
亚热带常绿阔叶天然林凋落物动态对短期氮沉降的响应(简报)
2020-10-26 03:45:02陈勇刘雨晖刘春华陈辉沈宝贵蒋宗垲苏素霞李丽红陈德叶刘小飞
亚热带资源与环境学报 2020年3期
陈勇,刘雨晖,刘春华,陈辉,沈宝贵,蒋宗垲,苏素霞,李丽红,陈德叶,刘小飞
(1.福建农林大学 莘口教学林场,福建 三明 365002;2.福建师范大学 地理科学学院,福州 350007)
据估计,近100年来,全球氮沉降量增加了110%,是自然氮沉降量的1.3~1.5倍,而且氮沉降速度仍然在上升[1-2]。中国已成为全球氮沉降最严重区域之一,如中国热带和亚热带地区一些森林生态系统氮沉降量高达30~73 kg N·hm-2·a-1,已超过生态系统氮饱和临界值(25 kg N·hm-2),并引起了土壤酸化、土壤肥力衰退和生物多样性降低等一系列严重的生态环境问题[3-4]。因此,全球氮沉降的现状及未来发展趋势对森林生态系统结构与功能的影响已成为全球生态学家关注的焦点之一。
森林凋落物是森林生产力重要组成部分,在维持森林生态系统物质、能量和碳循环过程中起着至关重要的作用[5-6],但其对氮沉降的响应仍然存在较大的争议。有研究表明,氮沉降增加提高土壤氮有效性[7],促进植物的生长,提高植物的生产力,因而增加凋落物量[8-9];也有研究表明,过量的氮输入改变了植物的生理生态机制,引起土壤酸化,降低植物多样性和生产力[3-4],进而减少凋落物产量[10]。虽然这些结果为探讨森林生态系统对氮沉降的响应提供了有力证据,但仍存在许多不足,如凋落物量组分只分为叶、枝和杂物,不能充分反映森林凋落物的组成特征[8-10]。此外,大部分研究主要集中在次生林和人工林[9-10],而对占有林地面积的63.7%的天然林[11]相关研究鲜见报道。2019年7月,中共中央办公厅、国务院办公厅联合印发《天然林保护修复制度方案》,提出全面保护天然林,维护天然林的完整性和原始性,实现人与自然和谐发展 。因此,在未来全球变化背景下,开展氮沉降对常绿阔叶天然林凋落物产量和组分的影响研究,有助于深入理解亚热带森林生态系统碳循环过程应对氮沉降的响应及其机制。
选择亚热带地区地带性植被米槠(Castanopsiscarlesii)天然林作为研究对象,探讨模拟氮沉降对凋落物产量及组分月动态变化的影响,为全面理解氮沉降背景下亚热带地区森林生态系统的碳库和碳循环研究提供科学支撑。
1 材料与方法
1.1 研究地概况
研究地位于三明市格氏栲自然保护区内(26°07′N,117°24′E)。本区属中亚热带海洋性季风气候,观测期间(2013—2015年)年均降雨量大约为1 906 mm,降雨主要集中在 4—8 月;2013—2015年年均温 18.7 ℃,最冷月平均气温 4.5~9.2 ℃,最热月平均气温 22.3~28.5 ℃。样地海拔 315 m,坡度 35°。2011年调查林分密度为 1 955 株·hm-2,郁闭度 0.89,平均树高 11.9 m。土壤类型为砂岩发育的山地红壤,土壤容重为1.12 g·cm-3,土壤全碳、全氮、全磷和全钾含量分别为24.5、1.86、0.21和2.9 g·kg-1。
试验地群落植物层次结构明显,其中乔木主要有米槠、木荷(Schimasuperba)、马尾松(Pinusmassoniana)等;林下植物主要有狗骨柴(Tricalysiadubia)、山黄皮(Aidiacochinchinensis)、赤楠(Syzygiumbuxifolium),草本层主要有草珊瑚(Sarcandraglabra)、山姜(Alpiniajaponica)、网脉酸藤子(Embeliarudis)等。
1.2 试验设计
2011年6月,在米槠天然林内设置12个20 m×20 m样地,相邻两个样地之间间隔10 m作为缓冲区。试验设置对照(CT,0 kg N·hm-2·a-1)、低氮(LN,40 kg N·hm-2·a-1)、高氮(HN,80 kg N·hm-2·a-1)3种处理(2011年氮沉降背景值36 kg N·hm-2·a-1),每个处理4个重复,实验详细设计见参考文献[7, 12]。
1.3 凋落物收集与处理
凋落物的收集采用直接收集法。于2012年12月在不同处理的每块样地中均随机布置5 个0.71 m×0.71 m 的凋落物框,共60个。凋落物框为60目尼龙网裁成的圆锥形网兜,框架离地面约50 cm,网兜深度为0.3 m。在2013—2015年每月月底收集1次。将凋落物框中的凋落物取回后,按叶(米槠叶)、杂叶(除米槠叶之外的叶)、枝、繁殖器官(花和果)、杂物(包括虫鸟动植物残体、碎屑等)等5部分进行分取,分取后在65 ℃ 恒温箱中烘至恒重,再分别进行称重。
1.4 数据处理与分析
绘图采用 Origin 2020 软件,数据分析运用R软件(R.3.6.1, R-project software, 2019)。利用单因素方差分析,对不同氮处理下年均凋落物量及各月各组分凋落量进行差异显著性检验,采用最小显著差异法(LSD法)进行多重比较。采用双因素重复测量方差分析检验氮沉降时间和氮沉降水平对总凋落物量和各组分的影响。
2 结果与分析
2.1 氮沉降对年凋落量及其组分月动态的影响
米槠天然林凋落物有大小年现象,总体呈波动性(图1)。2013年和2015年凋落物月动态呈三峰型,峰值分别出现在2、6、9月和3、5、9月。2014年凋落物月动态呈双峰型,峰值分别出现在4月和9月(图1a)。
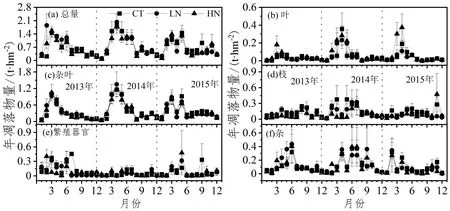
图 1 不同处理凋落物月动态变化Figure 1 Monthly dynamic in litter production of different nitrogen treatments
米槠天然林凋落物叶动态变化基本呈单峰型变化,但每年出现峰值时间略有差异。2013、2014和2015年出现峰值的时间分别为当年的3月、4月和4月(图1b)。2013年和2014年杂叶的月动态变化模式基本叶动态一致,而在2015年在3月份和5月份出现2个峰值;凋落物枝的变化无明显的季节变化模式(图1d);凋落物繁殖器官的动态变化年际变化较为明显,2013年和2015年月际变化较大,而2014年月动态变化较小(图1e)。凋落物杂物的变化较为复杂,2013、2014和2015年分别呈单峰型、三峰型和双峰型变化(图1f)。总之,氮沉降处理并未改变凋落物总量及其各组分的月动态变化模式(图1)。

注:不同小写字母代表不同氮处理间差异显著(P<0.05)。图 2 氮沉降对凋落物各组分和总量的影响 Figure 2 Effects of nitrogen deposition on the composition and total amount of litter

表 1 氮沉降水平和时间对凋落物各组分及其总量的影响 Table 1 Effects of nitrogen deposition level and duration time on the composition and total amount of litters
2.2 氮沉降对凋落量各组分及比例的影响
总体来讲,除2013年LN处理显著提高凋落物总量外(P=0.01),连续3年氮沉降处理并未改变凋落物及其各组分的总量(图2a)。3年CT,LN和HN处理年均凋落物总量分别为8.83 t·hm-2,8.99 t·hm-2和8.03 t·hm-2。氮沉降各个组分总量均无显著影响(图2b,c,d,e,f)。双因素重复测量方差分析表明,氮沉降时间对繁殖器官有显著影响(表1,P=0.04),而对其他组分和总凋落物量无显著影响;氮沉降水平对总凋落物量和各组分量均无显著影响,而且氮沉降时间和水平对总凋落物量和各组分均无交互作用(表1)。
在CT处理中各凋落物组分所占总凋落物比例大小分别为杂叶、枝、杂、繁殖器官和叶,但氮沉降处理改变了各组分所占总凋落物的比例(图3)。氮沉降显著降低枝占总凋落物量的比例,并随氮沉降量增加而降低(图3)。与CT(19.3%)相比,LN和HN处理中枝条所占比例分别降低至14.3%和12.5%。

图 3 氮沉降对凋落物各组分比例的影响Figure 3 Effects of nitrogen deposition on the proportion of litters components
3 讨论
3.1 凋落物动态变化
本研究结果表明,年凋落物量在7.05 ~10.25 t·hm-2·a-1之间,平均值为8.61 t·hm-2·a-1,与鼎湖山南亚热带常绿阔叶林平均年凋落物量(8.61 t·hm-2·a-1)[6]和安徽肖坑亚热带常绿阔叶林的凋落物量(6.91~9.85 t·hm-2·a-1)接近[13],高于中国亚热带天然林平均年凋落物量(6.66 t·hm-2·a-1),略低于中国热带天然林年凋落物量(9.30 t·hm-2·a-1)[14]。此外,本研究凋落物量基本高于中国的主要人工林年凋落物量[14]。由此可见,凋落物量的大小不仅和林分结构、树种有关,还与所处气候带有很大关系。研究表明森林年凋落物量均随纬度升高而减少,纬度每升高1°,年凋落物量下降0.11~0.27 t·hm-2[14-15]。 森林凋落物月动态变化规律因气候条件、森林类型和树种的组成而异,凋落物的月变化模式一般呈单峰型、双峰型或不规则型[14-16]。本研究中,凋落物的月动态变化基本呈双峰型(图1a),第一个峰值主要出现在3—4月份,这是由于每年3—4月份是常绿阔叶林开花季节,同时春季萌发新叶并旺盛生长,促进衰老叶的凋落,导致凋落物量增加[6]。而第二个峰值一般出现在夏季或秋季。本研究区处于台风区,每年夏季均有台风过境,受台风影响当月的凋落物量明显增加,而秋季是植物叶片衰老季节,大量叶片和果实脱落,因而形成峰值。另外,由于各器官的形成和发育特别不相同,各器官形成的峰值时间也不相同。
3.2 氮沉降对凋落物及其组分的影响
氮沉降对凋落物产量的影响与气候带、林分类型、氮沉降量的不同有关[12,14]。本研究通过3年的定位研究发现,LN处理除在试验初期表现为增加凋落物总量外,随着时间推移,并未影响总凋落物量。相对于CT,连续3年的HN处理对凋落物及其组分总量的影响并不显著。研究结果与亚热带常绿阔叶甜槠林和杉木人工林部分研究结果一致[9-10]。从凋落物对处理时间和氮沉降水平响应的差异来看,森林生态系统土壤氮有一定的饱和点[16],在未达到饱和之前,氮沉降增加会提高森林生产力和凋落物产量[9-10],但超过氮饱和点时,可能抑制林木的生产力,进而降低凋落物产量[4]。但本研究中并未发现氮沉降对凋落物产量显著影响,这可能与本地区降雨量较大(观测期间降雨量1 906 mm)有关,而充足的降雨量将对土壤产生较大的淋溶作用,大量的氮可能通过淋溶作用流出生态系统[17]。此外,凋落物对氮沉降的响应与氮输入水平有关,如刘文飞等[10]对亚热带杉木人工林模拟氮沉降研究表明,低氮处理凋落物量无显著变化,中氮处理凋落物量显著增加,而高氮处理表现为先增加后抑制的趋势;赵鹏武等[9]对兴安落叶松研究结果也表明,低氮和中氮氮沉降对凋落物量无影响,而高氮处理会降低凋落物量。本研究结果表明氮沉降水平和时间对凋落物量均无影响(表1),这可能由于本研究氮沉降时间相对较短(3年),还未超过土壤氮饱和点。
本研究发现,不同氮沉降水平对凋落物各组分无显著影响(图2),但改变了枝所占总凋落物量的比例(图3)。有研究表明,氮沉降在一定程度上抑制了凋落物叶的产生,而促进果凋落物和皮凋落物的产生[18]。如李德军等[19]研究南亚热带两种乔木幼苗生物量分配对氮沉降的响应结果表明,中等水平氮沉降量提高了叶的生物量,高氮沉降量抑制叶的生物量,这可能与不同树种对氮沉降的响应机制有所不同有关[10]。此外,本研究双因素方差分析结果表明,氮沉降持续时间对繁殖器官具有显著影响(表1,P=0.04),花果凋落物量的增加可能由于持续氮输入导致土壤氮过量使其不易坐果,进而增加花果的凋落物量。
4 结论
通过连续3年模拟氮沉降试验,结果表明氮沉降短期内对凋落物的无显著影响,但持续氮沉降增加改变了枝所占总凋落物的比例。凋落物是森林碳循环中的重要组成部分,但由于森林生态系统的群落结构复杂,短期试验结果难以反映长期的结果。因此,在未来的全球变化背景下,随着全球氮沉降量的持续增加,要更深入了解氮沉降对米槠常绿阔叶林凋落物量的影响还需进行长期定位研究。
猜你喜欢
地下水(2023年5期)2023-10-13 06:03:28
山西林业(2021年2期)2021-07-21 07:29:28
农村百事通(2020年21期)2020-11-23 02:04:24
绿色中国(2019年19期)2019-11-26 07:13:16
绿色中国(2019年19期)2019-11-26 07:13:16
亚热带农业研究(2019年2期)2019-09-05 09:39:06
植物研究(2017年5期)2017-11-10 01:47:20
星星·诗歌原创(2017年5期)2017-08-13 10:09:55
五邑大学学报(社会科学版)(2016年2期)2016-03-13 09:24:47
中国果菜(2015年2期)2015-03-11 20:01:08
