
滇西北4种野生观赏植物的细胞学研究
2020-11-02 12:06陈光富
中国野生植物资源 2020年10期
陈光富,徐 波
(1.丽江师范高等专科学校应用技术学院,云南 丽江,674199;2.西南林业大学林学院,云南 昆明,650224)
滇西北地处青藏高原至云贵高原的过渡带上,包括云南省迪庆州、怒江州、丽江市、大理州和保山市等18个县(市、区),面积达79800 km2[1-2]。该地区具有典型的高山峡谷垂直景观、高山湿地生态系统和温性、寒温性针叶林生态系统等,植物资源丰富,有种子植物6559种,约占云南地区种子植物的41%,全国种子植物的20%[3],被称为中国植物多样性最丰富的地区之一[2,4],同时也是自然灾害频发、水土流失严重的生态脆弱区[5]。因此,该地区是植物区系组成、物种多样性保护、植物资源开发利用等研究的热点区域,目前研究主要集中于资源调查[2]、植物区系多样性[6-7]、部分类群核型报道[8]等方面,居群水平的细胞学研究却鲜见报道。
细胞学研究可以为植物区系起源、系统进化、物种多样性分析提供新资料[9-10],通过居群水平的遗传和变异研究,为亲缘关系、育种、种质资源鉴定等方面提供一定依据[11-13]。鉴于上述原因,本研究选取报春花科(Primulaceae)报春花属(PrimulaL.)偏花报春(P.secundifloraFranch)和灰岩皱叶报春(P.forrestiiBalf. f.)、桔梗科(Campanulaceae)蓝钟花属(CyananthusWall. ex Benth.)美丽蓝钟花(C.formosusDiels.)、百合科(Liliaceae)葱属(AlliumL.)多星韭(A.wallichiiKunth)等4种滇西北著名高山野生花卉资源为研究材料,开展细胞学研究。为滇西北地区4种植物居群间的差异分化、种质资源挖掘利用、生态多样性保护等方面提供基础证据,同时也补充完善该地区植物的细胞学资料。
1 材料和方法
1.1 材料
供试材料均采自滇西北地区(表1)。凭证标本保存于中国科学院昆明植物研究所标本馆(KUN)和丽江师范高等专科学校,永久封片保存于云南师范大学生命科学学院植物室。

表1 材料采集信息Table 1 Samples and their locality information
1.2 方法
采用植物根尖常规压片观察[14]、KaryoType 2.0软件测量[15]、李懋学和陈瑞阳核型分析标准[16]及核型不对称性系数AI值计算法开展研究工作[17]。试验流程主要包括种子萌发、取材料、预处理、固定、解离、染色、镜检和综合分析等8个环节。①种子萌发:将种子放置于培养基上,置于23~26℃恒温箱内进行萌发培养;②取材料:定期观察种子,并在种子根尖长至1.2~1.8 cm时取下材料;③预处理:用0.002 mol/ L 的8-羟基喹啉处理液常温下避光预处理1.5~3.5 h;④固定:将材料放置于4℃冰箱中并用卡诺固定液固定处理4~8 h;⑤解离:用1 mol/L盐酸在60℃水浴锅中解离6~20 min;⑥染色:卡宝品红染色液染色12~24 h;⑦镜检:制作临时装片,显微镜下观察并拍照记录;⑧综合分析:选取5个以上中期染色体分散较好的细胞测量并运用SPSS Statistics 22.0软件包进行统计分析。
通过查阅植物染色体信息相关数据库Tropicos-Index to Plant Chromosome Numbers和Chromosome Counts Database[18-19],选取核型参数相对完整的居群进行对比分析,从细胞染色体水平上对滇西北4种植物不同居群间的差异分化进行讨论。
2 结果和分析
2.1 偏花报春P. secundiflora Franch
该试验材料采自香格里拉市中乡公路边的云冷杉林下,体细胞染色体数目为22条,中期染色体全由中部着丝粒染色体组成,核型公式为2n=2x=22=22m;核型不对称性划分类型为Stebbins's-1B型。染色体长度值在1.91~4.74 μm之间变化;染色体平均长度值为3.21 μm;着丝粒指数CI值为41.88,不对称系数AI值为2.91。核型特征及指标参数详见表2和表3,植物形态图见图1中A1,体细胞分裂间期、前期、中期染色体形态图和核型模式图见图1中A2~A5。

表2 4种植物的染色体参数Table 2 Chromosome parameters of four species
2.2 灰岩皱叶报春P. forrestii Balf. f.
该试验材料采自丽江市玉龙雪山黑水河的河边云冷杉林下,体细胞染色体数目为24条,中期染色体由11条中部着丝粒染色体和1条近中部着丝粒染色体组成,核型公式为2n=2x=24=22m+2sm;核型不对称性划分类型为Stebbins′s-1B型。染色体长度值在4.96~11.62 μm之间变化;染色体平均长度值为8.08 μm;着丝粒指数CI值为41.69,不对称系数AI值为3.99。核型特征及指标参数详见表2和表3,植物形态图见图1中B1,体细胞分裂间期、前期、中期染色体形态图和核型模式图见图1中B2~B5。
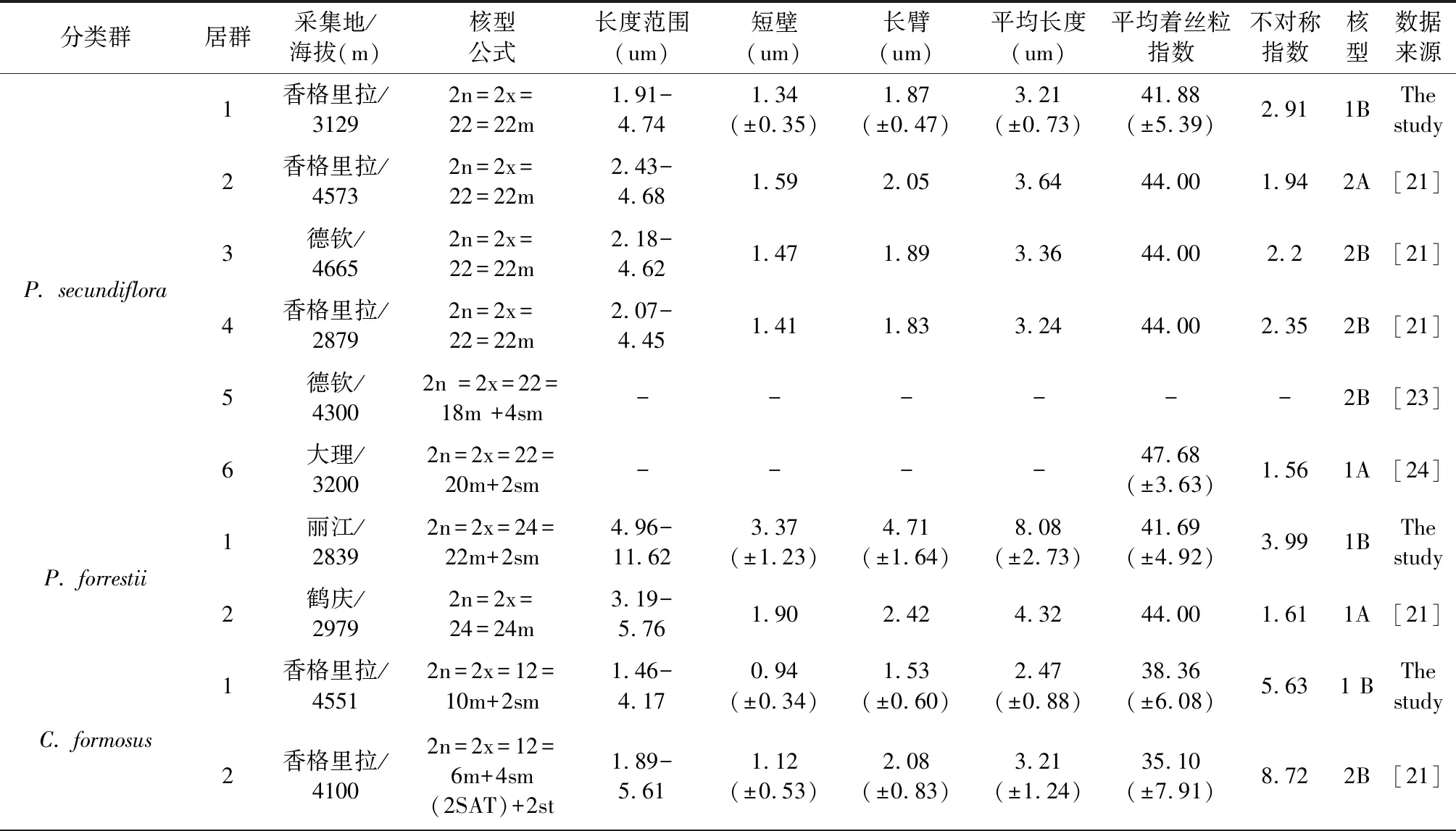
表3 滇西北4种植物不同居群的核型参数Table 3 Cytolotical characteristics of different populations of four species in Northwest Yunnan

表3 滇西北4种植物不同居群的核型参数Table 3 Cytolotical characteristics of different populations of four species in Northwest Yunnan
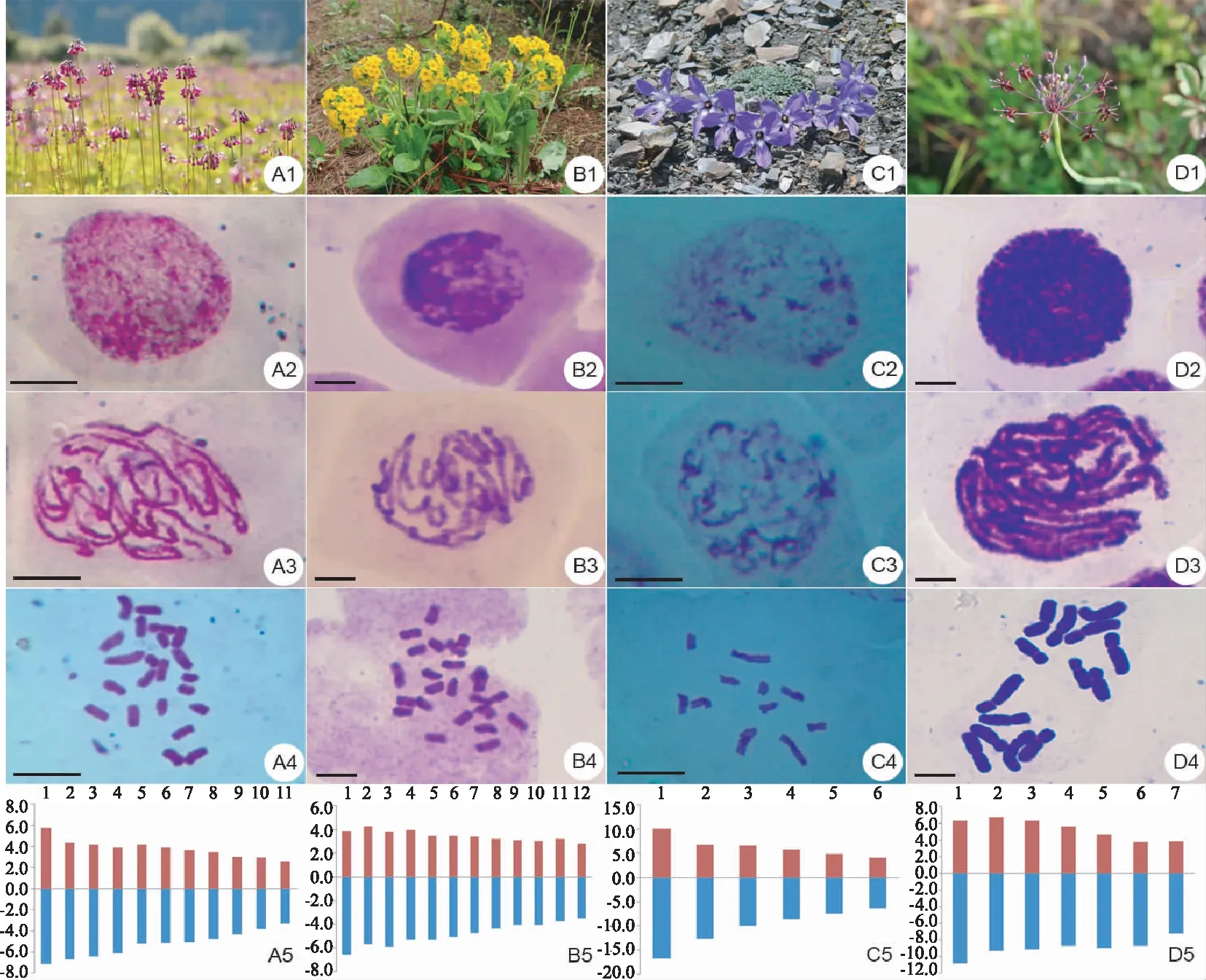
图1 4种植物的形态、间期核、有丝分裂前期和中期染色体及核型模式图Fig. 1 Morphology and cytological features four species注:A1-A5:偏花报春;B1-B5:灰岩皱叶报春;C1-C5:美丽蓝钟花;D1-D5:多星韭。标尺=5 m
2.3 美丽蓝钟花C. formosus Diels.
该试验材料采自香格里拉市格咱乡翁水的高山流石滩,体细胞染色体数目为12条,中期染色体由5条中部着丝粒染色体和1条近中部着丝粒染色体组成,核型公式为2n=2x=12=10m+2sm;核型不对称性划分类型为Stebbins’s-1B型。染色体长度值在1.46~4.17 μm之间变化;染色体平均长度值为2.47 μm;着丝粒指数CI值为38.36,不对称系数AI值为5.63。核型特征及指标参数详见表2和表3,植物形态图见图1中C1,体细胞分裂间期、前期、中期染色体形态图和核型模式图见图1中C2~C5。
2.4 多星韭A. wallichii Kunth
该试验材料采自丽江市玉龙雪山高山植物园的云冷杉林下,体细胞染色体数目为14条,中期染色体由3条中部着丝粒染色体和4条近中部着丝粒染色体组成,核型公式为2n=2x=14=6m+8sm;核型不对称性划分类型为Stebbins’s-2A型。染色体长度值在6.276~10.16 μm之间变化;染色体平均长度值为8.25 μm;着丝粒指数CI值为36.68,不对称系数AI值为3.12。核型特征及指标参数详见表2和表3,植物形态图见图1中D1,体细胞分裂间期、前期、中期染色体形态图和核型模式图见图1中D2~D5。
3 讨论
报春花属约有500种,研究表明沿喜马拉雅山两侧和云南、四川西部是报春花属的现代分布中心,该属植物花多艳丽,是世界著名的高山花卉,因此主要用于栽培观赏[1,20]。据统计,已有185种报春花属植物有染色体数目的相关报道,可以确定染色体数目范围为16~198条;染色体基数包括x=8,9,10,11,12等,其中以x=9 和 11最为普遍;倍性水平为2n=2x,4x,6x,8x,14x等,其中以二倍体较为普遍,占比高达该属已报道种数的59.10%[21]。在我们的研究中,偏花报春染色体数目为2n=22,灰岩皱叶报春染色体数目为2n=24,染色体数目在前人研究结果范围内;从而可以确定,这两种观赏植物的染色体基数分别为x=11和12,均为二倍体;研究结果也补充支持报春花属倍性进化水平以二倍体为主的观点。其次,通过统计偏花报春和灰岩皱叶报春两种植物在滇西北地区不同海拔梯度下分布居群的染色体核型数据[22-24],发现在染色体数目、倍性水平、染色体类型、核型不对称系数等核型特征上变化稳定,核型类型在1A,1B,2A,2B之间变化。以上两种植物在滇西北地区种内核型特征变化差异较小的现象,可能是植物在局部地区对环境长期适应中(2700~4665 m),已经形成稳定的细胞学特征。
蓝钟花属植物分布于喜马拉雅山及邻近地区,为中国—喜马拉雅特有属。在中国主要分布于西藏、云南(滇中、滇西北)、四川(川西)、甘肃和青海等省区,云南西北部认为被是其分布和变异中心[25]。已有报道表明,该属染色体数目2n=10,12,14,24,28等,倍性包括2n=2x和4 x,染色体基数有x=5,6,7[8,26]。此次报道的美丽蓝钟花翁水居群染色体数目为2n=12,可确定为二倍体,基数为x=6。以上结果与前人报道的香格里拉市石卡山居群相一致[26],但两居群在核型类型上有一定的差异(1B和2B型);同时染色体组成类型也有较大变化,石卡山居群有m、sm和st型,并出现了随体,而翁水居群仅有m和sm型,也未观察到随体出现。依据《植物志》记载[25],美丽蓝钟花在中国主要分布于云南西北部和四川西南部,生于海拔2800~4100 m的山地草坡、林间沙地和林边碎石地上;而石卡山居群和翁水居群的实验材料均采自海拔4100~4551 m的高山边缘带(流石滩),高山边缘带是陆地上海拔最高的植被带,生境具有氧气稀薄、昼夜温差大、强风、强日照辐射等特征,生存环境较为恶劣,只有具备特殊生物学特性的物种才能定居于此[27]。以上两居群的核型特征差异似乎也表明,植物染色体结构变化、核型特征分化可能是植物为适应环境而发生的改变。
多星韭是多年生草本,主要分布于东喜马拉雅地区及云贵高原,生于海拔2300~4800 m的草坡、林缘和灌丛下[28],在滇西北地区当地居民主要用于食用、药用和观赏。已报道的染色体资料表明[29-34],多星韭染色体数目2n=14,28,基数x=7,较为稳定,倍性2n=2x,4x。我们的研究结果与以上报道一致,可以确定玉龙雪山高山植物园居群2n=2x=14,为二倍体。统计的滇西北地区多星韭21个不同居群中,核型类型为2A型的居群均在2077~3425 m海拔梯度范围内分布,核型类型为3A型的居群仅在海拔为3638 m的香格里拉出现;而在核型组成上存在较大分化,主要表现为染色体类型组成多样(属m型、sm型和st型),随体和B染色体出现的不规律性,不对称系数AI值变化(在1.58~3.45之间变化),倍性变化(二倍体单生、四倍体单生、二倍体和四倍体混生等3种类型)等,可以推测滇西北地区分布的多星韭在居群水平上分化较为剧烈。在海拔梯度与倍性水平变化方面,二倍体居群的海拔分布梯度为2100~3638 m,四倍体居群的海拔分布梯度为2077~3245 m,二倍体的生态分布幅度更大;结合AI值变化,二倍体居群为1.58~3.45,四倍体居群为1.61~2.79,依据Stebbins“核型对称—原始,核型不对称—更进化”的原理[35],二倍体不对称系数变幅大于四倍体的不对称系数(1.58<1.61<2.79<3.4),以上结果支持前人[30]提出的多星韭二倍体可能来自同一个祖先,在从高海拔向低海拔方向发展的过程中,为了适应新环境,二倍体开始出现分化,新的四倍体也逐渐形成的推论。其次,随体出现的居群分布海拔范围为2077~3638 m,无随体的居群分布海拔范围为2470~3425 m,有学者认为随体的出现是染色体发生结构变异的形态表现,其数目可能与植物所处的海拔有关[36],这似乎也说明随体的出现使植物适应更广阔的生态分布。当然,要弄清楚这一热点地区中分布海拔与随体、倍性变化、B随体等核型特征的相关性,需要对多居群、多类群系统的细胞学研究。
致谢中国科学院昆明植物研究所高山植物多样性研究组提供实验种子,刘德团先生、段芳芳女士提供帮助,在此一并致谢。
猜你喜欢
植物研究(2022年5期)2022-10-12
河北科技师范学院学报(2022年2期)2022-08-26
江西农业学报(2022年5期)2022-08-10
北极光(2022年1期)2022-02-23
浙江中医药大学学报(2021年6期)2021-07-12
军工文化(2021年2期)2021-03-30
幽默大师(漫话国学)(2020年9期)2020-09-15
江苏农业科学(2017年16期)2017-10-27
现代农业科技(2017年8期)2017-06-10
航空模型(2017年2期)2017-05-22
