
IgA肾病相关的免疫细胞膜分子及系膜细胞激活的研究进展
2020-10-23 01:02赵成波王润秀谢富华
赣南医学院学报 2020年8期
赵成波,曾 欢,王润秀,谢富华
(1.赣南医学院第一附属医院肾内科;2.赣南医学院基础医学院分析实验室,江西 赣州 341000)
IgA 肾病是一组以系膜区IgA 或IgA 复合物沉积为主要特征的原发性肾小球疾病。以低糖基化的IgA1 为抗原形成的巨大特异性抗原抗体复合物沉积在肾小球为主要病理特点,以血尿、蛋白尿及肾功能减退为主要临床表现。由法国病理学家BERGER 和HINGLAIS 在1968 年最先报道[1],也是世界范围内最常见的原发性肾小球疾病,20%~40%的患者进展为终末期肾病,同时也是终末期肾功能衰竭、透析、肾移植的主要原因[2]。虽然本病的发病机制尚不完全清楚,但也取得许多可喜的成绩,无论是四重打击学说的提出,还是免疫细胞膜分子的鉴定都具有重要意义。因此,本文就免疫复合物的形成及沉积、免疫细胞膜分子(CD71、TG2、sCD89)和系膜细胞的激活的最新研究进展做一综述。
1 免疫复合物的产生
见图1。
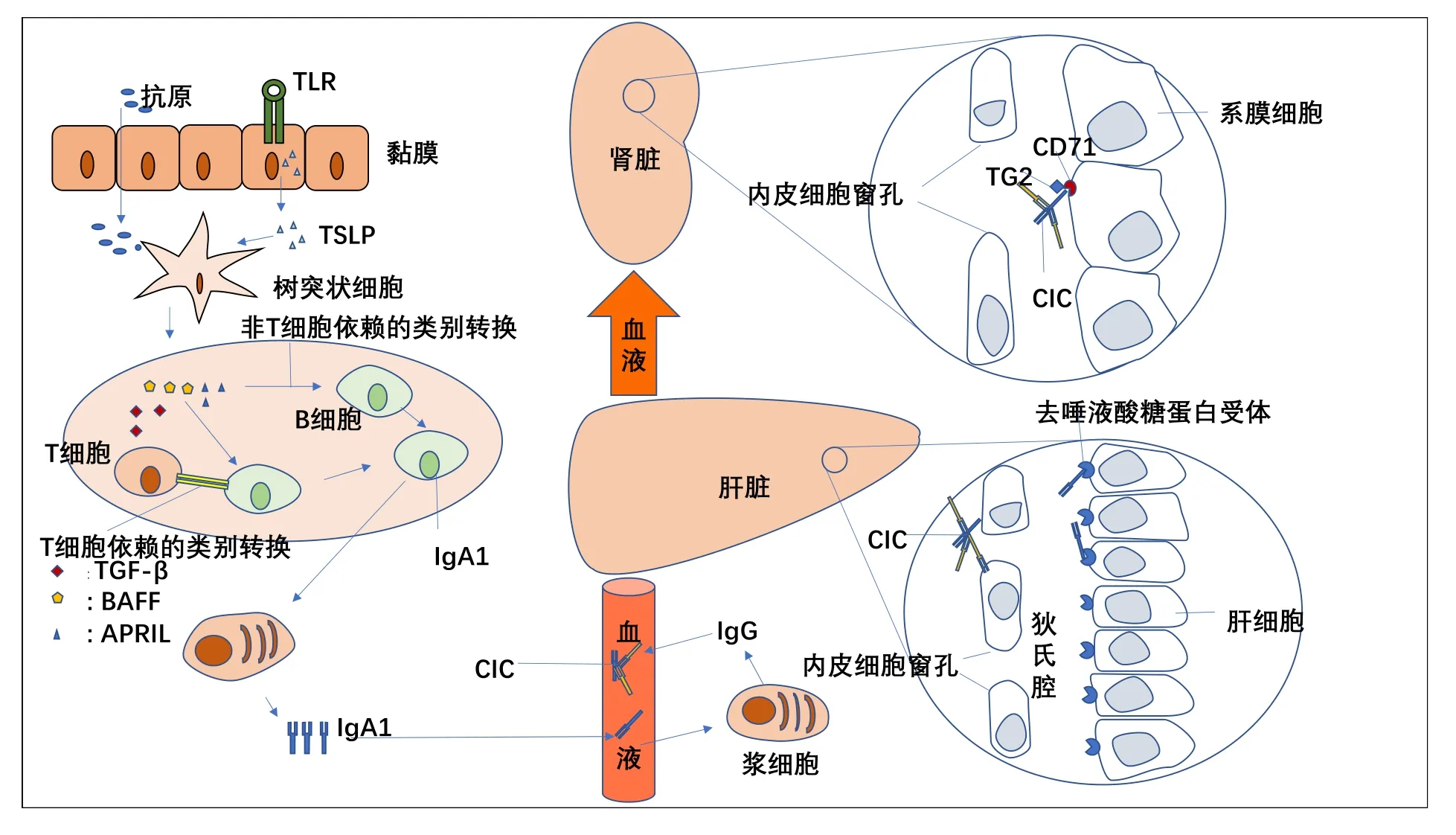
图1 IgA1的产生及循环免疫复合物形成及肾脏沉积病理过程
1.1 黏膜免疫反应与食物抗原大多数研究认为IgA 肾病的发病与扁桃体感染、肠道黏膜感染以及食物抗原引起树突状细胞活化,进而使B 细胞活化而产生的低糖基化的IgA1 密切相关。约三分之一IgAN 患者体内发现了食物抗原的IgA 循环复合物[3],特定的食物抗原能加剧IgAN 的发展,但当去除以谷蛋白为主的食物抗原仅能降低血尿及蛋白尿,并不能改变肾功能下降的速率。然而在黏膜感染急性期几乎均会伴发血尿,这可能是抗原通过刺激IgAN 患者循环血细胞毒效应细胞(Tc 和γδT 细胞)上CX3C趋化因子受体1(CX3CR1)的表达,从而促进淋巴细胞在肾小球跨内皮迁移,导致肾小球连续性的破坏而诱发血尿[4]。值得注意的是CX3CR1阳性的Tc细胞在IgAN患者的扁桃体中更常见[5],同时扁桃体能产生大量的IgA1加重疾病的发展[6-7],因此对于扁桃体反复感染的IgAN 患者切除扁桃体是具有可靠理论依据的。
1.2 低糖基化IgA1的产生IgA1是机体免疫反应的重要组成部分,人类每天每公斤体重大约分泌70 mg IgA,其中大部分都是含有两条重链两条轻链的单体IgA1。既往研究证实COSMC 基因表达异常是低糖基化IgA1 产生的关键。一些特异性的细菌通过细胞壁的脂多糖诱导机体产生IL-6 进而激活B细胞上的JAK/STAT3 信号通路导致STAT3 核移位而导致半乳糖缺乏的IgA1 产生,同时它也能激活TLR4 诱导COSMC 甲基化水平升高而使β1,3-半乳糖基转移酶(C1Galt1)活性降低进一步导致半乳糖缺乏的IgA1产生[8]。诸多的证据表明产生低糖基化的IgA1 诱因并不单一,IL-4、IL-17、TGF-β、IL-6 等细胞因子均可促进C1GALT1C1 基因启动子CpG 岛甲基化,导致C1GalT1C1 mRNA 表达下调,从而使B 细胞分泌低糖基化IgA1[9-12]。同时它们中的一些细胞因子如TGF-β、IL-10 等能直接参与IgM 发生类别转化促进IgA1 的生成。值得一提的是B 细胞成熟的两个关键因子增殖诱导配体(APRIL)和B 细胞活化因子(BAFF)在IgAN 中的作用逐渐明确[13],在小鼠实验中证实BAFF 不但可以加剧IgA1 的产生,而且参与了IgA1 在肾小球的沉积[14],同时亦能参与IgA类别转化[15]。然而黏膜中活化的B细胞可能并不是产生低糖基化IgA1 的根源,可能是因为初次免疫应答后B 细胞错误的归巢到骨髓,当再次免疫时骨髓中定居的记忆细胞会大量增殖,从而产生并释放半乳糖缺乏的IgA1 到达外周,近而诱导机体产生免疫复合,这不但解释了IgAN 血尿是间断出现,而且也解释了异基因骨髓移植后为什么IgAN 的组织学表现能完全缓解[16]。值得一提的是新的研究证实肾小球系膜细胞不但具有吞噬作用,亦能分泌IgA,而且有可能与免疫复合物沉积有关[17]。
1.3 IgA1 免疫复合物的形成单体的低糖基化IgA1并不能直接导致IgAN,但是因为其铰链区存在低糖基化糖链而被机体识别为新的抗原表位,从而刺激B 细胞分泌抗IgA1 的抗体IgG,从而形成抗原抗体复合物(circulating immune complexes,CIC),这些复合物包括GDIgA1-IgG、GDIgA1-sCD89、GDIgA1-gliadin 及IgA1-CIC-TMCD89[18](见 图2)。研究证实只有这些复合物的水平与疾病的活动及进展直接相关[19]。同时循环中的免疫复合能放大和延长IgAN进程[20]。少量的复合物并不会致病,现在认为肝脏是清除IgA1 的主要场所,但是它受诸多因素的影响,如:①复合物的分子量太大,不能进入肝脏的diss腔。②肝功能减退时去唾液酸糖蛋白受体(asialoglycoprotein receptor,ASGP-R)下调。③各种原因导致的IgA Fc(CD89)表达下降,CD89 介导的内吞减少。④IgA1 铰链区O-糖基化降低导致ASGP-R 结合降低[21],这可能是肝硬化患者能产生一种与IgAN十分相似的肾小球疾病的根本原因。

图2 IgA1-循环免疫复合物沉积的多靶点发病模型[18]
2 IgAN发病相关的重要免疫细胞膜分子
2.1 转铁蛋白受体(transferrin receptor,CD71)CD71广泛表达于人体各种组织中,尤其是增殖脂代谢旺盛的细胞,它是半乳糖缺陷的IgA1 免疫复合物 的 直 接 受 体[22]。在IgAN 患 者 中CD71 不 但 对IgA1及IgA1复合物表现出较高的亲合力[23],而且它的表达也异常升高,沉积的免疫复合物通可启动CD71 的过表达及自动扩增循环,从而增加IgA1 复合物系膜细胞的沉积[22-23]。这可能是因为:①它能参与并激活NF-kB 炎症信号通路;②促进了线粒体的氧化呼吸,从而增加了活性氧的产生;③细胞内铁含量异常从而影响其mRNA的表达。
2.2 转谷氨酰胺酶2(transglutaminase 2,TG2)转谷氨酰胺酶2 对于CIC 与系膜CD71 的结合稳定性起到关键性作用[24],当TG2 低表达或不表达时免疫复合物便不能与CD71 结合。研究证明:IgA1-sCD89 等免疫复合物可以启动自我扩增过程,它包括影响CD71 和TG2 的过表达,从而增加病理性IgA1 复合物在系膜沉积和慢性系膜细胞活化[24]。这个过程可能是因为TG2 与CD71 共同参与NF-kB的自身激活而导致TG2 的活性增强,再进一步激活NF-kB 通路,从而构成一个正反馈环[25]。然而TGF-β1、TNF-α、IL-1 和IL-6 的表达依赖于NF-κB 的激活,同时这些细胞因子也是TG2 的转录激活因子[26],这些细胞因子也在整个正反馈循环中起重要作用。由于存在这个正反馈也就能很好的解释部分患者在扁桃体切除后血尿明显改善,但是肾功能恶化并无改善。TG2 的重要性在IgAN 中表现在多个方面:①当系膜细胞TG2 表达下降时,CD71 的表达将减少[27]。②TG2 在IgAN 中的过表达与临床组织学病变的严重程度密切相关[27]。③TG2的过表达同时影响系膜细胞的代谢状态,同时它能通过改变细胞的自噬而调节系膜细胞的生长[25]。随着TG2 研究的深入,其更多功能可能会被逐渐揭开,这也可能为IgAN的治疗提供新的靶点与方向。
2.3 可溶性CD89(soluble CD89,sCD89) CD89主要表达于单核吞噬细胞及中性粒细胞表面,能与血清分泌型IgA1 结合而促进细胞的吞噬作用、释放炎症介质、超氧阴离子的产生及发挥ADCC 作用。在IgAN 患者的CIC 中发现较高水平的CD89 可溶性异构体(sCD89),而正常人CD89 水平较低,同时CD89 对IgA 只有低度亲和力,而对复合物的亲和力明显高于单体[28]。sCD89为CD71的配体,且能诱导CD71 的过表达[24],在活动性IgAN 患者随着时间的推移以较低且稳定的速率在CIC 中出现sCD89,这可能与系膜内CIC 的“诱捕”有关[29]。相比之下,稳定IgAN 患者CIC 中具有高且稳定的sCD89。双重转基因小鼠模型证实了IgA1-sCD89s 相互作用使CIC 在系膜上具有致病特性[30]。没有sCD89,IgA 似乎只与内皮细胞结合。而且在移植后复发的IgAN中IgA-sCD89 复合物水平显著高于异常。因此,sCD89可能与IgAN的发病机制及进展存在相关性。
3 系膜细胞的激活
系膜细胞的激活在IgAN中的重要性不言而喻,在免疫复合物与系膜细胞稳定结合后即能促进细胞增殖,合成系膜基质和释放细胞因子从而诱发肾小球肾炎。但是系膜细胞的激活机制复杂多样,至今尚未完全明确,传统观点认为系膜细胞的激活是半乳糖缺乏的IgA1 复合物激活CD71 导致。但最近研究对此提出了不同的观点:①β-1,4-半乳糖基转移酶是组成型表达的肾小球膜细胞IgA 受体,其在肾小球系膜IgA 清除和对IgA 沉积的初始反应中具有重要作用[31]。②新月体型IgAN 患者中的免疫复合物的水平明显高于非新月体型以及健康人,而非新月体IgAN患者和健康人之间则无明显差异,且健康人体内都存在一定水平的低糖基化IgA1 免疫复合物[32]。③在缺乏正常抗原驱动成熟的情况下形成的低亲和力先天样IgA 更容易参与IgA 肾小球沉积而诱发肾炎[33]。④大约50%的IgAN 患者血清中半乳糖缺乏的IgA1 是轻度升高或正常[34]。⑤在一定条件下系膜细胞本身就能表达IgA 且其本身具有吞噬作用[35]。这些都明显提示系膜细胞的激活可能并不是半乳糖缺乏的IgA1 及其复合物。因此,我们认为:IgAN 可能存在多种亚型,同时系膜细胞的激活也存在多样性,寻找新的潜在的激活物可能会是IgAN的一个新的研究方向。
4 展 望
IgAN 肾病的发病机制复杂多样,是由多因素共同导致,文中未一一提及,虽然随着研究的深入某些方面变的愈发清晰,但是目前尚无基于其发病机制的特异性治疗问世,因此对IgAN的机制研究仍然任重道远。然而研究IgAN 的免疫细胞膜分子如转铁蛋白受体(CD71)、转谷氨酰胺酶2(TG2)、可溶性CD89(sCD89)是非常有价值的,深入研究系膜细胞的激活机制可能会有意想不到的收获。
猜你喜欢
天津医科大学学报(2021年4期)2021-08-21
癌症进展(2018年11期)2018-12-30
中成药(2018年8期)2018-08-29
中成药(2018年3期)2018-05-07
中成药(2017年12期)2018-01-19
中成药(2017年8期)2017-11-22
中成药(2016年8期)2016-05-17
中华老年多器官疾病杂志(2016年7期)2016-04-28
医学研究杂志(2015年9期)2015-07-01
医学研究杂志(2015年7期)2015-06-22
