
镧对铜、镉单一胁迫下荻幼苗的生长及生理特性的影响
2020-10-24 03:41魏媛媛谢思敏
安徽师范大学学报(自然科学版) 2020年5期
马 冰, 张 杰, 魏媛媛, 谢思敏
(1.安徽师范大学 生命科学学院,安徽 芜湖 241000;2.安徽省重要生物资源保护与利用研究重点实验室,安徽 芜湖 241000)
近年来,随着我国经济的快速发展,土壤中铜镉污染状况越发严重[1]。在诸多治理方法中,植物修复技术因其环境友好,且不产生二次污染等特点而受到人们的重视。研究者在选择治理植物时倾向于超积累植物,但其往往生长缓慢,生物量小,导致实际修复中无法有效应用[2]。有研究发现荻[Miscanthussacchariflorus(Maxim.)Benth.]生长迅速,生物质产量高[3],对铜和镉具有较强的耐性和稳定化能力[4-5],同时又是一种很重要的能源植物。此外,荻是多年生植物,这使得连续修复易于开展,有助于缓解能源压力和环境保护[6]。而且荻各器官中的金属含量具有元素特异性,地上部镉的含量高于根部,相反,根部铜含量高于其他器官[7],可以使地上部和根部分别富集重金属镉和铜。因此,荻在重金属污染土壤修复方面必将具有广阔的应用前景。当植物生长在重金属污染的土壤中时,植物体内的镉达到一定的浓度时,会影响植物的光合作用,抑制植物保护酶的活性等,使植株发育缓慢,导致植株矮小[8]。铜虽然是植物体内所需的微量元素之一,但当土壤中铜浓度提高至某一阈值时,会抑制植物的光合作用,减缓植物生长[9]。镧(lanthanum,La)元素作为稀土元素之一,在有机体中含量很少,但对生命活动具有重要的调节作用。王蕊研究表明,5~10mg/L的稀土镧元素可以提高豌豆幼苗的抗氧化酶活性,提高植物对外界重金属胁迫的抗逆性[10];加西亚等人研究表明低浓度的镧可以提高植株的生物量[11];孙嘉欣研究表明低浓度的镧可以促进野生扁穗雀麦幼苗根系的生长,降低幼苗的相对电导率,减轻细胞膜的受损伤程度[12]。但是将荻作为研究对象,研究稀土元素镧对重金属胁迫下植物生长产生的影响还鲜见报道。
因此本实验以荻为实验材料、La(NO3)3(10mg/L)作为调节因子,研究镧对不同浓度铜、镉单一胁迫下荻幼苗生长及抗氧化酶活性的影响以及对Cu2+、Cd2+在荻体内的富集特征进行研究比较,为稀土元素在重金属污染方面的研究以及荻修复重金属污染土壤提供依据。
1 材料与方法
1.1 幼苗的培养
供试验植物荻种子于2017年12月18日采自芜湖市近郊,采集荻穗,经自然风干后,剥离并挑选籽粒饱满的种子。沙子于2018年3月22日采自安徽师范大学赭山校区操场沙坡,过2mm孔径筛,用自来水冲洗干净后,再用蒸馏水冲洗3次,放入恒温箱110℃杀菌消毒3h后,放入塑料盆中备用。将荻种子用75%酒精消毒杀菌60s,用蒸馏水冲洗干净,静置风干一夜。将处理过的种子整齐放入沙盆中,加水湿润,放入30℃气候箱中催芽。待荻种子出芽后,改浇改良的Hangland营养液。
1.2 幼苗处理
将沙培60d后的荻幼苗分别移入高13.5cm,底部直径13cm,杯口直径17cm的塑料杯中,加改良的Hoagland营养液分组稳苗10d,稳苗结束后对分组的幼苗进行处理(表1)。处理后的营养液中不再施加NH4H2PO4,改为每天日照结束后向叶面喷洒,目的是为了防止和La(NO)3、CdCl2形成沉淀。按照处理分为10组,每组15杯平行,每杯十株长势基本相同的荻幼苗,每杯一次性加入1L营养液,试验在自然光照下进行。

表1 幼苗处理
荻培养阶段于2018年3月24号开始,2018年6月2号结束,实验地点在安徽师范大学生态学实验室。前期培养(3月24日—5月23日)采用人工气候箱培养,培养条件为日间14h,28℃,80%光照强度,夜间10h,22℃,无光,每天添加1次营养液;后期稳苗(5月23日—6月2日)采用室内自然方式培养。2018年6月2日开始取样。
1.3 指标测定
1.3.1 超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性和过氧化氢酶(CAT)活性的测定 称取各株相同位置荻叶片0.5g,分次加入5ml预冷的50mmol/L pH=7.8的磷酸缓冲液在冰浴中研磨成匀浆,在冷冻离心机中于4℃下15000r/min离心15min,取部分上清液经适当稀释后用于酶活性测定。SOD活性测定采用NBT光化还原法,以抑制NBT光化还原50%的酶量为一个酶活力单位[13];POD活性测定采用愈创木酚法,以每分钟吸光度值变化(升高)0.01为1个酶活性单位(U)[14];CAT活性测定使用紫外吸收法,以每分钟A值减少0.01为一个酶活性单位(U)[15]。
1.3.2 叶绿素含量和丙二醛(MDA)含量的测定 叶绿素含量采用浸提法,称取0.2g叶片放入乙醇丙酮混合液(乙醇∶丙酮=1∶1)10ml中,放置于避光处,40h后叶片完全变白,取浸提液用紫外分光光度计测定[16];丙二醛(MDA)含量测定采用硫代巴比妥酸(TBA)显色法[17]。
1.3.3 生长指标及铜、镉质量分数的测定 用直尺测量荻幼苗的株高、根长,用分析天平称量荻幼苗的整株重量,记为鲜重。将荻幼苗的根部、地上部分开,分别放到干燥箱中100℃杀青30min、80℃烘干至恒重。然后将根部、地上部的样品分别消化,送至校外专业机构检测其中铜、镉和镧的含量。
在第0、9d,测量荻幼苗的生长指标;在第0、3、6、9d,测定荻幼苗的SOD、POD和CAT活性、MDA含量、叶绿素含量;在第9d采样测定荻幼苗体内的铜、镉和镧含量。为遵循重复性原则,每一指标测定3次。
1.3.4 数据统计分析 实验数据的处理和分析采用SPSS 21.0软件完成。
2 结果与分析
2.1 镧对铜、镉单一胁迫下荻幼苗生长特征的影响
植株的根长、株高以及单株重量可以直观地表现出植株的生长情况,La组与CK组相比,荻幼苗的株高、根长和鲜重均有提高,说明稀土元素镧对荻幼苗生长有一定的促进作用。由表2可知,在不同铜浓度胁迫下,镧提高了25mg/L铜浓度胁迫下荻幼苗的株高、根长和鲜重,降低了100mg/L铜浓度胁迫下荻幼苗的株高和根长(P>0.05)。在不同镉浓度胁迫下,镧显著提高了5mg/L镉浓度胁迫下荻幼苗的根长(P<0.05),降低了20mg/L镉浓度胁迫下荻幼苗的株高和根长,植株鲜重有所升高(P>0.05)。说明铜、镉浓度低时,镧对荻幼苗生长有一定的促进作用,但随着铜、镉胁迫强度的加强,镧与铜、镉发生了协同作用,加深了对荻幼苗生长的胁迫影响。

表2 镧对铜、镉单一胁迫下荻幼苗生长特征的影响
2.2 镧对铜、镉单一胁迫下荻幼苗叶绿素含量的影响
由表3可知,第3天时,Cu2La,Cd1La,Cd2La组的叶绿素a/b高于Cu2,Cd1,Cd2组,但无显著性差异(P>0.05),各组的叶绿素总量无显著性差异(P>0.05)。第6天时,La组的叶绿素a/b显著高于CK组(P<0.05),是CK的1.45倍;Cu2La组和Cd2La组的叶绿素总量分别高于Cu2组和Cd2组(P>0.05),La组低于CK组(P>0.05)。第9天时,Cu1La组和Cd1La组的叶绿素总量分别显著高于Cu1组和Cd1组(P<0.05),分别是无镧组的1.91倍和2.03倍。我们可以通过叶绿素总量与叶绿素a/b的数值衡量植物在逆境下受到的伤害大小,一般来说,植物的叶绿素总量越大,叶绿素a/b数值越高,其生长情况也就越好。可以看出,随着时间的延长,镧可以缓解低浓度铜、镉重金属对荻幼苗的伤害,促进叶绿素的积累与幼苗生长。
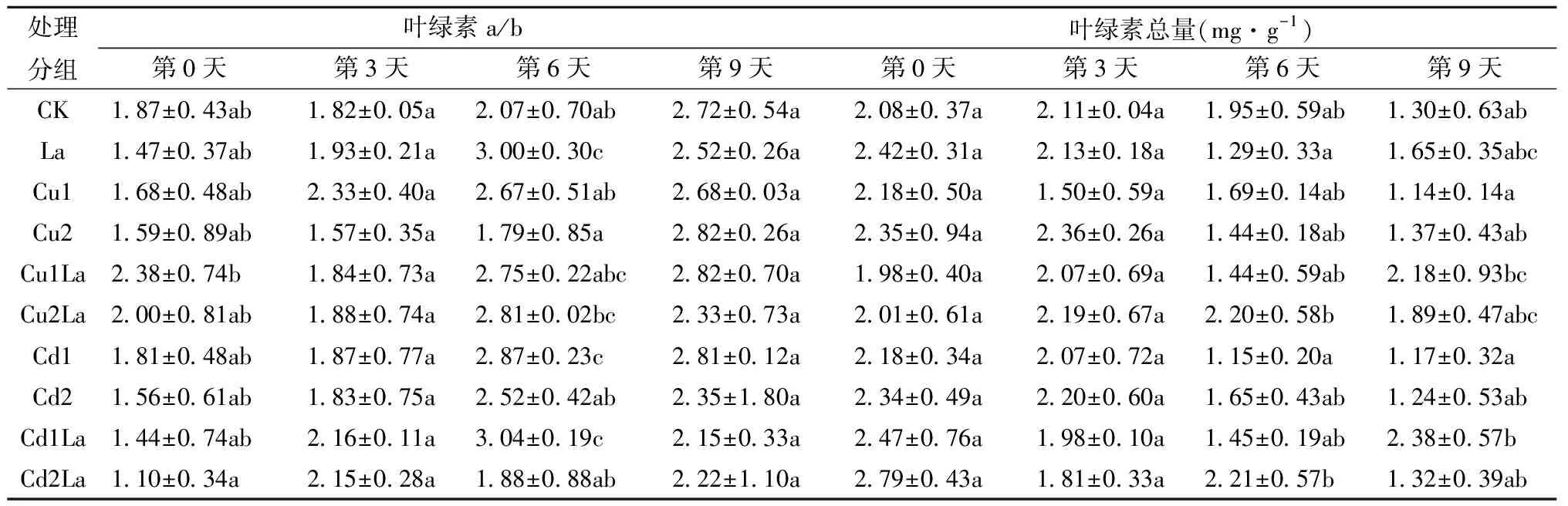
表3 镧对铜、镉单一胁迫下荻叶片叶绿素a/b及叶绿素总量的影响
2.3 镧对铜、镉单一胁迫下荻幼苗SOD活性的影响
由表4可知,第3天时,Cd1组与Cd1La组有显著性差异(P<0.05),Cd1La组是Cd1组的1.67倍。第9天时,Cu1、Cd2组低于Cu1La、Cd2La组,Cu2、Cd1组高于Cu2La、Cd1La,但差异不显著(P>0.05);Cu1La组显著高于其他组(P<0.05)。说明在镉胁迫下,初期镧有提高SOD活性的潜力,但与Cd浓度和时间有关,随着镉胁迫强度的增强,镧可能与镉发生了协同作用。而在铜胁迫下时,镧也有提高铜胁迫下荻幼苗SOD活性的能力,与Cu浓度有关,低浓度的铜胁迫下效果更为显著。La组相对于CK组,SOD活性随时间先升高后降低。说明镧有着提高荻植株SOD活性的能力,但是这与时间长短有关。

表4 镧对铜、镉单一胁迫下荻幼苗SOD活性的影响
2.4 镧对铜、镉单一胁迫下荻幼苗POD活性的影响
由表5可知,Cd处理组中第6天和第9天时加镧组均高于无镧组,且第9天时Cd2La组显著高于Cd2组(P<0.05),是其2.37倍。Cu处理组中加镧组也均高于无镧组,但差异不显著(P>0.05)。说明在Cu、Cd分别胁迫下,镧有提高荻幼苗POD活性的能力,且可以显著减轻高浓度镉对荻幼苗的危害,提高了荻幼苗POD活性。

表5 镧对铜、镉单一胁迫下荻幼苗POD活性的影响
2.5 镧对铜、镉单一胁迫下荻幼苗CAT活性的影响
由表6可知,第3天时,除La组外其他实验组的CAT活性均低于CK组,但差异性不显著(P>0.05)。第6天时,La组和CK组数值接近,Cu2、Cd1、Cd2、Cu1La、Cu2La、Cd1La、Cd2La组均显著低于CK组(P<0.05),分别为CK组的31.41%、32.76%、26.35%、22.44%、15.96%、9.68%、36.28%。第9天时,实验组均显著低于CK组(P<0.05)。说明镧有提高CAT活性的能力,但与时间有关,随着时间的增长,镧可能与铜和镉发生协同作用降低了CAT活性。

表6 镧对铜、镉单一胁迫下荻幼苗CAT活性的影响
2.6 镧对铜、镉单一胁迫下荻幼苗MDA含量的影响
由表7可知,第3天时La组显著高于CK组(P<0.05),是其1.97倍。第6天时,Cu1、Cd1组低于Cu1La、Cd1La组,Cu2、Cd2组高于Cu2La、Cd2La组,但无显著性差异(P>0.05)。第9天时,Cd2La组显著低于Cd2组(P<0.05),且Cd1La组显著高于Cd1组(P<0.05),说明镧在Cd(20mg/L)胁迫下具有保护作用,在Cd(5mg/L)胁迫下可能是因为荻本身对低浓度镉具有抗性,而镧的添加使得质膜透性更大,导致MDA含量升高。Cu处理组均与CK组无显著性差异。
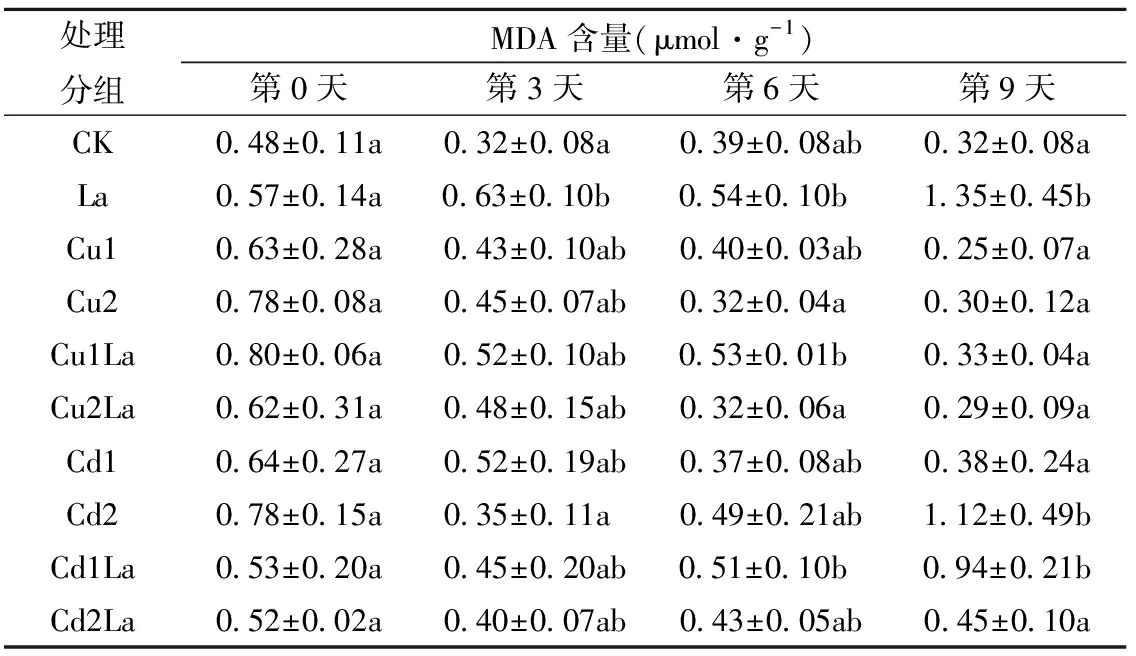
表7 镧对铜、镉单一胁迫下荻幼苗MDA含量的影响
2.7 镧对铜、镉单一胁迫下荻幼苗体内铜、镉分布和富集的影响
由表8可知,第9天时,Cu1La组地上部Cu浓度低于Cu1组,但差异不显著(P>0.05),而Cu2La组地上部Cu浓度显著高于Cu2组(P<0.05),是其1.43倍。Cu1组根部Cu浓度高于Cu1La组,Cu2组低于Cu2La组,但无明显差异(P>0.05)。Cd2La组地上部La浓度显著高于La组(P<0.05)。Cd1La组地上部Cd浓度高于Cd1组(P<0.05),Cd2La组地上部Cd浓度显著高于Cd2组(P<0.05)。加镧的镉处理组根部Cd浓度均低于无镧的镉处理组,但无显著性差异(P>0.05)。转运系数是指植物地上部中的重金属含量与地下部中的金属含量的比值,Cd1La组和Cd2La组镧的转运系数均大于Cu1La组、Cu2La组和La组,但无显著性差异(P>0.05)。Cd1La组和Cd2La组铜转运系数均大于1,且与CK组有显著性差异(P<0.05),说明镉和镧的添加促进了荻地上部对铜的富集。Cd1La组中镉的转运系数显著大于Cd1组(P<0.05),说明镧的添加提高荻植株地上部对镉的富集,且与镉胁迫的浓度有关,在镉浓度为5mg/L时效果显著(P<0.05),但是本实验为水培,具体机制有待于后期进一步的研究。

表8 镧对铜、镉单一胁迫下荻幼苗中镧、铜和镉质量分数及转运系数的影响
3 结论和讨论
株高、根长是植物重要的生长指标,叶绿素总量、叶绿素a/b是植物重要的生理指标。刘建新等人研究表明低浓度硝酸镧(10~50mg/L)能够提高黑麦草种子的活力,促进幼苗的生长[18];克嘉文等人研究表明低浓度La3+处理对大豆幼苗的生长和光合作用有明显的促进作用[19]。不同浓度的Cu、Cd胁迫会使植物叶绿素含量减少,同时抑制根的生长,对植物生长产生胁迫。本研究中,叶绿素总量在第9天时与La浓度间存在显著的正相关关系(P<0.05),相关系数为0.762,说明镧提高了荻幼苗叶绿素总量,其原因可能是因为镧的加入促进了荻幼苗对氮和磷的吸收,从而诱发了叶绿素前体化合物的大量合成,同时也可能作为相关酶的活化剂增加了叶绿素的合成[20]。荻幼苗的鲜重和Cd浓度间存在显著的负相关关系(P<0.05),说明随着Cd浓度的上升,荻幼苗生长状况变差,其原因可能是因为镉不是植物所必需的的微量元素,所以相对于铜来说,镉对荻幼苗的胁迫更加严重。由此可见10 mg/L的La3+的加入可以提高荻幼苗的叶绿素总量,镧可能诱发了叶绿素前体化合物的大量合成,参与了提高植物体内叶绿素的生理生化反应,加强了荻幼苗的光合作用,提高了荻幼苗的生存能力。

实验结果表明,铜和镉在荻幼苗内的富集情况为:根部>地上部。铜处理的浓度越大,在植物体内富集越多,荻幼苗地上部和根部的铜含量与铜浓度间存在显著正相关关系(P<0.05),相关系数为0.737和0.994,且荻根部铜含量(y)与铜处理浓度(x)间存在线性关系,线性方程为:y=0.772x-7.435,R2=1,(P<0.01),说明荻幼苗体内铜含量与铜处理浓度密切相关。同一铜处理浓度下,Cu2La组地上部的铜含量显著高于Cu2组(P<0.05),可能是因为铜本身是植物体内多种抗氧化酶的重要组成部分[27],镧的加入提高了荻体内抗氧化酶的数量,从而提高了荻体内的铜含量。荻幼苗根部的镉含量也和镉处理浓度有着正相关关系(P<0.05),相关系数为0.987,且荻根部镉含量(y)与镉处理浓度(x)间存在线性关系,线性方程为:y=0.872x+0.439,R2=0.998,(P<0.05)。荻幼苗地上部La含量与镉处理浓度有着正相关关系(P<0.05),相关系数为0.922;而根部La含量与铜处理浓度有着正相关关系(P<0.01),相关系数为0.979。可能是镧与镉和镧与铜之间在荻体内启动了某种防御机制,促进了荻地上部对镉和镧的吸收以及根部对铜和镧的吸收。说明镧在一定程度上可以缓解铜和镉对荻幼苗生长的胁迫,提高荻体内抗氧化酶的活性,而另一方面又可以提高荻地上部和根部对铜、镉重金属产生特异性的富集。镧的加入促进了荻幼苗根部对铜的富集,加强了荻根部对镧和铜的吸收,镧加强了荻幼苗体内的镉从根部对地上部的转运能力,提高了荻幼苗地上部富集镉的能力,稀土元素镧提高了荻幼苗对重金属铜、镉的吸收富集能力。因此认为,镧在荻治理铜、镉重金属污染土壤方面具有一定的应用潜力和前景。
猜你喜欢
今日农业(2021年10期)2021-11-27
阅读(科学探秘)(2020年8期)2020-11-06
西藏文学(2020年5期)2020-09-21
快乐语文(2019年35期)2020-01-06
中国诗歌(2019年6期)2019-11-15
绿色科技(2019年2期)2019-05-21
女性天地(2016年10期)2017-04-25
腹腔镜外科杂志(2016年11期)2016-06-01
中国火炬(2015年12期)2015-07-31
食品工业科技(2014年6期)2014-05-10
