
烟草NtbHLH112 基因的克隆、鉴定及表达模式分析
2020-11-04 08:16孙晋浩牛文利陈志华余祥文丁安明杨兴有孙玉合
中国烟草科学 2020年5期
孙晋浩,牛文利,陈志华,余祥文,丁安明,杨兴有,孙玉合*
(1.中国农业科学院烟草研究所,烟草行业烟草基因资源利用重点实验室,青岛 266101;2.四川省烟草科学研究所,成都 610000)
转录因子能与顺式作用元件相互作用,特异性调控与环境应激相关的基因,以维系植物正常生命活动[1-2]。bHLH(basic helix loop helix)转录因子含有高度保守的bHLH 结构域,是植物体内第二大类转录因子[3]。其结构域由大约60 个氨基酸组成,包括碱性氨基酸区域和螺旋-环-螺旋区域(HLH)。碱性氨基酸区域位于bHLH 结构域的N-末端,包含15 个氨基酸,其功能主要是识别和特异性结合启动子中的碱基序列[4];螺旋-环-螺旋区位于bHLH 结构域的C 端,约含有45 个氨基酸,其主要功能是促进蛋白质与蛋白质的互相作用,并允许形成同源二聚体或异二聚体复合物来控制基因转录[5]。尽管该类转录因子含有高度保守的bHLH 结构域,但其他序列的保守性相对较低,存在较大差异性[6]。
bHLH 转录因子家族成员在植物抗逆应答过程中起到极其重要的调控作用[7-11]。已有研究表明在拟南芥以及水稻、苦荞麦、山茶树等重要的粮食作物和经济作物中鉴定得到bHLH 转录因子[10,12-14]。例如,拟南芥中的bHLH92、bHLH112 和bHLH122转录因子响应干旱、盐、渗透等多种胁迫[15-17]。水稻中的bHLH 转录因子OsbHLH068 和OsbHLH148参与干旱胁迫响应过程[18-19]。烟草同源作物辣椒和番茄中也鉴定到bHLH 转录因子CabHLH40 和SlICE1a 受干旱、盐、ABA 非生物胁迫诱导[20-21]。
烟草的抗旱、抗冷和抗盐性对烟草的产量和品质起到极为重要的作用[22],深入研究烟草的抗旱和抗盐的调控机理,能够为烟草育种提供重要的理论依据[23-25]。但是在烟草中关于bHLH 转录因子家族的研究却相对较少。目前,仅有一个烟草bHLH 类转录因子NtbHLH123 被研究。该转录因子可以直接作用于NtCBF基因启动子并正向调节其表达,进而调控烟草的耐冷性;同时过表达NtbHLH123基因的烟草植株在冷胁迫下表现为较低的电解质渗透,较低含量丙二醛、H2O2和活性氧(ROS),从而减轻冷胁迫条件下的细胞膜氧化损伤[26]。本研究基于已经报道的拟南芥bHLH112基因序列和茄科基因组数据库中的番茄SlbHLH112、辣椒CabHLH112的基因信息,在普通烟草中克隆NtbHLH112基因,并运用生物信息学分析和亚细胞定位等试验手段对其保守结构域、进化关系、亚细胞定位、转录激活以及基因的表达模式进行分析,预测其生物学功能,为烟草抗逆育种提供理论依据。
1 材料与方法
1.1 试验材料
普通烟草品种K326。
1.2 试剂和仪器
菌株:大肠杆菌菌株(E.coli)DH5α、农杆菌菌株EHA105、酵母AH109 菌株由本实验室保存。
载体:pYG51-GFP 和pBridge 由本实验室保存或改造。
酶类:试验所用限制性内切酶、Pfu 高保真酶、Easy Taq 酶、Infusion 连接酶购自宝生物工程有限公司(TaKaRa)。
其他试剂:RNA 提取试剂盒、cDNA 反转录试剂盒、DNA PCR 产物纯化试剂盒、质粒小量提取试剂盒、pEASY-Blunt 载体、SYBR® Premix Ex TaqTM荧光定量试剂购于全式金生物技术有限公司;酵母基本培养基(SD Base)、色氨酸缺素培养基(-Trp)、X-Gal 等购于Takala 公司;Amp、β-巯基乙醇、Kan、EB、琼脂粉、SDS、琼脂糖等购自索莱宝生物公司;NaOH、盐酸、LiCl、氯仿、NaCl、无水乙醇、异丙醇等为国产分析纯试剂。
仪器:荧光定量PCR 仪ABI-Q3(美国ABI 公司生产),激光共聚焦显微镜TCS-SP8(德国Leica公司生产)。
1.3 试验方法
1.3.1 烟草材料种植和处理 (1)正常生长的烟草材料。烟草品种K326 种子点播于花盆中,种植于中国农业科学院烟草研究所气候室。在烟株盛花期,收集植株根、茎、叶、花、花蕾、腋芽等组织,液氮速冻后放置于-80 ℃封存备用。收集根部组织时,将烟草根系部位营养土用水轻轻冲洗干净,剪取侧根,迅速置于液氮中;叶组织分别收集上部叶、中部叶、下部叶。(2)不同逆境胁迫下的烟草材料。用MS 固体培养基萌发幼苗,待长至6 片真叶期,进行盐、干旱、温度、激素处理。盐处理:将6 片真叶期苗转移至含有60 mmol/L NaCl 溶液处理1、3、6 h,取整株,以MS 盐溶液处理作为对照。干旱处理:将6 片真叶期苗转移至吸水滤纸表面进行干旱处理1、3、6 h[25],取整株,用未处理的烟苗作为对照。温度处理:将6 片真叶期苗转移至4 ℃和37 ℃培养箱中进行处理1、3、6 h,取整株,以25 ℃处理作为对照。激素处理:将6 片真叶期苗转移至含有10 μmol/L ABA 的MS 盐溶液处理1、3、6 h,取整株,以MS 盐溶液处理作为对照。处理完成后,每组选取3 株烟苗,液氮速冻后放置于-80 ℃封存备用。
1.3.2 RNA 提取及反转录PCR 将备用样品在含有液氮的研钵中研成粉末,依试剂盒说明书步骤,提取总RNA,-80 ℃保存备用;依反转录试剂盒说明书反转录为cDNA,稀释至10 ng/μL,-20 ℃封存,备用。
1.3.3 基因NtbHLH112的克隆 以拟南芥AtHLH112 蛋白序列作为query 序列,在茄科基因组数据库(http://solgenomics.net/)中进行BLAST 比对,在该数据库(Database)中选择N.tabacumK326 protein sequences,获得候选基因(Ntab0435090),依据候选基因的CDS 序列,设计特异性PCR 扩增引物。上游引物: 5'-ATGGCTGATGAATTTC AAGCAGG-3',下游引物:5'-TTATCTGAATGTA CCACCAAAATTTGG-3',以普通烟草 K326 总cDNA 为模板,使用Pfu 高保真酶扩增目的基因,然后用纯化试剂盒回收目的片段。取5 μL 纯化基因片段连接于pEASY-Blunt 载体并转入大肠杆菌感受态中,用含100 μg/mL 卡那霉素的LB 平板筛选,将阳性克隆送至上海生工生物科技有限公司进行测序,保存测序正确的菌种,并命名为Blunt-NtbHLH112。
1.3.4 NtbHLH112 的生物信息学分析 利用ProtParam 程序(https://web.expasy.org/protparam/)预测NtbHLH112基因编码的蛋白氨基酸序列的分子量、等电点、蛋白亲水值;在拟南芥数据库TAIR(www.arabidopsis.org)中下载 AtbHLH112 和AtbHLH122,在茄科基因组数据库(http://solgenomics.net/)中下载番茄SlHLH112、辣椒CaHLH112,在NCBI 数据库(https://www.ncbi.nlm.nih.gov/)中下载玉米 ZmbHLH112、水稻OsbHLH112、大豆GmbHLH112、葡萄VitbHLH112,利用DNAMAN 进行多序列比对,使用WebLogo(http://weblogo.berkeley.edu/logo.cgi)将结果可视化,分析NtbHLH112 的保守结构域;利用MEME(http://meme-suite.org/)在线软件分析保守基序(motif),使用MGEA7 建立进化树图谱(Neighbor-Joining,NJ tree)。
1.3.5 NtbHLH112 的亚细胞定位分析 以 Blunt-NtbHLH112亚克隆载体质粒作为模板利用Pfu 高保真酶PCR 扩增。将NtbHLH112基因CDS 序列使用Infusion 重组酶连接到pYG51-GFP 亚细胞定位的载体上,转化至大肠杆菌感受态,置于含150 μg/mL壮观霉素的LB 平板筛选,获得阳性克隆测序,将正确的重组载体转入农杆菌感受态EHA105,置于含150 μg/mL 壮观霉素和60 μg/mL 利福平的LB 平板筛选,PCR 阳性鉴定。根据李晓旭等[26]方法注射本氏烟叶,获取注射材料后置于激光共聚焦显微镜观察GFP 荧光信号。
1.3.6 NtbHLH112 的转录激活试验 以Blunt-NtbHLH112亚克隆载体质粒作为模板利用Pfu 高保真酶PCR 扩增,将NtbHLH112基因CDS 序列使用Infusion 重组酶连接到pBridge 载体上,测序获得阳性克隆,并构建NtbHLH112与GAL4-BD 的融合表达载体。将融合载体与pBridge 空白载体分别转化酵母AH109菌株,在SD/-Trp缺陷培养基上培养3 d,将阳性克隆转移至SD/-Trp/X-gal 显色培养基上避光培养至显色。
1.3.7NtbHLH112表达模式分析 使用qRT-PCR反应分析NtbHLH112基因的表达模式。利用NCBI设计特异性扩增qRT-PCR 引物,上游引物:5'-CACCCTTGTTGAAACCTTCT-3',下游引物:5'-TCAACATCCGACTTGGCAGA-3',以烟草 26S rRNA 基因作为内参对照,上游引物:5'-CAAGGAAATCACCGCTTTGG-3',下游引物:5'-CAAGGAAATCACCGCTTTGG-3'。经胁迫、激素处理的所有样品提取RNA,反转录成为cDNA,然后进行qRT-PCR,依据2-ΔΔCt算法处理数据。
2 结果
2.1 NtbHLH112 基因CDS 序列克隆及分析
根据基因组数据库BLASTP 比对结果,进行候选基因的特异性扩增引物设计,以K326 植株的全cDNA 作为模板扩增,目的片段约1300 bp(图1)。然后将片段纯化回收,连接至Peasy-Blunt 载体,挑取单克隆测序,结果表明,NtbHLH112基因CDS 序列全长1368 bp,共编码455 个氨基酸。ProtParam分析显示该蛋白相对分子量为50.46 kDa,理论等电点为6.39,蛋白亲水值(Protein GRAVY)为-0.72。

图1 NtbHLH112 基因PCR 扩增产物电泳图Fig.1 Electrophoretogram of NtbHLH112 PCR amplification
依据NtbHLH112 的蛋白序列,将其与拟南芥、番茄和玉米等物种中的bHLH112 蛋白进行多序列比对(图2),高度保守的氨基酸用深蓝标记,其他相对保守的氨基酸用红色以及浅蓝色标注,发现NtbHLH112在N端具有高度保守的bHLH结构域。
2.2 NtbHLH112 转录因子进化分析及保守基序分析
以拟南芥、番茄和玉米等物种中的bHLH112 转录因子作为参考序列进行比对,运用MEGA 7.0 建立进化树(图3)。基于进化树分析,烟草NtbHLH112与番茄SlbHLH112、辣椒CabHLH112 聚在一起,表明三者是直系同源。利用保守序列识别工具MEME 分析,结果显示同一支的bHLH112 转录因子成员具有较强的一致性。并且,NtbHLH112 与番茄SlbHLH112、辣椒CabHLH112 保守基序的种类以及组织形式高度一致,暗示NtbHLH112 与番茄SlbHLH112、辣椒CabHLH112 可能具有功能相似性。

图2 NtbHLH112 转录因子与其他植物 bHLH 蛋白的氨基酸序列比较Fig.2 Multiple sequence alignment of NtbHLH112 and bHLH proteins of other plant species

图3 NtbHLH112 转录因子系统进化及保守基序分析Fig.3 Phylogenetic and conserved motifs analysis of NtbHLH112 transcription factor
2.3 NtbHLH112 转录因子亚细胞定位
构建35s::NtbHLH112::GFP融合载体,利用农杆菌注射本氏烟叶片,瞬时表达NtbHLH112,以检测NtbHLH112 转录因子的亚细胞定位(图4),试验发现,融合GFP 的NtbHLH112 只在细胞核中被观察到,并且能够与DAPI 染色信号重叠,这个结果表明NtbHLH112 是一个核定位蛋白。
2.4 NtbHLH112 转录因子转录激活分析
根据转录因子转录激活原理[26-27],将NtbHLH112-pBridge 载体转入AH109 酵母感受态,pBridge 空载作对照,在SD/-Trp 缺素平板培养基上培养3 d 筛选阳性酵母单克隆并转移至显色培养基(SD/-Trp/X-gal)上避光培养直至显色。试验结果表明,含有NtbHLH112-pBridge 载体酵母表达菌株在显色培养基上能够启动报告基因的表达进而显蓝色(图5)。而只含有pBridge 空载的对照不能激活报告基因表达,说明NtbHLH112 具有转录激活活性。
2.5 NtbHLH112 表达模式分析
RT-qPCR 结果表明,NtbHLH112在根(R)、茎(ST)、幼叶(YL)、成熟叶(NS)、衰老叶(SL)、全花(F)、花蕾(FB)、腋芽(LB)组织中都有表达,在叶中相对表达量较高,特别是在衰老叶片中表达量最高(图6)。

图4 NtbHLH112 的亚细胞定位Fig.4 Subcellular localization of NtbHLH112

图5 NtbHLH112 酵母转录激活试验Fig.5 Transactivation analysis of NtbHLH112 in yeast
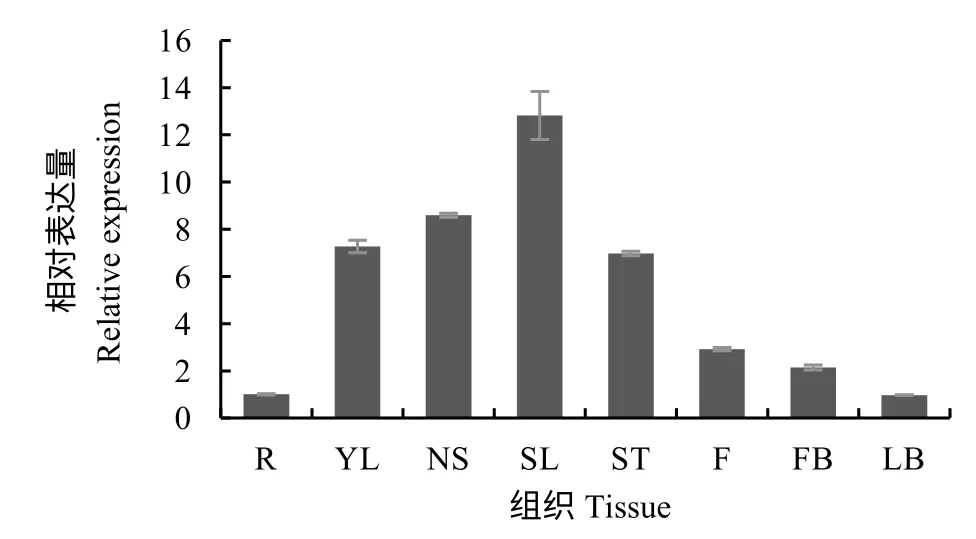
图6 NtbHLH112 在烟草不同组织的表达量Fig.6 Expression of NtbHLH112 in different tissues of tobacco
表达模式分析结果表明(图7),NtbHLH112基因表达响应多种胁迫以及ABA 处理,其中响应干旱胁迫最为明显。在干旱胁迫处理条件下,NtbHLH112基因表达量在干旱处理1 h 时最高,是0 h 的5 倍,随着时间推移表达量开始降低,即使在处理6 h 后,表达量仍然为0 h 的2 倍。在盐胁迫处理条件下,处理1 h,该基因表达量显著增高,是未处理的2 倍,随着时间的推移,表达量急剧降低,在3 h 时,表达量已经低于0 h。冷处理后该基因随着时间的推移表达量有增高,但低于两倍。在热胁迫中,表达量基本不变。此外,ABA 处理中,随着时间的推移,该基因的表达量显著降低,在处理6 h 后,表达量降低了一半多。说明,NtbHLH112基因主要受干旱和盐胁迫诱导,同时在ABA 调控途径的非生物逆境应答过程中发挥作用。

图7 NtbHLH112 在非生物胁迫下的表达分析Fig.7 Analysis of expression of NtbHLH112 gene under abiotic stresses
3 讨 论
bHLH 转录因子在植物的生长发育、生理代谢及胁迫过程中起着重要的调控作用。bHLH 转录因子都具有一个高度保守的典型bHLH 结构域。本试验从普通烟草中克隆获得NtbHLH112基因,通过生物信息学分析发现其确实具有高度保守的bHLH 结构域。转录因子必备的特性是其亚细胞定位应在细胞核且具有转录激活活性[26-27],NtbHLH112 亦符合以上特性。因此NtbHLH112 属于bHLH 转录因子家族。不同物种中很多bHLH 转录因子都受干旱诱导[15-17,28-29],表达量升高从而提高植株的耐旱性。本试验中在干旱胁迫处理条件下,NtbHLH112基因表达显著增高,说明NtbHLH112也受干旱诱导。有试验表明胡杨PebHLH35 转录因子能够调节植株叶片的气孔密度,使其开度显著降低,蒸腾速率下降,以提高植物的干旱耐受性[29],而基因表达模式分析表明,NtbHLH112基因主要在叶片中表达,特别是在成熟叶片中的表达量明显上升,这表明NtbHLH112基因可能参与一些叶片感知的干旱胁迫。作为NtbHLH112的同源基因,拟南芥AtbHLH112和AtbHLH122,水稻OsbHLH068基因均响应盐胁迫,并且在植物不同组织的不同部位均有表达,特别是叶片[17-18]。在盐胁迫处理条件下,NtbHLH112基因表达增高,说明NtbHLH112基因在响应植物的抗盐胁迫过程的模式与拟南芥和水稻相似。ABA 作为一个重要的信号分子参与植物的胁迫响应,GhbHLH1和AtMYC2经ABA 处理后表达量升高,以提高植物抗性[11,28];而NtbHLH112基因与以上两个基因不同,经ABA 处理后,随着时间的推移表达量显著降低,即ABA 能够负调控NtbHLH112的表达。
4 结 论
本研究初步验证烟草NtbHLH112 属于典型的bHLH 转录因子,在烟草叶片中高量表达,并受到干旱胁迫、盐胁迫的诱导,同时在ABA 调控途径的非生物逆境应答方面发挥重要作用。本研究为烟草抗逆育种提供了重要的理论依据。
猜你喜欢
环球时报(2022-09-20)2022-09-20
湖北农业科学(2022年11期)2022-07-18
河北农业大学学报(2022年2期)2022-04-26
园艺与种苗(2020年12期)2021-01-08
今日农业(2020年24期)2020-12-15
实用肿瘤学杂志(2020年4期)2020-12-08
飞碟探索(2015年9期)2015-11-05
小资CHIC!ELEGANCE(2015年14期)2015-09-23
红领巾·探索(2015年9期)2015-09-10
小资CHIC!ELEGANCE(2015年15期)2015-09-01
