
支持高密度BHK-21细胞培养和高产FMD病毒的无血清培养基开发与优化
2020-11-02 08:49陈敏刘旭平赵亮
生物技术通报 2020年10期
陈敏 刘旭平 赵亮
(1. 上海倍谙基生物科技有限公司,上海 201203;2. 华东理工大学 生物反应器工程国家重点实验室,上海 200237)
口蹄疫是一种具有高度传染性的偶蹄类动物疫病,由口蹄疫病毒(含多种血清型:O、A、C、SAT1[Southern african territories 1)、SAT2、SAT3和Asia1]引起,猪、牛、羊等养殖畜类及水牛等野生动物易感,感染发病率100%,其爆发会对畜牧业造成严重的损失。目前最为有效的防控手段是口蹄疫疫苗的接种[1-5]。畜牧业的发展、种群密度的加大都增加了防疫的难度,也进一步提高了对疫苗的需求。近年来,各类新型疫苗不断涌现,大量文献介绍了这些新型疫苗所能解决的、现有疫苗存在的问题[6-10]。但是,目前被广泛使用的疫苗仍以灭活病毒疫苗为主。考虑到灭活疫苗价格昂贵、保质期短、保护作用短、需要重复接种的问题[7-10],提高现有工艺的疫苗生产能力,加大产能,降低生产成本,无疑是解决当前疫苗需求困境的最佳策略。从生产工艺的角度,BHK-21细胞悬浮培养工艺优于贴壁生产工艺并将逐步取代后者已在领域内达成共识,急需解决的是血清引入可能导致的安全隐患和完全无血清体系下支持高密度细胞培养和高效病毒扩增的问题。
目前利用BHK-21悬浮细胞生产口蹄疫病毒疫苗使用的接毒细胞密度通常在3×106cells/mL-5×106cells/mL,根据病毒血清型不同,收获病毒完整颗粒在1 μg/mL-10 μg/mL不等。“细胞密度效应”是指基于动物细胞培养技术的病毒生产工艺中发生的病毒收获量不能随接毒时细胞密度增高而增高,甚至出现下降的现象[11-14],是生产工艺在扩大规模和提高产能时的主要瓶颈。这一现象最有可能的原因是高密度细胞培养导致体系内重要营养物质缺乏[15](如葡萄糖、氨基酸)、有毒代谢副产物积累[16-17](如乳酸、氨)以及培养环境参数改变[18-19](如pH),进而影响了细胞的生理状态以及后续的细胞产毒过程。
本单位前期已开展了大量开发工作,获得了能够支持BHK-21悬浮细胞稳定传代和口蹄疫病毒扩增的多个无血清培养基配方和生产工艺[20-22]。然而,在实际应用时仍需添加低浓度的血清以达到最佳的病毒产量和病毒效价[20-21]。为实现完全的无血清培养工艺,本研究首先将BHK-21细胞培养于前期开发的一款无血清培养基中[20],观测细胞生长和接毒时的生理代谢状态,通过代谢通量分析,了解细胞生长和病毒扩增时的营养需求以及可能导致有毒代谢物积累或培养环境恶化的原因,识别关键的营养成分,采用合理的实验设计,旨在得到一款能够支持BHK-21细胞高密度悬浮生长和口蹄疫病毒高效扩增的无血清培养基,从而取代现有的低血清培养工艺,并获得超高细胞密度接毒(>107cells/mL)、病毒高产量和简化接毒操作的新工艺。
1 材料与方法
1.1 材料
1.1.1 细胞株 BHK-21贴壁细胞购自中国科学院上海生命科学研究院,使用本单位开发的无血清培养基悬浮驯化,并经单克隆筛选后获得悬浮细胞株。
1.1.2 培养基 优化前的无血清培养基为本单位自主开发,命名为primary SFM(简称P-SFM),包含葡萄糖、氨基酸、维生素、微量元素以及多种血清替代物等营养物质。
低血清培养基(简称LSM)为优化前的无血清培养基加1%新生牛血清。
TCID50测试中使用的DMEM购自Sigma。
1.1.3 毒株 O型口蹄疫毒株,悬浮细胞生产获得,由合作客户提供,种毒TCID50值为7.25 lgTCID50/0.1mL。
1.2 方法
1.2.1 BHK-21细胞在摇瓶体系中的培养 取复苏后可稳定传代且活性大于95%的BHK-21细胞用于本研究。
取处于对数生长期、活性大于95%的足量细胞置于50 mL离心管,1 000 r/min离心5 min后弃上清,加入30 mL实验组无血清培养基后重悬细胞并转移至125 mL摇瓶内。摇床转速设定为130 r/min,培养条件为37℃、5% CO2、饱和湿度。
批培养时,每24 h取样计数,样品于10 000 r/min下离心5 min,保留上清冻存于-20℃,用于营养代谢物的分析。
传代培养或扩培时,每48 h将细胞液用新鲜的无血清培养基稀释至活细胞密度为1×106cells/mL,培养体积根据传代或扩培的需要进行调整。
1.2.2 葡萄糖浓度的优化 依据BHK-21细胞在P-SFM中的生长和代谢情况,以指数期葡萄糖的比消耗速率(1.07 mmol/109cells·day)和预期达到的最高活细胞密度(2×107cells/mL)进行计算,葡萄糖需增加的浓度约为8 mmol/L(公式1)。为避免乳酸过量累积,本节设计在初始浓度的基础上分别增加5 mmol/L和10 mmol/L葡萄糖进行考察。
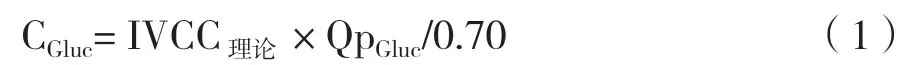
1.2.3 氨基酸浓度的优化 依据BHK-21细胞在P-SFM中的生长和代谢情况,对谷氨酰胺、天冬氨酸、天冬酰胺3种氨基酸的浓度进行优化。假设活细胞密度和指数期(D0-D3期间)平均比消耗速率均保持不变,以第3天各氨基酸剩余浓度为初始浓度的30%、45%为准,算得各氨基酸的推荐增加浓度(表1)。计算公式如下:


表1 氨基酸浓度优化的理论添加量
依据表1推荐的浓度,该部分实验分别将3种氨基酸按照原浓度增加60%和30%作为两个浓度水平,高浓度记为“+”,低浓度为“-”,设计如表2。

表2 氨基酸浓度优化的实验设计
1.2.4 铜锌离子浓度的优化 铜离子、锌离子分别设计3个浓度水平进行考察,实验设计如表3所示。
1.2.5 基于摇瓶培养体系的接毒操作 直接接毒:将BHK-21细胞以1×106cells/mL接种于摇瓶,待细胞扩增至0.9 cells/mL-1.0×107cells/mL,以MOI=0.1接种口蹄疫病毒,细胞病变90%以上时收获毒液。
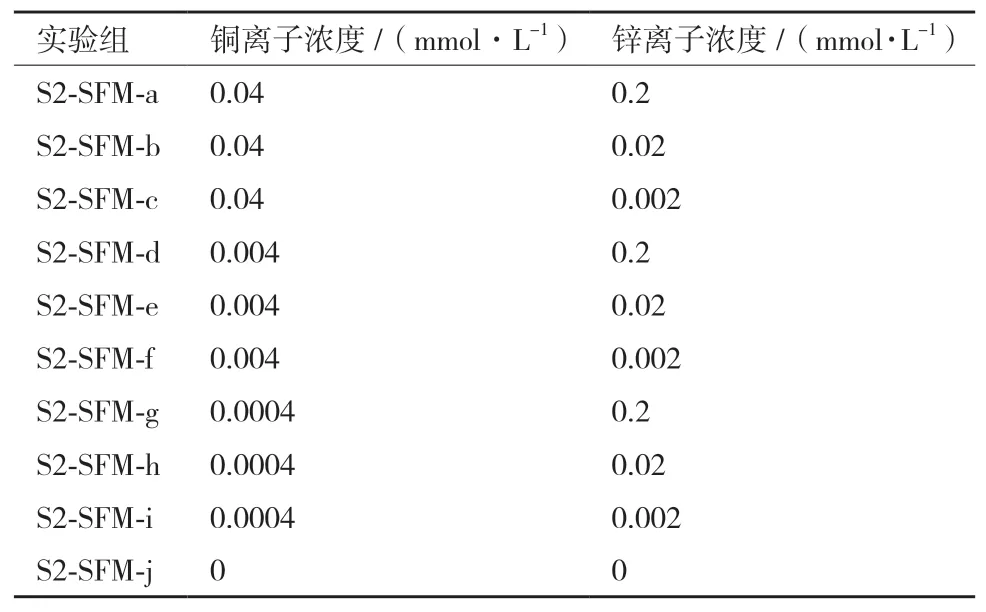
表3 铜锌离子浓度实验设计表
接毒前流加补液:将BHK-21细胞以1 cells/mL×106cells/mL接种于摇瓶,待细胞扩增至0.9 cells/mL-1.0×107cells/mL,流加20%浓缩培养基,并以MOI=0.1接种口蹄疫病毒,细胞病变90%以上时收获毒液。
接毒前离心换液:将BHK-21细胞以1 cells/mL×106cells/mL接种于摇瓶,待细胞扩增至0.9 cells/mL-1.0×107cells/mL,离心去上清,使用新鲜培养基重悬细胞,并以MOI=0.1接种口蹄疫病毒,细胞病变90%以上时收获毒液。
1.2.6 接毒时设定不同的活细胞密度 将BHK-21细胞以1×106cells/mL接种于摇瓶,48 h后取样计数,离心更换新鲜培养基,调整活细胞密度分别为3×106cells/mL、6×106cells/mL和9×106cells/mL,以MOI=0.1接种口蹄疫病毒,细胞病变90%以上时收获毒液。
1.2.7 BHK-21细胞在生物反应器中的培养 将BHK-21细胞以1×106cells/mL的活细胞密度接种于确认无菌的2 L生物反应器,培养体积1 L,每24 h取样计数。生物反应器的操作参数设置为:温度37℃,转速150 r/min,pH控制在7.2-7.4,溶氧设定为40%,表层通气控制在(50-100)mL/min,碱液使用1 mol/L的Na2CO3溶液。
1.2.8 基于生物反应器培养体系的接毒操作 按照方法1.2.7在2 L生物反应器中培养BHK-21细胞,48 h后4℃低温沉降换液,换液量约80%,按照MOI =0.1接种口蹄疫病毒,调节pH为7.2-7.4,接毒6 h后每2 h取样计数,待细胞病变90%以上,收集病毒液于样品瓶中,-80℃保存。留样用于测定TCID50和146s含量。
1.2.9 细胞计数 使用自动计数仪(IC1000,Countstar)进行细胞计数,并在计数前添加台盼蓝染液识别体系中的死细胞。
1.2.10 代谢物浓度检测 葡萄糖、乳酸和氨的浓度使用生化分析仪(BP400,Nova)进行检测;氨基酸浓度使用HPLC(1260,Agilent)基于OPA+FMOC在线衍生的方法进行检测。
1.2.11 TCID50的测定 病毒感染半数细胞培养物的稀释倍数常用TCID50进行表示,是病毒感染能力的重要指标。检测方法参考文献方法[20-21]。简言之,将不同稀释倍数的病毒液加入贴壁培养的单层BHK-21细胞,观察细胞的病变情况,并以下式计算TCID50:
其中,L为孔板中病变率在50%以上的最高稀释度对数;D为孔板中病变率在50%以上的最高稀释度对数与病变率低于50%的最低稀释度对数之间的差值;S为孔板中病变率的总和。
1.2.12 146s含量的测定 口蹄疫病毒完整粒子146s采用密度梯度离心法进行检测,参考文献方法[23]。
1.2.13 数据处理
(1)比生长速率
μ:比生长速率(day-1);X2、X1:两个采样点的活细胞密度(cells/mL);∆t:两个采样点的间隔时间(day)。
(2)活细胞密度对时间积分

IVCC:活细胞密度对时间的积分(109cells·day/L)。
(3)比生成/比消耗速率
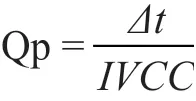
Qp:两个采样点之间葡萄糖的比消耗速率或乳 酸/氨/氨 基 酸 的 比 生 成 速 率(mmol/(109cells·day));∆p:两个采样点之间单位体积培养物中该物质的消耗量或生成量(mmol/L)。
(4)细胞病变率
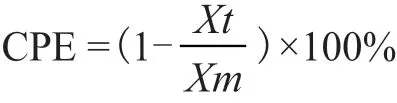
CPE:BHK-21细胞被口蹄疫病毒感染后的病变率(Cytopathic effect,%);Xt:t时刻培养体系中的活细胞密度(cells/mL);Xm:细胞从接种口蹄疫病毒到病变完成过程中的最大活细胞密度(cells/mL)。
(5)单细胞产毒能力
Svy:单位细胞产毒量(Cell-specific virus yield,virions/cell);146s:口蹄疫病毒完整粒子量(10-6g/mL);Mv:常数,单个完整病毒粒子的质量,1.146×10-17g/virion;Xm:细胞从接种口蹄疫病毒到病变完成过程中的最大活细胞密度(cells/mL)。
(6)数理统计 文章中的数据以均值±标准偏差进行标示,组间数据对比采用学生T检验法,当P<0.05时认为差异显著。
2 结果
2.1 BHK-21细胞在P-SFM中的生长和代谢
BHK-21细胞在P-SFM中进行批培养的生长和关键代谢情况如图1所示。0-2 d是明显的指数生长期,平均比生长速率0.96/day,第2-3天比生长速率下降为0.39/day,第3天最高活细胞密度达到13.7×106cells/mL并开始下降。培养至第3天时,葡萄糖的浓度为初始浓度的30%左右,不构成营养限制,而谷氨酰胺、天冬氨酸、天冬酰胺3种氨基酸的浓度不足初始浓度的15%(图1-B),有可能是细胞生长的限制因素。
如图1-A所示,0-3 d内,乳酸和氨处在持续积累的状态,乳酸最高浓度为26.6 mmol/L,氨为5.02 mmol/L,均已达到抑制细胞生长和影响病毒表达的水平[17,24-25]。有研究结果表明,不适当的营养成分补充[26-27](如葡萄糖、谷氨酰胺)和一些微量元素(如铜离子[28-29]和锌离子[30])会影响细胞的代谢和产毒过程。因此,针对此款无血清培养基,本研究从葡萄糖、氨基酸(谷氨酰胺、天冬氨酸、天冬酰胺)、微量元素(铜离子、锌离子)3个方面进行优化。

图1 BHK-21细胞在P-SFM批培养过程中的生长和代谢情况
2.2 无血清培养基的优化
如图2-A,葡萄糖浓度提高5 mmol/L和10 mmol/L都对细胞的生长有促进作用,两者之间无显著性差异,但乳酸的积累也随葡萄糖浓度的增高而增高。因此,优化后的培养基中葡萄糖浓度的增加量定为5 mmol/L。
氨基酸浓度优化的实验中,以指数期的细胞比生长速率和葡萄糖浓度为指标时,天冬氨酸、天冬酰胺和谷氨酰胺的浓度在较高浓度添加时都显示了微弱但不显著的优势。但是乳酸的累积和谷氨酰胺的增加呈正相关,且具有显著性差异(图2-B)。因此,优化后的培养基中天冬氨酸、天冬酰胺和谷氨酰胺的浓度分别以“+”、“+”、“-”的水平添加。
铜锌离子浓度优化的实验中,葡萄糖、谷氨酰胺的消耗、乳酸、氨的累积均没有显著性差异,而以0-3 d细胞总IVCC作为指标筛选出的最佳铜离子浓度为0.04 mmol/L,锌离子浓度为0.2 mmol/L,且锌离子浓度对细胞生长的影响更为显著(图2-C)。
调整以上成分的浓度后,新的无血清培养基被命名为Opt-SFM。
2.3 BHK-21细胞在Opt-SFM中的生长和代谢
摇瓶体系中,BHK-21细胞在Opt-SFM中0-3 d的细胞总IVCC显著高于P-SFM组(图3-A);Opt-SFM支持细胞稳定传代,48 h内的比生长速率为1.07-1.13/day(图3-C),传代过程中细胞活性始终高于98%;与P-SFM相比,Opt-SFM中的葡萄糖比消耗速率和乳酸比生成速率显著降低(图3-B);在Opt-SFM中,细胞形态饱满、尺寸均匀,且结团情况极少(图3-D)。
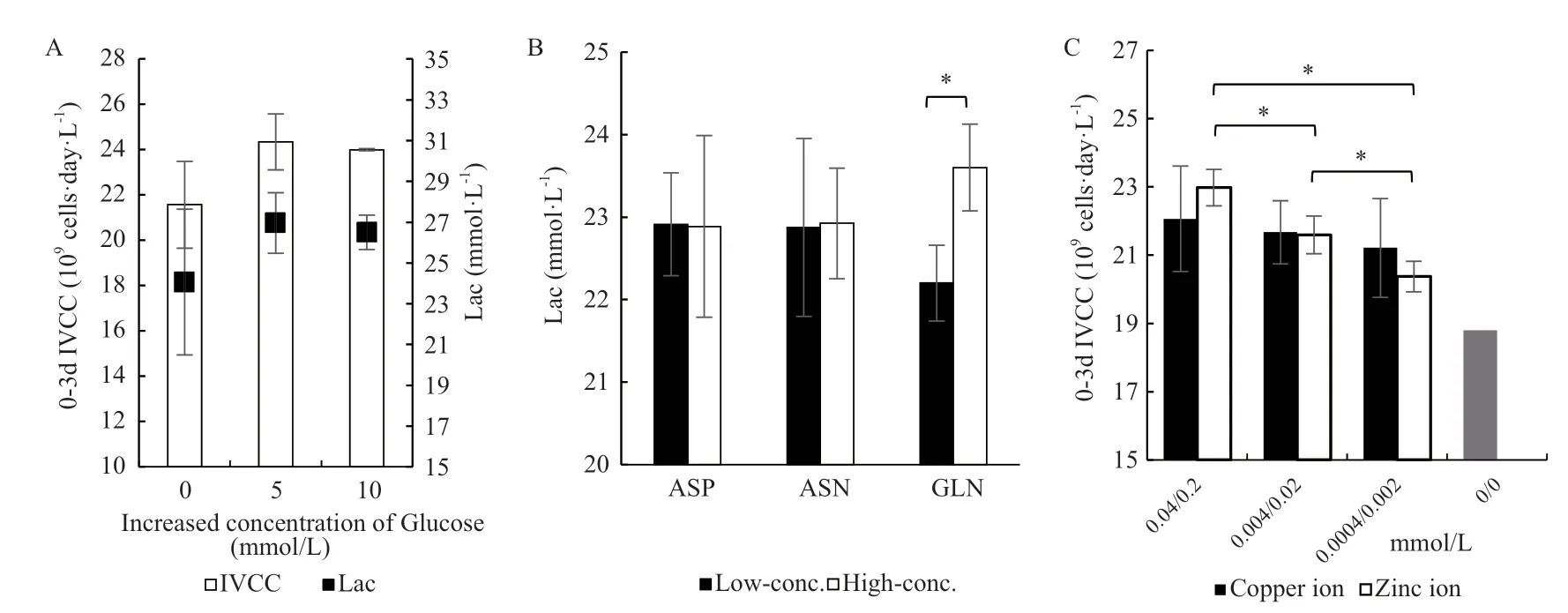
图2 无血清培养基的优化
反应器体系内,细胞的生长情况相比摇瓶体系有了更大的提升,如图4-A所示,细胞在Opt-SFM中最高活细胞密度达到17×106-18×106cells/mL(最高纪录为17.8×106cells/mL),与P-SFM相比提高约23.5%;前3 d的葡萄糖比消耗速率和乳酸、氨的比生成速率皆低于P-SFM(图4-B)。
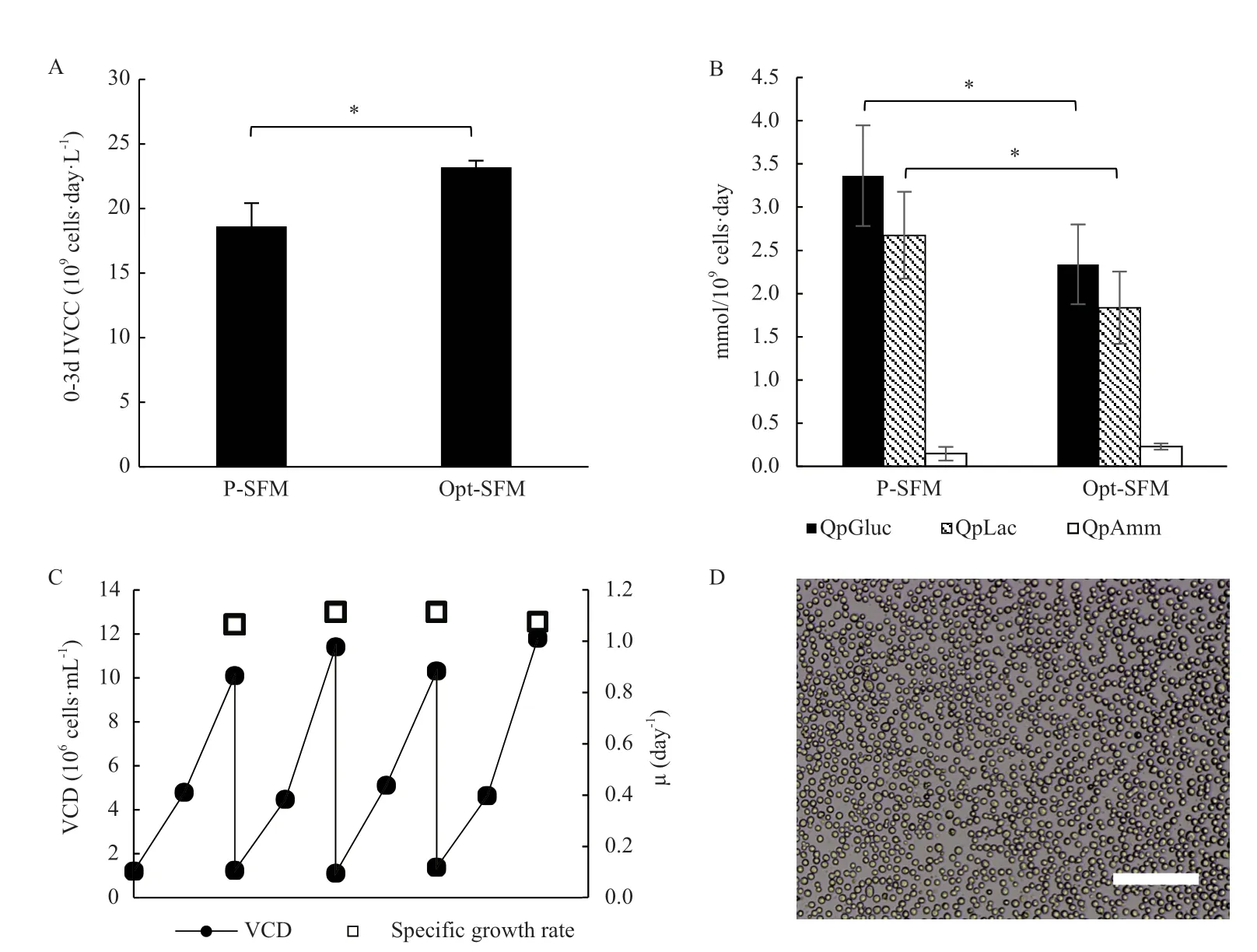
图3 基于摇瓶体系,BHK-21细胞在Opt-SFM中的生长和代谢
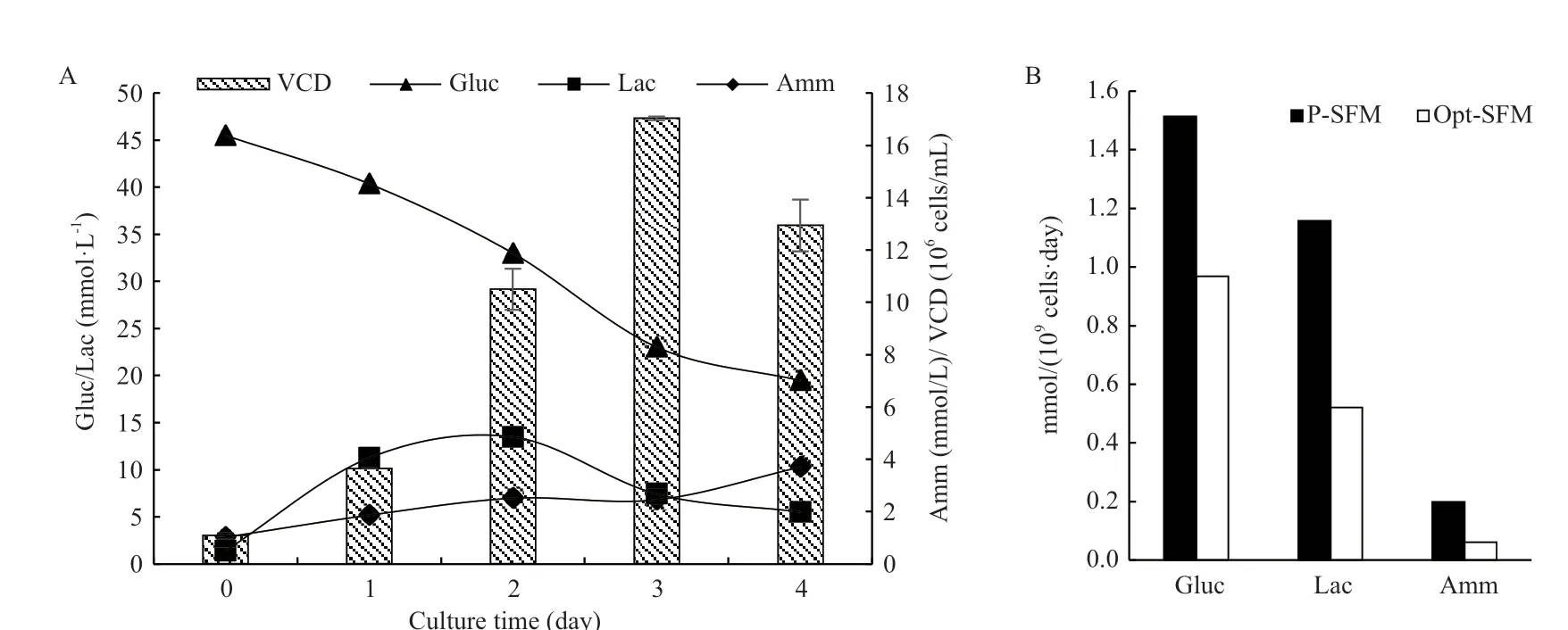
图4 基于反应器体系,BHK-21细胞在Opt-SFM中的生长和代谢
2.4 口蹄疫病毒在Opt-SFM中的扩增
2.4.1 摇瓶体系中口蹄疫病毒的扩增情况 以低血清培养基LSM作为对照,在Opt-SFM中,以任何接毒条件获得的病毒完整颗粒146s产量和单细胞产毒能力都高于对照组。如图5所示,病毒产量随接毒时细胞密度升高而升高,接毒细胞密度9×106cells/mL时,Opt-SFM中146s产量达到15.45 μg/mL。接毒操作方式上,最优选择是接毒前离心全量换液,流加补液其次,直接接毒所得病毒产量最低;在Opt-SFM中,接毒前离心换液的接毒操作最终得到了14.39 μg/mL的146s病毒产量,单细胞产毒能力达到13.95×104virions/mL。
图5 细胞密度和接毒方式对病毒产量和单细胞产毒能力的影响
2.4.2 反应器体系中口蹄疫病毒的扩增情况 反应器体系中,接毒前BHK-21细胞的密度达到10.1×106cells/mL,比生长速率为1.14/day,优于摇瓶体系,说明细胞不但达到了非常高的密度,而且仍处在活跃的指数生长期内。接毒0 h细胞形态饱满、均匀分散(图6-B)。接毒12 h后细胞病变率达到90%以上(图6-A),计细胞数时视野中出现大量死细胞和细胞碎片(图6-C)。
如图7,在Opt-SFM中,细胞病变90%以上收毒后测得毒液的效价为7.25 lgTCID50/0.1mL,与LSM实验组水平相似,但是在Opt-SFM中获得的146s含量为15.6 μg/mL,比LSM组提高了50.7%。
3 讨论

图6 BHK-21细胞在Opt-SFM中接种口蹄疫病毒细胞生长与病变情况
口蹄疫病毒属于烈性侵染病毒,一般24 h内使宿主细胞完全病变。作为病毒生产的“工厂”,接毒时BHK-21细胞的密度和状态都与病毒的最终产量有重要的关系。Maranga等[31]利用昆虫细胞杆状病毒表达系统在无血清培养基中生产猪细小病毒样颗粒(VLPs)时发现,选择合适的接毒时间,确保细胞处于指数生长期,比选择细胞密度高但处于稳定期的细胞更容易获得较高的产量。本研究中BHK-21细胞在Opt-SFM中培养48 h时细胞密度虽已达到107cells/mL,但仍处于指数生长期,此时细胞状态健康,细胞内各种酶的活性高,细胞产毒能力高,是理想的接毒时间。

图7 培养基优化前后病毒效价TCID50和146s完整病毒颗粒的含量
破除“细胞密度效应”的关键在于营养成分的供应和有毒代谢副产物的消除[11-12]。Dill等[15]的实验发现,培养体系中增加葡萄糖和谷氨酰胺的浓度时均不会影响病毒效价。然而,其文章中考察的葡萄糖浓度最低为22 mmol/L,仅支持最高3×106cells/mL的细胞密度,按照本文的结论,支持1.7×107cells/mL的BHK-21细胞密度仅需要45 mmol/L葡萄糖,因此,Dill等[15]的葡萄糖添加量已经超过了细胞对葡萄糖消耗的需求,成为葡萄糖浓度持续增加无法引起产量进一步提高,反而会导致乳酸大量积累的原因。进一步证明了按照细胞的代谢规律和细胞需求进行培养基设计和优化的重要性。本研究对无血清培养基中葡萄糖、3种氨基酸和铜锌离子浓度的设计均以营养成分的代谢规律为基础,满足细胞需求的同时,降低细胞的生长压力,因此代谢副产物的积累情况不升反降。降低乳酸和氨本身对病毒生产过程影响的同时[16-17],也降低了因乳酸积累可能引起的pH值降低和渗透压升高等环境条件的改变对产毒过程的负面影响[18-19]。
提高接毒时细胞密度和改善培养环境的另一个重要意义在于优化接毒操作。摇瓶体系筛选出的接毒前离心换液操作,在生物反应器水平实施时,通常采用低温沉降的方式,与多数企业现行的操作相同,目的是减小培养体系积累的代谢副产物或其他不利因素对产毒过程的干扰[21]。但是,低温沉降耗时耗力,而且不可避免地会影响细胞状态,进而降低其产毒能力。作为可替代的接毒操作方式,接毒前流加补液一定程度上使细胞维持在稳定状态,而且减少了换液操作可能引入的染菌风险。本文中,BHK-21细胞在Opt-SFM中接毒前流加补液的操作所获得的病毒产量高于在LSM中实施换液后接毒,这一结果为改良接毒工艺提供了可能性。
采用不含血清和蛋白、化学成分明确的培养基进行BHK-21细胞悬浮培养和口蹄疫病毒疫苗生产,能够在提高产能效率的同时降低血清等动物源性添加物的安全风险,并减少生产过程中杂蛋白对病毒疫苗纯化制剂的压力[32]。因此,使用无血清悬浮培养工艺生产病毒疫苗是生物制品产业发展的必然趋势,本研究的成果对于无血清培养基的工业化应用具有极强的现实意义。
4 结论
本研究开发的无血清培养基能够支持BHK-21悬浮细胞高密度培养和稳定传代,最高活细胞密度可达1.78×107cells/mL,培养过程中,乳酸和氨等代谢副产物都维持在较低水平。以1×106cells/mL的接种密度,长至48 h时接毒,相同操作条件下,无血清培养基可获得与低血清培养基相似的病毒效价(7.25 lgTCID50/0.1mL),而完整病毒粒子146s则达到了15.6 μg/mL,是低血清培养体系的1.5倍。无血清培养基具有完全替代低血清悬浮培养工艺生产口蹄疫病毒的潜能。
猜你喜欢
今日畜牧兽医(2022年10期)2022-12-23
今日农业(2021年8期)2021-11-28
中华养生保健(2020年8期)2021-01-14
中国甜菜糖业(2020年1期)2020-12-07
江苏卫生保健(2018年7期)2018-07-31
中国乳业(2016年4期)2016-11-07
小学生导刊(低年级)(2016年7期)2016-07-29
腹腔镜外科杂志(2016年12期)2016-06-01
恋爱婚姻家庭·养生版(2016年2期)2016-02-17
肿瘤影像学(2015年3期)2015-12-09
