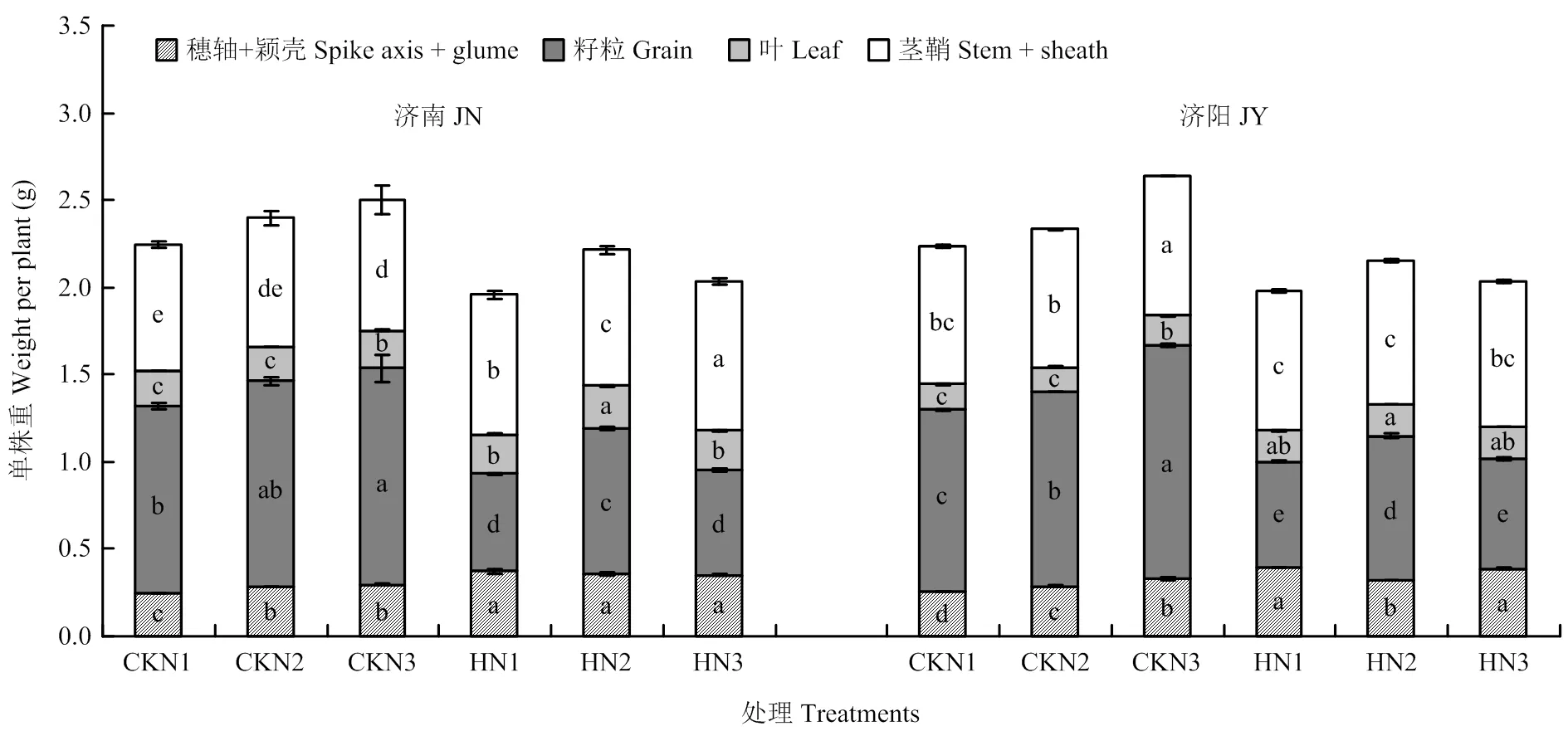
不同小写字母表示相同器官处理间差异达5%显著水平。下同
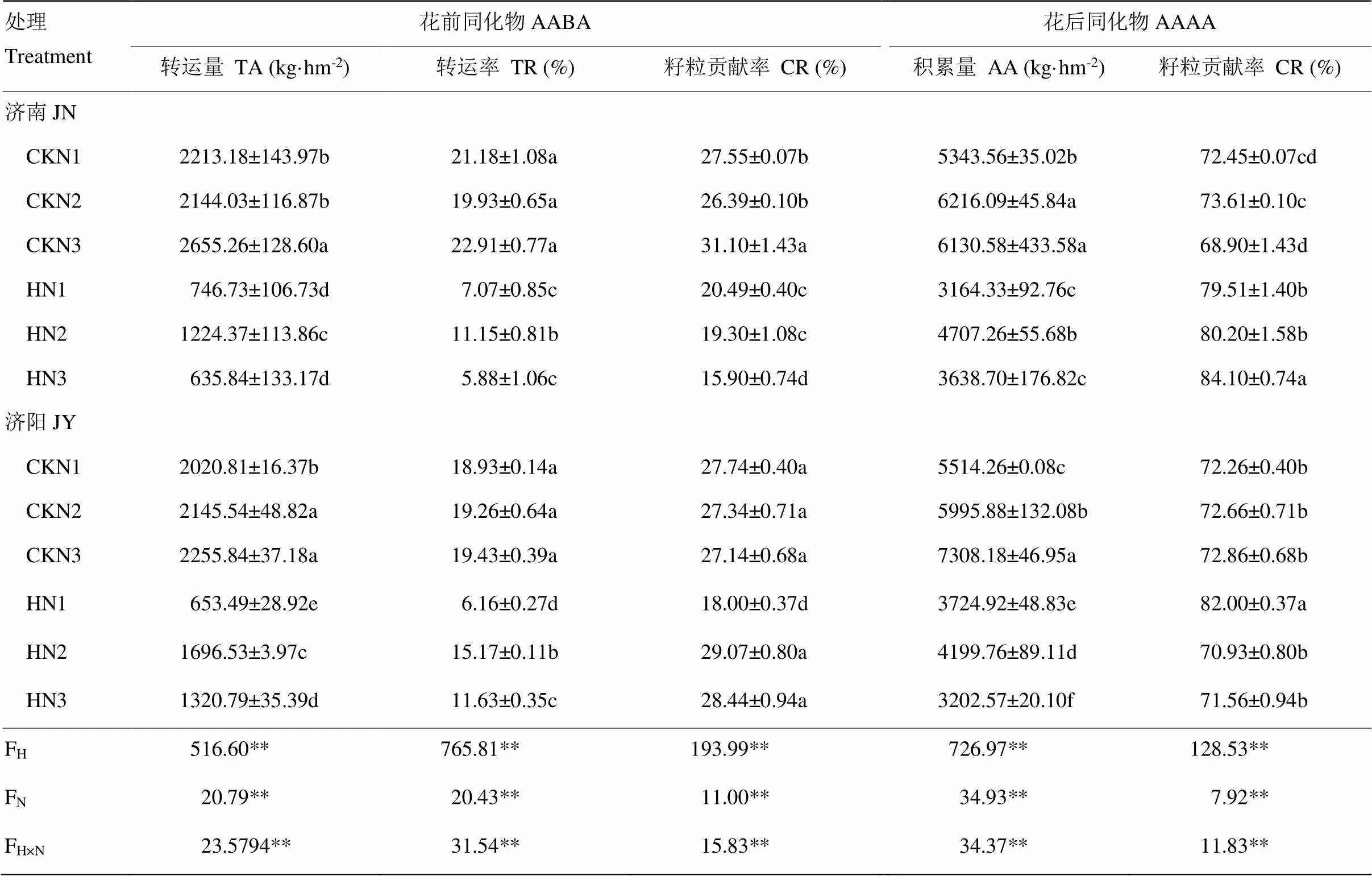
表2 花前同化物向籽粒的转运量及花后同化物积累量对高温胁迫和施氮量的响应
AABA:花前同化物,AAAA:花后同化物,TA:转运量,TR:转运率,CR:对籽粒的贡献率,AA:积累量
AABA: Assimilate accumulation before anthesis, AAAA: Assimilate accumulation after anthesis, TA: Translocation accumulation, TR: Translocation rate, CR: Contribution rate, AA: Assimilate accumulation
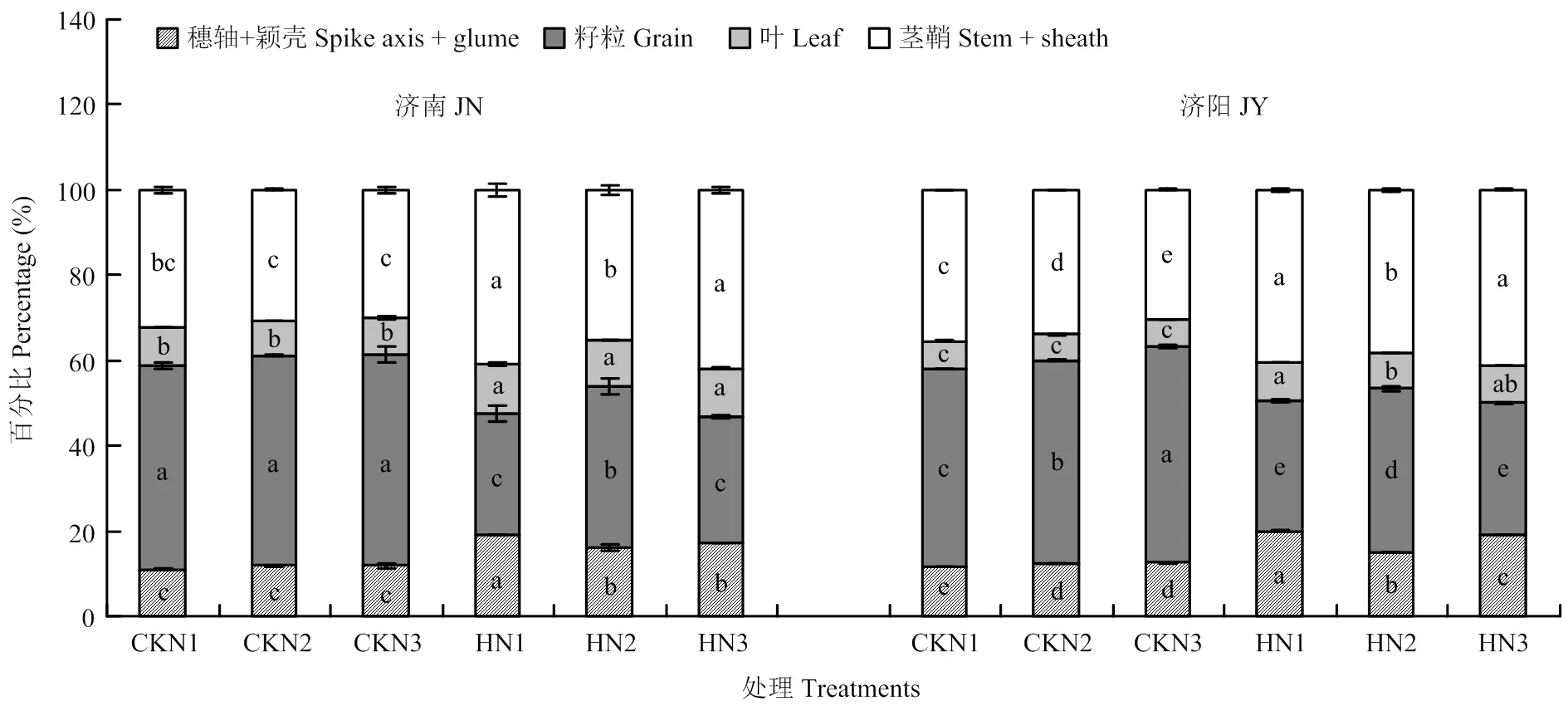
图4 成熟期同化物在不同器官中的分配比例
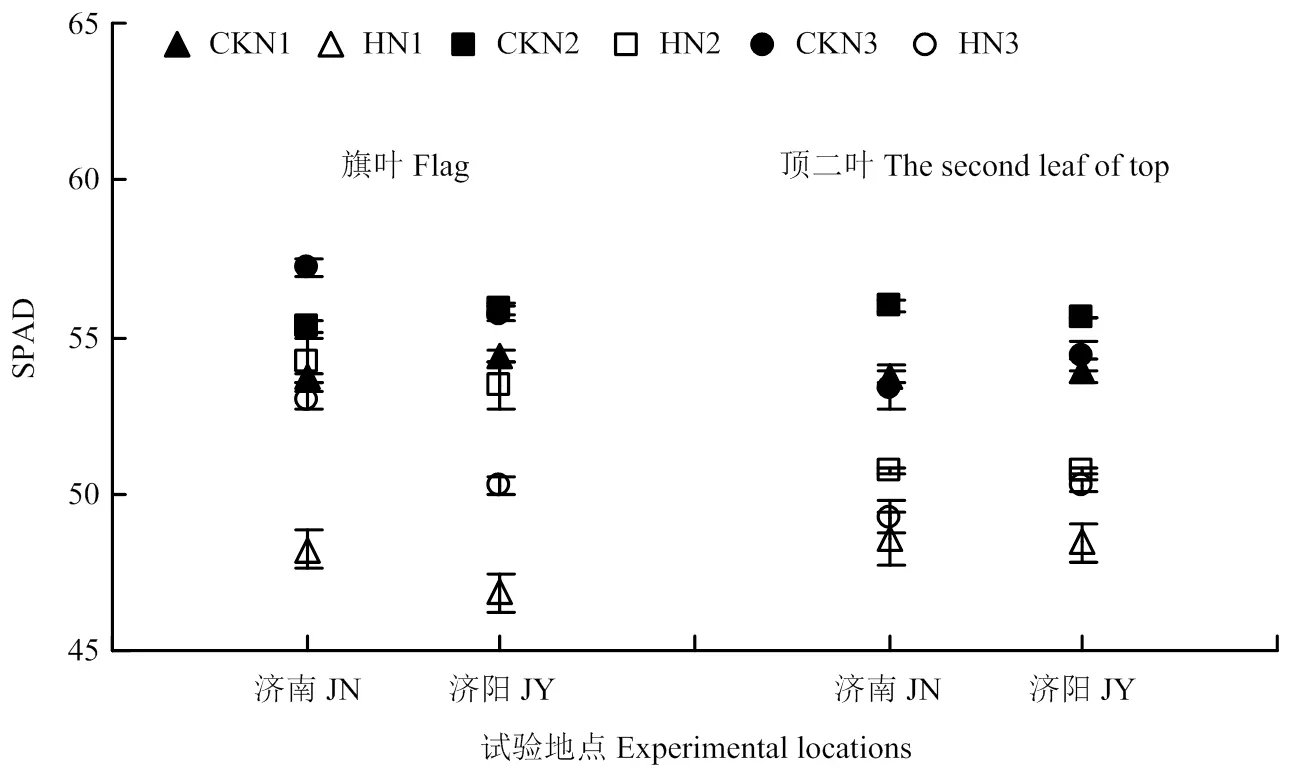
图5 高温胁迫和施氮量对不同叶位叶片SPAD值的影响
2.6 籽粒和旗叶中蔗糖合成酶活性
由图6可知,与对照相比,高温胁迫后籽粒中蔗糖合成酶分解方向(SS-Ⅰ)酶活显著降低,N1处理降幅为16.36%,N2降幅为13.43%,N3降幅为22.69%。籽粒中蔗糖合成酶合成方向(SS-Ⅱ)的酶活显著升高,N1处理升幅为34.85%,N2处理升幅为21.31%,N3处理升幅15.94%。说明籽粒中合成淀粉的底物减少,淀粉的合成量减少,最终影响粒重。
高温胁迫后旗叶在下蔗糖合成酶分解方向活性升高,其中N1和N3处理较对照差异显著,N2处理与对照无显著差异(图7)。各施氮量处理旗叶蔗糖合成酶合成方向活性显著降低,N3处理显著低于N1和N2处理。说明高温胁迫导致旗叶蔗糖分解能力升高,合成蔗糖能力降低,输出物质减少。
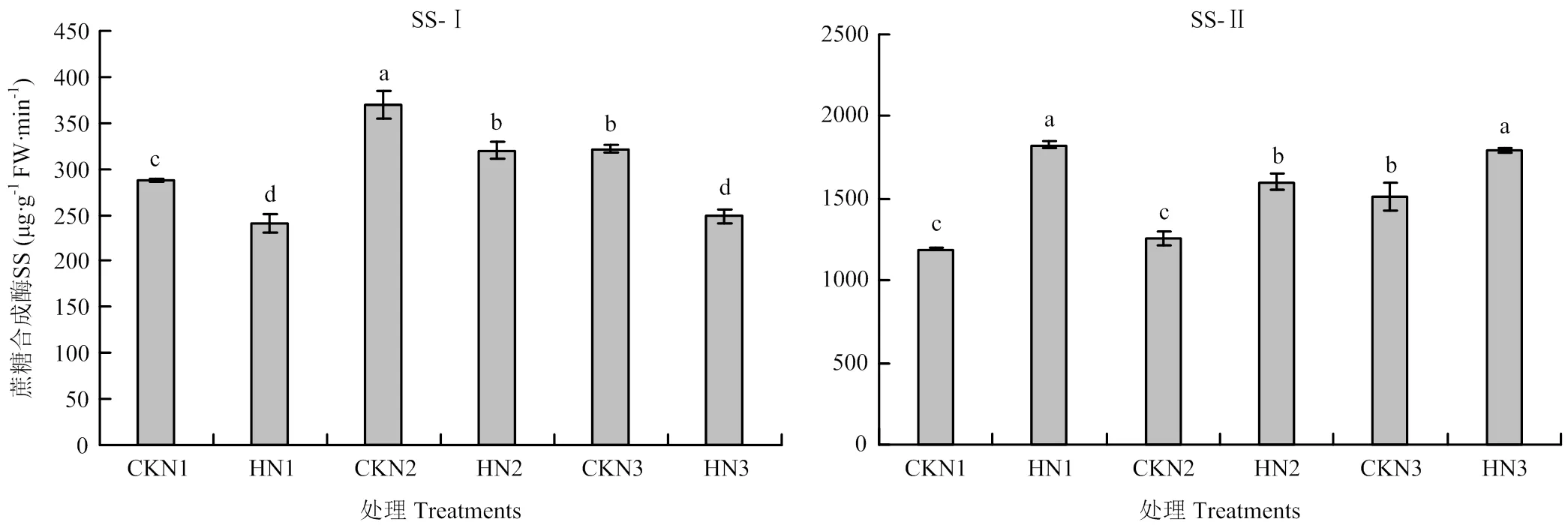
SS-Ⅰ:蔗糖合成酶分解方向,SS-Ⅱ:蔗糖合成酶合成方向。不同小写字母表示处理间差异达5%的显著水平。下同
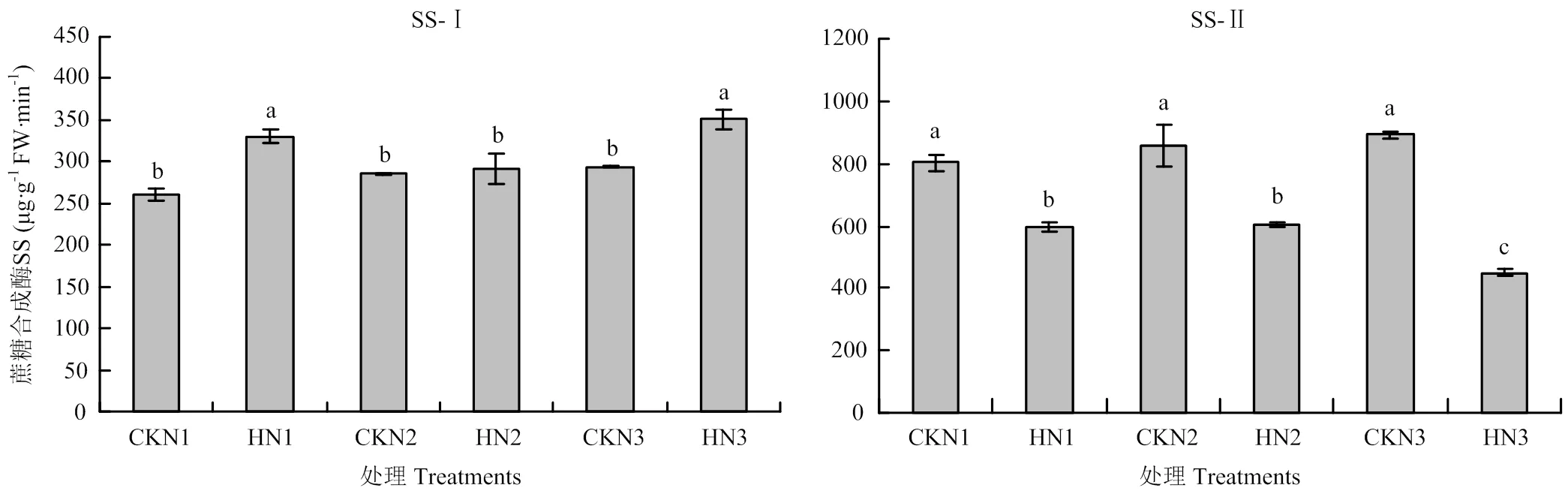
图7 不同处理旗叶中蔗糖合成酶活性(济南)
3 讨论
3.1 施氮量对高温胁迫后小麦籽粒产量及产量构成因素的影响
Liu等[30]通过冬季增温和氮肥试验表明,施氮处理在增温条件下籽粒产量显著低于未施氮处理。对玉米和春小麦的高温肥料试验表明,随着氮肥供给的增加,籽粒减产率增加[30-31],MÓNICA等[17]通过在不同地力条件下进行高温胁迫试验发现,土壤供氮量越高籽粒减产率越大。本试验在基础地力相当的济南、济阳两地进行,通过不同的施氮量改变土壤供氮量,籽粒产量和减产率均表现显著差异,施氮量从180 kg·hm-2(N1)增加到240 kg·hm-2(N2),花后高温胁迫对籽粒产量影响降低,施氮量增加到300 kg·hm-2(N3),高温胁迫对籽粒产量的影响显著增加,在对照条件下,N3的籽粒产量高,高温胁迫后籽粒产量减产的绝对值也大。说明低施氮量和过高施氮量均加剧高温胁迫对产量的抑制作用,合理施氮量可以相对减小花后高温胁迫的危害,获得较高籽粒产量。在本试验条件下,240 kg·hm-2为合理的施氮量。
灌浆期高温胁迫使穗粒数和粒重均显著降低,但降低量随着氮供应量升高而升高[17]。也有盆栽试验发现,花后高温胁迫后,适量氮肥增加穗粒数及籽粒千粒重,提高籽粒产量[31]。本试验高温胁迫后,各处理单穗重、穗粒数、千粒重、成熟期同化物积累量、花后同化物积累速率和收获指数均显著降低。其中,N2处理单穗重、穗粒数、千粒重、成熟期同化物积累量、花后同化物积累速率和收获指数显著高于N1和N3处理,说明N2处理能降低同化物积累和花后同化物积累速率降幅,降低高温胁迫对单穗重、穗粒数、千粒重的损害程度,减轻产量损失。
3.2 施氮量对高温胁迫后小麦同化物积累转运的影响
小麦籽粒产量的影响因素是同化物的合成能力和同化物向籽粒中转运能力,转运能力与库对同化物的竞争能力有关。籽粒产量的1/3来自花前贮藏同化物的转运,2/3来自花后叶片光合同化物的积累[32]。研究表明,高温处理使同化物向库器官的分配受到阻碍,输出时间缩短,输出速率下降,产量降低[11-12]。适量氮肥处理能够促进高温胁迫下春小麦花前营养器官贮藏同化物向籽粒中转运,增加花后积累同化物对籽粒贡献率[31]。本研究结果表明,高温胁迫降低了花前贮藏同化物向籽粒中转移量和花后同化物的积累量,从而降低了成熟期同化物积累总量。成熟期籽粒中同化物的分配量及比例减少,而穗轴+颖壳和茎鞘的分配量和比例显著增加,这与前人的研究结果一致[10-12]。两地试验中,N2处理的花前营养器官同化物转运量和花后同化物的积累量均显著高于N1和N3处理,N2处理籽粒中同化物积累量和分配比例降幅低于N1和N3处理,在穗轴+颖壳和茎鞘中的分配量及比例的升幅显著高于N1和N3处理。济南试验地,N3处理的花后同化物的转运量低于N1和N2处理,花后同化物的积累量低于N2处理。济阳试验地,N3处理的花后同化物的转运量低于N2处理,高于N1处理,花后同化物的积累量低于N1和N2处理,表明在N3处理条件下不利于同化物的积累和转运。以上结果说明,N2处理较N1和N3处理能优化花后高温胁迫后同化物向籽粒中转运和分配,具有较高的同化物转运能力。
3.3 施氮量对高温胁迫后叶片衰老和籽粒中酶活的影响
小麦开花后,旗叶作为功能叶其活力持续时间对于产量形成具有重要作用。Mónica等[17]研究表明,高温对衰老模式的影响微乎其微,叶片SPAD读数无显著差异。而Talukder等[33]和Wang等[34]研究认为,高温胁迫加速了旗叶叶绿素含量的损失,促进旗叶衰老。本试验结果表明,高温胁迫显著降低了旗叶和顶二叶的SPAD值,N2处理较N1和N3处理能有效减缓旗叶叶绿素下降,延缓旗叶衰老。高温胁迫后,N2处理顶二叶SPAD值与N1、N3处理间差异不显著,这可能与顶二叶中衰老酶活性有关,尚需进一步研究。
蔗糖是光合作用过程中最主要的产物,蔗糖合成酶是催化蔗糖进入其他新陈代谢途径的关键酶,具有双向催化作用。小麦籽粒中蔗糖合成酶SS-Ⅰ分解方向活性高低反映了籽粒降解蔗糖的能力,较高的活性可提供充足的淀粉合成底物。温度和施氮量影响蔗糖合成酶活性。小麦花前夜间增温下,蔗糖合成酶活性随施氮量的增加而增加[21]。刘霞等[35]研究认为,高温降低了籽粒中蔗糖合成酶活性。陈洋等[22]和赵宏伟等[23]发现施氮量不足或过量均导致玉米穗位叶蔗糖合成酶活性下降。本试验在济南和济阳结果表现一致,以济南试验点为代表检测酶活性,发现灌浆期高温胁迫后籽粒中蔗糖合成酶分解方向活性降低,合成方向活性升高;旗叶中蔗糖合成酶分解方向活性升高,合成方向活性降低。N2处理籽粒中蔗糖合成酶分解方向活性和旗叶中蔗糖合成酶合成方向活性高于N1和N3处理;高温胁迫后,N3处理籽粒中蔗糖合成酶分解方向活性最低,而旗叶中蔗糖合成酶合成方向的活性最高,表明N3处理籽粒中合成淀粉的底物少,降低了籽粒的粒重。说明N2处理籽粒中淀粉合成底物较多,旗叶具有较强的蔗糖合成能力,这是N2处理获得较高干物质积累量和粒重的基础。
4 结论
高温胁迫降低小麦成熟期干物质的积累量、向籽粒中的转运量及转运率,从而降低产量。N2(240 kg·hm-2)处理较N1(180kg·hm-2)和N3(300kg·hm-2)处理能够延缓花后高温胁迫后的旗叶衰老,使旗叶保持较高的蔗糖合成能力,籽粒保持较高的蔗糖分解能力,同化物积累和同化物向籽粒转运能力受损较低,获得较高籽粒产量。
[1] Barlow K M, Christy B P, O’Leary G J, Riffkin P A, Nuttall J G. Simulating the impact of extreme heat and frost events on wheat crop production: A review., 2015, 171: 109-119.
[2] Asseng S, Ewert F, Martre P, Rötter R P, Lobell D B, Cammarano D, White J W. Rising temperatures reduce global wheat production., 2014, 5(2): 143-147.
[3] Rahmstorf S, Coumou D. Increase of extreme events in a warming world.2011, 108(44) : 17905-17909.
[4] Xiao Y G, Qian Z G, Wu K, Liu J J, Xia X C, Ji W Q, He Z H. Genetic gains in grain yield and physiological traits of winter wheat in Shandong province, China, from 1969 to 2006., 2012, 52(1) : 44-56.
[5] FAROOQ M, Bramley H, Palta J A. Heat stress in wheat during reproductive and grain-filling phases., 2011, 30(6) : 479-492.
[6] 冯波, 李升东, 李华伟, 王宗帅, 张宾, 王法宏, 孔令安. 灌浆初期高温胁迫对不同耐热性小麦品种形态和产量的影响. 中国生态农业学报, 2019, 27(3): 451-461.
Feng B, Li S D, Li H W, Wang Z S, Zhang B, Wang F H, Kong L A. Effect of high temperature stress on plant morphology and grain yield of different heat resistance varieties in wheat during early grain-filling stage., 2019, 27(3): 451-461. (in Chinese)
[7] 王晨阳, 张艳菲, 卢红芳, 赵君霞, 马耕, 马冬云, 朱云集, 郭天财, 马英, 姜玉梅. 花后渍水、高温及其复合胁迫对小麦籽粒淀粉组成与糊化特性的影响. 中国农业科学, 2015, 48(4): 813-820.
Wang C Y, Zhang Y F, Lu H F, Zhao J X, Ma G, Ma D Y, Zhu Y J, Guo T C, Ma Y, Jiang Y M. Effect of post-anthesis waterlogging, high temperature and their combination on strarch compositions and pasting properties in wheat grains., 2015, 48(4): 813-820. (in Chinese)
[8] 邓志英, 田纪春, 胡瑞波, 赵亮, 盛峰, 王延训, 张永祥, 孙国兴, 孙彩玲. 适度高温对不同筋力冬小麦蛋白组分、面粉品质和面条加工品质的影响. 中国粮油学报, 2006, 21(4): 25-31.
Deng Z Y, Tian J C, Hu R B, Zhao L, Sheng F, Wang Y X, Zhang Y X, Sun G X, Sun C L. Effects of moderate high temperature on protein components, flour quality and noodle making quality from winter wheat cultivars with different gluten strength., 2006, 21(4): 25-31. (in Chinese)
[9] Niwas R, Khichar M L. Managing impact of climatic vagaries on the productivity of wheat and mustard in India., 2016, 67(1): 205-222.
[10] 郑飞, 何钟佩. 高温胁迫对冬小麦灌浆期物质运输与分配的影响. 中国农业大学学报, 1999, 4(1): 73-76.
Zheng F, He Z P. Effect of high temperature stress on transportation and distribution of14C-assimilaes in grain filling period of winter wheat., 1999, 4(1): 73-76. (in Chinese)
[11] Kim J, Shon J, Lee C K, Yang W, Yoon Y, Yang W H, Lee B W. Relationship between grain filling duration and leaf senescence of temperate rice under high temperature., 2011, 122(3): 207-213.
[12] 郭文善, 施劲松, 彭永欣, 封超年, 葛才林, 朱新开. 灌浆期高温对小麦光合产物运转的影响. 核农学报, 1998, 12(1): 21-27.
Guo W S, Shi J S, Peng Y X, Feng C N, Ge C L, Zhu X K. Effect of high temperature on transportation of assimilate from wheat flag leaf during grain filling stage., 1998, 12(1): 21-27. (in Chinese)
[13] Dupont F M, Hurkman W J, Vensel W H, Tanaka C, Kothari K M, Chung O K, Altenbach S B. Protein accumulation and composition in wheat grains: Effects of mineral nutrients and high temperature., 2006, 25(2): 96-107.
[14] Zahedi M, McDonald G, Jenner C F. Nitrogen supply to the grain modifies the effects of temperature on starch and protein accumulation during grain filling in wheat., 2004, 55: 551-564.
[15] Ordoñez R A, Savin R, Cossani C M, Slafer G A. Yield response to heat stress as affected by nitrogen availability in maize., 2015, 183: 184-203.
[16] Elía M, Slafer G A, Savin R. Yield and grain weight responses to post-anthesis increases in maximum temperature under field grown wheat as modified by nitrogen supply., 2018, 221: 228-237.
[17] Altenbach S B, Dupont F M, Kothari K, Chan R, Johnson E L, Lieu D. Temperature, water and fertilizer influence the timing of key events during grain development in a US spring wheat., 2003, 37(1): 9-20.
[18] 姜丽娜, 李冬芬, 李春喜, 邵云. 小麦蔗糖合成酶和腺苷二磷酸葡萄糖焦磷酸化酶的研究进展. 作物杂志, 2008(6): 11-15.
Jiang L N, Li D F, Li C X, Shao Y. Recent advances on wheat sucrose synthase and ADP-glucose pyrophosphorylase., 2008(6): 11-15. (in Chinese)
[19] 刘霞, 尹燕枰, 姜春明, 贺明荣, 王振林. 花后不同时期弱光和高温胁迫对小麦旗叶荧光特性及籽粒灌浆进程的影响. 应用生态学报, 2005, 16(11): 2117-2121.
LIU X, Yin Y P, Jiang C M, He M R, Wang Z L. Effects of weak light and high temperature stress after anthesis on flag leaf chlorophyll fluorescence and grain fill of wheat., 2005, 16(11): 2117-2121. (in Chinese)
[20] 陆伟婷. 冬小麦产量和品质对花前夜间增温及不同施氮量的响应特征[D]. 南京: 南京农业大学, 2017.
Lu W T. Response characteristics on different nitrogen rates under night warming of grain yield and quality of winter wheat[D]. Nanjing: Nanjing Agricultural University, 2017. (in Chinese)
[21] 陈洋, 赵宏伟. 氮素用量对春玉米穗位叶蔗糖合成关键酶活性的影响. 玉米科学, 2008, 16(1): 115-118.
Chen Y, Zhao H W. Effect of nitrogen application on activities of key enzymes of sucrose synthesis in the leaf located near the ear of spring maize., 2008, 16(1): 115-118. (in Chinese)
[22] 赵宏伟, 邹德堂, 马凤鸣. 施氮量对不同品种春玉米穗位叶蔗糖合成的影响. 中国农学通报, 2005, 21(10): 196-199.
Zhao H W, Zou D T, Ma F M. Effect of different nitrogen utilization on sucrose synthesizing in spike leaf of different spring maize., 2005, 21(10): 196-199. (in Chinese)
[23] RACHANA D, HIMANSHU P, SHIVDHAR S, BIDISHA C, AMOD K T, RAM K F. Impact of sowing dates on terminal heat tolerance of different wheat (L. ) cultivars., 2019, 42(7): 445-449.
[24] 邵宇航, 石祖梁, 张姗, 贾涛, 王飞, 戴廷波. 高温胁迫下镁对小麦旗叶光合特性及产量的影响. 麦类作物学报, 2018, 38(7): 802-808.
Shao Y H, Shi Z L, Zhang S, Jia T, Wang F, Dai T B. Effect of magnesium rates on photosynthetyic characteristics of flag leaf and grain yield in winter wheat under heat stress., 2018, 38(7): 802-808. (in Chinese)
[25] 王东, 徐学欣, 张洪波, 林祥, 赵阳. 微喷带灌溉对小麦灌浆期冠层温湿度变化和粒重的影响. 作物学报, 2015, 41(10): 1564-1574.
Wang D, Xu X X, Zhang H B, Lin X, Zhao Y. Effects of irrigation with micro-sprinkling hoses on canopy temperature and humidity at filling stage and grain weight of wheat., 2015, 41(10): 1564-1574. (in Chinese)
[26] 曹云英. 高温对水稻产量与品质的影响及其生理机制[D]. 扬州: 扬州大学, 2009.
Cao Y Y. Effect of high temperature on the quality and quantity of rice yield and its physiological mechanism[D]. Yangzhou: Yangzhou University, 2009. (in Chinese)
[27] 姜东, 谢祝捷, 曹卫星, 戴廷波, 荆奇. 花后干旱和渍水对冬小麦光合特性和物质运转的影响. 作物学报, 2004, 30(2) : 175-182.
Jiang D, Xie Z J, Cao W X, Dai T B, Jing Q. Effects of post- anthesis drought and water-logging on photosynthetic characteristics, assimilates transportation in winter wheat., 2004, 30: 175-182. (in Chinese)
[28] Rufty T W, Kerr P S, Huber S C. Characterization of diurnal changes in activities of enzymes involved in sucrose biosynthesis., 1983, 73(2) : 428-433.
[29] Sebkova V, Unger C, Hardegger M, Sturm A. Biochemical, physiological, and molecular characterization of sucrose synthase from daucus carota., 1995, 108(1) : 75-83.
[30] Liu L T, Hu C S, Olesen J E, Ju Z Q, Yang P P, Zhang Y M. Warming and nitrogen fertilization effects on winter wheat yields in northern China varied between four years., 2013, 151: 56-64.
[31] 赵晶晶. 花后高温胁迫下不同施氮量对春小麦产量形成的影响机理[D]. 银川: 宁夏大学, 2015.
ZHAO J J. Effect of different nitrogen application on spring wheat yield formation under high temperature stress after flowering[D]. Yinchuan: Ningxia University, 2015. (in Chinese)
[32] 牟会荣, 姜东, 戴廷波, 荆奇, 曹卫星. 遮荫对小麦旗叶光合及叶绿素荧光特性的影响. 中国农业科学, 2008, 41(2): 599-606.
Mu H R, Jiang D, Dai T B, Jing Q, Cao W X. Effect of shading on photosynthesis and chlorophyll fluorescence characters in wheat flag leaves., 2008, 41(2): 599-606. (in Chinese)
[33] Talukder A S M H M, McDonald G K, Gill G S. Effect of short-term heat stress prior to flowering and early grain set on the grain yield of wheat., 2014, 160: 54-63.
[34] Wang X, Cai J, Jiang D, Liu F, Dai T, Cao W. Pre-anthesis high-temperature acclimation alleviates damage to the flag leaf caused by post-anthesis heat stress in wheat., 2011, 168: 585-593.
[35] 刘霞. 小麦淀粉品质形成机制及其对温光因子的响应[D]. 泰安: 山东农业大学, 2005.
Liu X. Mechanism and response to temperature and light of starch quality formation in wheat[D]. Taian: Shandong Agricultural University, 2005. (in Chinese)
Effects of nitrogen application rate on assimilate accumulation, transportation and grain yield in wheat under high temperature stress after anthesis
GAO Chunhua1, 2, FENG Bo1, CAO Fang1, LI Shengdong1, WANG Zongshuai1, ZHANG Bin1, WANG Zheng1, KONG Lingan1, WANG Fahong1
(1Crop Research Institute, Shandong Academy of Agricultural Sciences, Ji’nan 250100;2Cotton Research Center, Shandong Academy of Agricultural Sciences,Ji’nan 250100)
【】This study was aimed to identify the effects of nitrogen application rates on grain yield, assimilate accumulation and translocation, physiological basis of winter wheat under the condition of high temperature stress, so as to provide technical and theoretical support for stress-resistance and stable yield cultivation.【】 Field experiments were conducted at Ji’nan experiment station and Jiyang experiment station of Crop Research Institute, Shandong Academy of Agricultural Sciences in 2018 and 2019, designed with two temperature treatments (CK: Control, H: High temperature stress) and three nitrogen levels (N1:Low nitrogen with 180 kg·hm-2, N2: Regular nitrogen with 240 kg·hm-2, and N3: High nitrogen with 300 kg·hm-2) . By analyzing assimilate accumulation, allocation, sucrose synthetase activity in leaf and grain, and grain yield with winter wheat cultivar JM44, the regulation of nitrogen application rates on grain yield of wheat under high temperature stress circumstances was studied. 【】 The results showed that grain yield decreased significantly under high temperature stress conditions, and grain yield reduction at N1, N2, and N3 were 54.78%, 24.05%, 54.49% and 50.19%, 25.29%, 44.13% in Ji’nan and Jiyang, respectively. Significant increases were noticed in assimilate accumulation amount, pre-anthesis assimilate translocation amount and rate to grain yield, post-anthesis assimilate accumulation amount and rate, ratio of grain to other organs udner N2 treatment, compared with N1 and N3 treatment, same as in SPAD value, sucrose synthetase-I activity (direction of synthetic) in flag leaf, and sucrose synthetase-Ⅱ activity (direction of decomposition) in grain. 【】Higher grain yield could be achieved after high temperature stress during grain-filling stages with nitrogen application rate of 240 kg·hm-2, which could postpone flag leaf senescence, maintain high sucrose synthetase-I activity in flag leaf and sucrose synthetase-Ⅱ activity in grain, and keep high ability cof assimilate accumulate and transport to grain.
winter wheat; nitrogen application rate; high temperature stress; assimilate accumulation; assimilate transportation; grain yield

10.3864/j.issn.0578-1752.2020.21.006
2020-05-11;
2020-07-29
国家重点研发计划(2018YFD0300600-1)、国家公益性行业(农业)科研专项(201503130)、山东省重点研发计划(2017GNC11106)、山东省农业科学院农业科技创新工程项目(CXGC2016B04)、山东省农业科技资金(园区产业提升工程)项目(2019YQ001)
高春华,E-mail:chunhuaaa009@163.com。通信作者冯波,E-mail:fengbo109@126.com
(责任编辑 杨鑫浩)
猜你喜欢
中国果业信息(2022年1期)2022-11-28
作物学报(2022年1期)2022-11-05
作物学报(2022年9期)2022-07-18
作物学报(2022年9期)2022-07-18
农业工程学报(2022年7期)2022-07-09
华北农学报(2022年2期)2022-05-12
干旱地区农业研究(2021年6期)2021-12-08
今日农业(2021年17期)2021-11-26
养殖与饲料(2021年4期)2021-04-17
河南农业·综合版(2020年10期)2020-10-20
