
土壤可溶性有机氮研究进展
2020-10-31 08:08陈仕友王祎白彦锋姜春前
生态科学 2020年5期
陈仕友,王祎,白彦锋,姜春前,
1. 中国林业科学研究院林业研究所国家林业和草原局林木培育重点实验室,北京100091
2. 福建农林大学资源与环境学院,福州 350002
0 前言
氮是地球上所有生命体不可替代的必须生命元素,也是农业生产的三要素之首,还是农业增产的关键要素[1]。 土壤中的氮素绝大部分以有机态存在[2],而土壤可溶性有机氮是土壤有机氮的重要组成部分[3],也是土壤氮库最活跃的组分之一[4]。土壤可溶性有机氮(Soluble Organic Nitrogen,SON)是指在土壤中能被水或盐溶液(如 KCl、K2SO4、CaCl2等)浸提出来的有机态氮,或用电超滤法(Electroultrafiltration,EUF)提取的土壤有机氮素[5-6]。土壤可溶性有机氮可作为土壤微生物的氮源和碳源[7],小分子的氨基酸类物质也可以直接被植物吸收利用[8]。有报道表明有些植物吸收有机氮的含量超过了该植物吸收无机态氮(NO3-,NH4+)的含量,如野外收集的北极珊瑚藻(Eriophorum vaginatum)根系迅速吸收游离氨基酸,至少占该物种吸收氮的60%[9]。菌根真菌可以吸收可溶性有机氮之后向宿主植物转运[10]。可见,土壤可溶性有机氮在氮循环过程中扮演着十分重要的角色[11]。但是土壤可溶性有机氮可以被水溶解进入土壤溶液成为土壤溶解态有机氮(Dissolved Organic Nitrogen,DON),由于土壤胶体带负电,溶解态有机氮很容易随土壤水分一同向下运动[12]、淋失,造成农业面源污染等环境问题[13],所以土壤可溶性有机氮研究越来越受到重视。
本文通过土壤可溶性有机氮的含量、土壤可溶性有机氮的来源和成分、土壤可溶性有机氮的迁移特征以及土壤可溶性有机氮的影响因素几个方面展开论述,最后提出土壤可溶性有机氮(SON)未来主要的研究方向。
1 土壤可溶性有机氮的来源和成分
土壤可溶性有机氮的来源主要是大分子的颗粒态有机氮在由土壤中微生物主导的解聚作用分解形成[14],而这些解聚作用往往是在胞外解聚酶的作用下发生氨化过程的第一步[15]。生物残体(包括植物、动物和微生物)或者施用有机肥会带入大量大分子的有机氮。其中最主要的大分子有机氮为蛋白质、几丁质和肽聚糖,蛋白质在细胞外蛋白酶和肽酶的作用下将蛋白质和多肽水解成肽和氨基酸,几丁质在几丁质酶的作用下分解成 N-乙酰氨基葡萄糖,肽聚糖在不同的酰胺酶,肽酶和糖基化酶的作用下分解为氨基糖和氨基酸[7]。

图1 微生物利用可溶性有氮途径[7]Figure 1 Main steps of the microbial N utilization from organic substrates[7]
土壤可溶性有机氮同土壤可溶性有机物的来源相同,主要为新近凋落物和土壤腐殖质,而土壤微生物、有机质(包括植物残体和动物残体)的分解、土壤胶体的解吸、根系代谢产物及分泌物、土壤微生物发生腐殖化作用、施入的有机肥、降雨淋溶及大气沉降等都是土壤可溶性有机质的重要来源[15]。
土壤可溶性有机氮组成目前国内外有关研究报道较少,主要可能是土壤可溶性有机氮成分复杂且在土壤中含量较低,利用普通的测试手段难以达到测试需求,且土壤有机氮的组成也会随着生态系统的结构、群落的演替、土壤的发育过程,土壤类型、土地利用的不同和人为干扰等因素产生差异。Murphy等[16]认为土壤中可溶性有机氮的组成非常复杂,从其生态功能看可溶性有机氮的组分主要包括氨基酸、氨基糖、蛋白质等小分子含氮化合物。(Michalzik等[17]人认为土壤可溶性有机氮的重要组成部分是土壤游离氨基酸和蛋白质。Currie等[18]研究发现半分解状态的植物、微生物、动物残体形成的难以降解的有机酸是凋落物土壤可溶性有机氮的主要成分。Jones等[19]研究表明,三种草地土壤可溶性有机氮的各组分中,大部分的土壤可溶性有机氮组分都是难降解的含氮有机物,而易降解的低分子量含氮有机物仅占总土壤可溶性有机氮的 3%—15%,葛体达等[20]人测量的不同园艺生产系统中游离氨基酸占土壤可溶性有机氮的 2.77%—10.93%,占土壤可溶性总氮的1.19%—2.01%。但是,也有研究发现[21]耕作土壤中游离氨基酸和氨基糖含量低,仅占土壤可溶性有机氮的 5%以下,杂环氮化合物占土壤可溶性有机氮的比例为 15%,而肽和蛋白质等氨氮化合物占土壤可溶性有机氮的比例则为35%—57%。Paul等[22]人的研究也有相似结论,耕地中主要成分是蛋白质及多肽,占土壤可溶性有机氮的 35%—57%,游离氨基酸占土壤可溶性有机氮的3%,氨基糖占 15%,杂环化合物占 15%;而森林土壤中自由氨基酸占土壤可溶性有机氮总量的 1.5%—25%。
综上所述,对于土壤可溶性有机氮的组分未形成一致的科学认识,今后还需要深入开展不同生态系统、不同土壤类型中土壤可溶性有机氮组分研究工作。
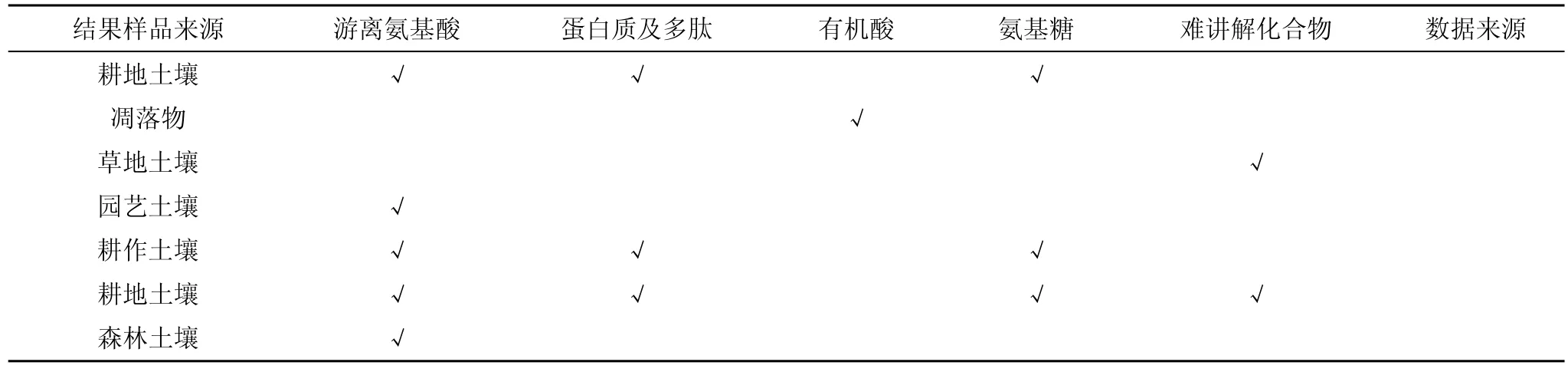
表1 土壤可溶性有机氮的主要组分Table 1 Main component of soluble organic nitrogen
2 土壤可溶性有机氮的含量
现在的测试手段还无法直接测量土壤可溶性有机氮的含量,主要运用差减法,即土壤可溶性总氮减去土壤可溶性无机氮(NO3-,NO2-,NH4+)来计算获得土壤可溶性有机氮含量[23],不同生态系统的土壤可溶性有机氮含量不同。
森林生态系统的可溶性有机氮含量研究较深入。Chen等[11]收集调查了澳大利亚22个亚热带林地表层土壤(0—10 cm),测得可溶性氮(2 mol KCl浸提)含量为 5—45 mg·kg-1,占土壤可溶性总氮的39%,占土壤全氮含量的 2.3%。江玉梅等[24]调查了四个亚热带不同林分的表层退化红壤表明土壤可溶性有机氮(热水浸提)含量为 24—73 mg·kg—1。郭新春[25]发现亚热带森林生态系统表层土壤可溶性有机氮最高含量可占总可溶性总氮的63%—89%。Zhong和 Makeschin[5]对德国的温带森林调查发现,在表层0—10 cm的土壤可溶性有机氮(0.5 M K2SO4浸提)的含量为 6.6—10.2 mg·kg-1,占全氮的比例为0.3%—2.2%,并发现在矿质土壤中氯化钾(1 M KCl)和硫酸钾(0.5 M K2SO4)的浸提效果相似,但在有机土壤层的氯化钾对土壤可溶性有机氮的提取量约为硫酸钾提取量的1.2—1.4倍。杨绒等[26]发现温带林地土壤枯枝落叶层中土壤可溶性有机氮含量为 245.11—254.32 mg·kg-1,平均 248.26 mg·kg-1,占土壤可溶性总氮的 74.07%—79.91%,占土壤全氮的3.84%—4.73%,林地0—20 cm土层土壤可溶性有机氮含量为 21.80—38.64 mg·kg-1,平均为31.03 mg·kg-1,占土壤可溶性总氮的 37.60%—50.68%,占土壤全氮的1.30%—2.69%。庄舜尧等[27]发现热带高山地区有机层(O层)的土壤可溶性氮在310.1—462.35 mg·kg-1,占全氮的比例为 2.51%—5.43%,腐殖质层(A 层)土壤可溶性有机氮含量为17.6—44.8 mg·kg-1,占全氮的比例为 0.34%—0.54%。退耕还林可显著增加土壤硝态氮、铵态氮和可溶性有机氮含量,尤其是0—10 cm 土层增加明显。0—10,10—20,20—30 cm 土层间差异显著,沙棘林和山杏林间也达到显著差异水平。总体来看,硝态氮含量随季节变化明显,3月最高,3—6 月下降,6—9 月上升,9—11 月又下降,11 月最低。各样地间0—30 cm 土层铵态氮含量变化幅度较小。可溶性有机氮含量随季节变化明显,6月最高,11 月最低,3—6 月上升,6—9 月下降,9—11 月又上升。可溶性有机氮含量季节变化规律与硝态氮变化规律相反。沙棘林地土壤可溶性氮含量较杏树林地高,且阳坡高于阴坡。0—30 cm 土层可溶性氮组分以可溶性有机氮为主,占 73.3%—99.0%,其次是硝态氮,占 0.9%—24.3%;铵态氮所占比例最少,在2%以下。退耕还林措施可显著增加0—10 cm 土层土壤可溶性氮含量,不同林分土壤可溶性氮含量增加不同,沙棘林地高于杏树林地[28]。
在农田和草原生态系统中,Ghani等[29]研究了新西兰牧区93个地点的表层土壤(0—7.5 cm)发现土壤可溶性有机氮的含量为13—93 mg·kg-1,土壤可溶性氮的含量占土壤可溶性总氮的 16.7%—89.3%。柯英等[30]调查发现宁夏设施菜田0—150 cm土壤剖面溶解性总氮、硝态氮和可溶性有机氮含量都显著高于水旱轮作大田,前者分别是后者的1.5—5.6、1.5—3.4倍和 1.6—9.8倍,并且设施菜田土壤氮素主要累积在0—5 cm和5—20 cm土层,而水旱轮作大田主要积累在4—100 cm土体。Murphy等[31]对12个英国农田土壤的测定发现,KCl提取的土壤可溶性有机氮含量在22—34 kg·hm-2之间,约占土壤可溶性氮总量的 40%—50%。葛体达等[20]发现有机蔬果园可溶性有机氮的含量为 5.4—16.6 mg·kg-1,占土壤可溶性总氮的 18.4%—42.9%,占全氮的7.4%—11.9%。
在农业生产中,农民常采用施用有机肥的方式提高地力,保证农业生态系统的可持续利用。有机肥中通常含有一定量的可溶性有机氮,并且里面含有大量的蛋白质等多聚物,可以发生解聚作用形成土壤可溶性有机氮。研究表明长期施用有机物料可以显著地提高土壤可溶性有机氮的含量[32],且随着有机物料投入量的增加,土壤可溶性有机氮呈增加的趋势[33]。高C/N比的如小麦秸秆和黑麦草回田对农田土壤可溶性有机氮含量影响不明显,而施用低 C/N比的牧草如苜蓿后土壤可溶性有机氮含量明显提高,并呈先增后降的变化规律[34]。
综上所述,在开展土壤可溶性有机氮相关方面的研究工作时,应从生态系统类型、土壤类型、样地土壤质地、土壤有机质含量等方面综合考虑。
3 土壤可溶性有机氮的迁移特征
DON 作为土壤有机氮矿化和无机氮固定过程中重要的中间氮库,调控着土壤铵态氮(NH+4-N) 的供应和氮素的生物转化过程,是土壤有效养分的来源之一[3-4]。氮素淋溶及随径流损失是氮素向周围水体迁移的主要途径。前期研究土壤淋失过程中通常以无机态氮为主,但是近年来研究发现 DON 淋失现象可能比无机态氮更严重[35]。土壤可溶性有机氮可以被土壤水分溶解进入土壤溶液,随着土壤水盐一起运动。Perakis等[36]研究发现南美温带原始森林生态系统中氮素主要以土壤可溶性有机氮的形式流失。Yang等[37]研究表明土壤可溶性有机氮是水田土壤氮素径流和下渗淋失的主要形态之一,分别占相应淋失总氮量的 21.6%—39.9%和 22.3%—41.9%,施用控释氮肥可降低水田土壤可溶性有机氮的淋失;施用化学氮肥显著增加温室蔬菜地土壤可溶性有机氮的数量,且土壤可溶性有机氮迁移与无机氮迁移是同步的,施氮水平对水田土壤渗滤液中土壤溶解态有机氮浓度具有极显著影响,土壤溶解态有机氮最高值可占渗滤液中土壤可溶性总氮的 50%[38];Song等[39]研究结果表明我国南方水稻—小麦轮作农田系统中水稻和小麦生长季土壤可溶性有机氮累积淋失量分别达 1.1—2.3 kg·hm-2和 0.01—1.3 kg hm-2,氮肥对土壤可溶性有机氮淋失的影响极显著。Zhao等[40]人在亚热带农田土壤上施用稻草生物炭、微生物肥料表明生物炭和微生物肥料可以减少土壤可溶性有机氮淋失;Long等[41]在亚热带酸性土壤上连续施用高量猪粪(600 kg·hm-2每年),3年后玉米地土壤可溶性有机氮淋失量显著提高,年均淋失量高达 3.8—5.4 kg·hm-2。Vinther等[42]研究表明砂质草地土壤翻压三叶草后砂壤土和粗砂土土壤可溶性有机氮年平均淋失量分别达 3—4 kg·hm-2和 10—31 kg·hm-2。土壤可溶性有机氮是西班牙东南部以农业为主集水区河水中氮素的主要形态,总量占河水总氮量的 72%—97%,且地下水中溶解态有机氮也占总氮量的 1%—99%[43];茶园 40 cm 土层内土壤可溶性有机氮下渗迁移主要发生在0—20 cm,其迁移通量与茶园植茶年限显著正相关[44]。耕作频率对土壤可溶性氮影响十分明显,土壤可溶性氮与随扰动频率增加而增加,连续性耕作会增加水体氮素污染风险[45]。土壤可溶性氮通常在红壤下层(40—60 cm)有积累的趋势[46]。土壤可溶性有机氮是英国 South Pennine泥炭地流域河流中土壤可溶性总氮的主要成分,土壤可溶性有机氮所占比例可高达土壤可溶性总氮的 54%[47]。总之,土壤可溶性有机氮的迁移特征尚有待进一步研究。
4 土壤可溶性有机氮的影响因素
土壤可溶性有机氮的产生是在大部分的酶促反应进行的,所以影响微生物活动的和影响酶活性的因素均会影响土壤可溶性有机氮的含量。影响土壤可溶性有机氮含量变化的因素较为复杂,产生的土壤可溶性有机氮一部分会经过氨化作用、硝化作用转变为铵态氮和硝态氮直接被植物吸收利用,一部分被土壤胶体或者粘粒吸附保护,随土壤水分向下迁移运动,一部分会被微生物进行腐殖化作用生成大分子的颗粒态有机氮。
Murphy等[3]通过线性相关分析表明好气培养试验中土壤可溶性有机氮与氮素净矿化率和无机氮总量均有显著的相关性。田飞飞等[48]人在大沽河流域选取农田土施肥培养实验,设置三个不同的温度(15℃、25 ℃、35 ℃),三个不同的水分梯度(60%、75%、90%)培养84天,结果表明土壤的矿化率随温度的增加而增加,但水分对各处理土壤的 SON 无显著差异。石思博等[49]人稻田定位观测实验结果表明,菌渣化肥配施能够显著提高土壤可溶性碳氮,但不随着施用量的增加持续呈增加态势,土壤可溶性碳氮比下降。施用有机物料后,不同土层氮矿化率也不一样。有机物料的种类不同,所含的C/N比不一样,矿化过程中土壤矿质氮和土壤可溶性有机氮的释放也各不相同。安小娟[50]利用川西北高寒草甸土频度施氮实验进行研究也表明不同频度(低频度、高频度)处理都对植物整个生长期内的土壤 DOC 含量呈上升趋势,高寒草甸生态系统表现出 DOC 的积累,而土壤 DON 的含量与温度变化一致。研究表明[17]土壤中土壤可溶性有机氮的含量与总氮(TN)和总可溶性氮(TSN)间呈现出极显著的相关性。葛体达等[22]也指出土壤可溶性有机氮与全氮、全碳、NO3--N、NH4+-N、总可溶性氮、游离氨基酸之间均有极显著的相关性。pH通常被认为是农业土壤中影响有机质周转的重要因素,通过影响微生物的群落和数量进而影响有机物质的分解。Kemmitt等[51]研究表明土壤可溶性有机氮含量与 pH之间存在一定的相关关系,施用石灰能够显著地增加洛桑试验土壤中土壤可溶性有机氮的含量。杨绒[50]研究表明添加猪粪处理土壤可溶性有机氮与微生物量碳(MBC)、微生物量氮(MBN)和NO3--N间存在显著正相关。陈安强等研究也得出了相似的结论,土壤中微生物量碳、微生物量氮与可溶性有机碳、可溶性有机氮之间呈现极显著的正相关,这说明了可溶性有机碳与可溶性有机氮既是微生物生长繁殖的能量源,又是微生物死亡后的消纳汇,它们不断处于动态转化过程中[52]。Xing等[53]对亚热带森林生态系统的研究表明除了可溶性有机碳(SOC)、总碳(TC)、土壤总氮、土壤微生物的影响外,脲酶、蛋白酶、天冬酰胺酶与 SON含量也高度相关。MBC、DOC 与SOC,MBN 与TN均呈极显著正相关,DON 和 TN 呈显著正相关,MBC 和 MBN、DOC 和 DON、DOC+MBC 和DON+MBN 之间均呈极显著正相关[49]。在自然水体中,N 主要以DIN、DON 以及颗粒性有机N(DPN)的形式存在,赤潮异弯藻对 DON 具有较强的利用能力[54]。
土壤微生物是土壤中物质转化和能量代谢的动力。随着分子生物技术的发展,国内外学者利用高通量测序技术(High Throughput Sequencing)、宏转录组(Metatranscriptome)、基因芯片(Geo chip)技术等现代生物技术在氮循环微生物生态功能研究上取得了重要的进展,认为氮代谢途径与环境类型、土壤碳、氮含量相关,高分子有机氮的氨化作用是土壤可溶性有机氮形成的重要过程。氨化过程由微生物功能基因控制,包括相关胞外酶基因和胞内脱氨酶基因等。研究表明,氨化过程的关键限速步骤正是这些天然有机氮大分子[55]。最初参与氨化过程的天然有机氮大分子,其在微生物细胞外物理、化学、生物作用(胞外酶)下生成低分子可溶性有机氮。土壤可溶性有机氮通常会经过氨化作用、硝化作用转变为铵态氮和硝态氮,完成土壤氮素的转化迁移过程。内转录间隔序列和16s rRNA的高通量测序表明,生物肥料氨化剂改变了微生物群落的组成,可溶性有机氮浓度增加[56]。当可溶性有机氮后经过一系列的转氨作用形成蛋白,在微生物系统中不断被循环利用。关于土壤可溶性氮相关的功能微生群及其在土壤氮库的转化机制研究较少。随着生物化学和分子生物学研究方法不断发展,新技术和新方法将被应用于土壤可溶性有机氮的研究中特别是加强土壤可溶性有机氮在土壤氮素循环和供氮能力两个方面的微观响应研究。
5 展望
土壤氮素形态各异,十分复杂。土壤可溶性有机氮是大气、陆地、江河、湖泊以及海洋等生态系统中氮循环的一个重要组成部分。氮素各种形态间的转化过程处于同一个系统内。这些转化过程往往同时发生、相互影响、相互作用,共同决定氮形态及去向。因此,研究土壤可溶性有机氮对全球生态系统碳、氮循环具有重要意义。土壤可溶性有机氮在数量及其影响因素上研究的学者较多,但从分子水平、生态学角度上看,土壤可溶性有机氮的组分特征及其在氮循环过程的速率及机制尚不清楚。今后,有待进一步的研究工作有: (1)关于土壤可溶性有机氮在生态系统组分特征,大量学者集中于小分子氨基酸的组分,需要进一步从构件上确认土壤可溶性有机氮中复杂的大分子有机物的特征,如带什么类型的官能团,并深入土壤可溶性机氮和所处环境的相互影响机制研究(2)关于土壤微生物对土壤可溶性有机氮作用特征集中在土壤微生物多样性及生物量对其的影响,可以从分子生态学的角度,研究土壤可溶性有机氮关键功能基因及特征种群,定量和定性判断对参与土壤氮循环中的土壤可溶性微观机制。 (3)关于土壤可溶有机氮流失效应,我们应该从生态系统、小流域、景观、全球不同的尺度量化土壤可溶性有机氮在氮循环各个过程的速率,模拟土壤可溶性有机氮在未来长时间的动态变化动态,阐明土壤可溶性有机氮环境响应特征和氮动态与全球变化的相关关系。
猜你喜欢
今日农业(2022年14期)2022-09-15
今日农业(2020年17期)2020-10-27
当代陕西(2019年23期)2020-01-06
天然产物研究与开发(2019年1期)2019-03-01
中成药(2018年8期)2018-08-29
山东青年(2016年2期)2016-02-28
医学研究杂志(2015年9期)2015-07-01
中国当代医药(2015年9期)2015-03-01
植物营养与肥料学报(2011年5期)2011-11-06
植物营养与肥料学报(2011年2期)2011-10-26
