
杭州地区19个樱花品种无性系苗期生长适应性与生长节律研究
2020-10-22 09:26:30旷一明陈雅静柳新红
林业调查规划 2020年5期
旷一明,陈雅静,张 颖,唐 丽,柳新红
(1.祁阳挂榜山林场,湖南 永州 426100;2.中南林业科技大学林学院,湖南 长沙 410004; 3.浙江省林业科学研究院,浙江 杭州 310023)
樱花是指蔷薇科(Rosaceae) 李亚科(Prunoideae) 樱属(Cerasus) 的一类观花乔木,主要分布于中国、日本和朝鲜等国家[1-2]。随着“樱花热”的掀起,国内学者对樱花的研究越来越多,主要集中在繁殖技术[3-5]、遗传多样性[6-8]、资源和群落结构的调查研究[9-12]等方面。对于樱花花期生物学方面的研究大部分以物候为重点,而对于樱花花期早晚类型、形态多样性及变异和自身花期内源物质动态变化过程鲜有报道。本文以19个樱花无性系为对象,研究其生长状况及物候节律、生长量等指标,追踪它们的生长发育对周围环境变化的响应,进而探索樱花在杭州地区的生长适应性,以及环境对植物生长规律的影响,以期为樱花的园林应用提供理论依据。
1 试验地概况
试验在杭州浙江省林业科学研究院苗圃(30°13′N,120°1′E)进行。杭州地区处于亚热带季风区,属于亚热带季风气候,四季分明,雨量充沛。年均温17.8℃,平均相对湿度70.3%,年降水量 1 454 mm,年日照时间 1 765 h。
2 材料及方法
采用嫁接繁殖方法,砧木采用2 a生福建山樱花(CerasuscampanulataMaxim)实生苗,19个樱花(表1)1 a生枝条为接穗,于2015年12月进行嫁接。嫁接成活后移栽至苗圃地,采用完全随机设计,株行距60 cm×100 cm。正常田间管理,当砧木侧芽长出时进行抹芽处理,保留接穗顶芽成活。

表1 19个樱花无性系情况
2.1 苗期适应性调查
2.1.1 病虫害调查
于2017年6月中旬和9月中旬对试验植株进行病虫害情况统计,设置6个病虫害级别:(1)无发生:所调查的无性系植株叶片无病斑发生情况;(2)轻发生:植株叶片病斑面积占叶片总面积的20%以下;(3)偏轻发生:植株叶片病斑面积占叶片总面积的20%~40%;(4) 中等发生:植株叶片病斑面积占叶片总面积的40%~60%;(5)偏重发生:植株叶片病斑面积占叶片总面积的60%~80%;(6)全发生:植株叶片病斑面积占叶片总面积的80%以上。
2.1.2抗热性调查
于2017年8月中旬观测植株是否有日灼现象,此时日最高温达到40℃。有出现叶色变褐、变黄、萎蔫的现象计“1”,生长情况正常的计“0”。
2.2 物候观测
观测2年生19个樱花无性系嫁接苗是否开花与开花时期、展叶时期、叶变色期与落叶期。以植株叶和花物候状态达到整株50%的日期为准,具体标准见表2和表3。

表2 叶物候观测标准

表3 花物候观测标准
2.3 生长量测定
从2017年4月开始,每个月对植株的株高、地径进行测定。采用塔尺对株高进行测定,精确到0.01 cm。地径用游标卡尺测量,精确到0.01 mm。每个品种测量3株,取平均值。
2.4 数据处理
用Excel 2007软件对数据进行初步处理,运用SPSS 22进行相关性分析、主成分分析,采用Origin软件作图。
3 结果与分析
3.1 植株病虫害和日灼状况
19个无性系病虫害及日灼情况如表4所示。

表4 19个樱花无性系生长状况
从6月和9月的病虫害状况来看,主要病害为叶枯病、穿孔性褐斑病,常见虫害为介壳虫、蚜虫。兰兰樱、太白樱、阳春樱3种无性系在2次调查中都没有出现病虫害,表现出较强的抗病虫害能力,安徽野樱、吉野枝垂、绿樱和香味山樱则受病虫害影响比较严重,2次病虫害调查均表现出偏重发生或者全发生状态,抗病虫害性水平较低,其余12种无性系的抗虫害性介于两者之间,表现出轻发生或者偏轻发生状态。夏季杭州地区温度较高,过强的阳光直射造成植物叶片干旱失水,蒸腾作用减弱,发生日灼现象,叶片向阳面出现不均匀的褐色火烧状斑块。调查发现,安徽野樱、吉野枝垂、绿樱、太白樱、阳春樱和越之彼岸樱无性系在夏季高温下出现了不同程度的日灼现象,其他无性系生长情况良好。
3.2 物候观测状况
3.2.1 花期物候特征差异比较
试验的19个樱花无性系开花情况见表5。
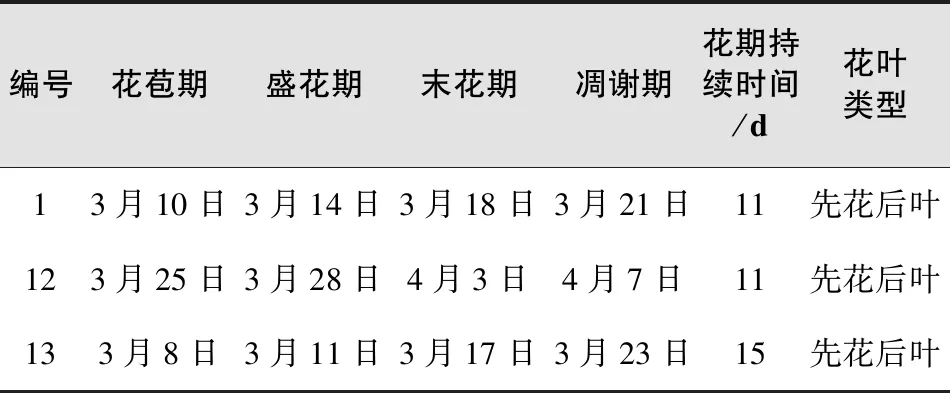
表5 3个开花无性系的物候差异
从表5中可以得知,于2017年春季开花的有安徽野樱、霞樱、香味山樱3种,其余的16个无性系不开花。其中香味山樱花期最早且持续时间最长,在3月8日就进入了花苞期,3月11日进入盛花期,3月23日凋谢,花期持续15 d。霞樱花苞出现的时间最晚,3月25日才进入花苞期,3月28日进入盛花期,4月7日凋谢,花期持续时间仅11 d,比13号樱花短了4 d;安徽野樱花期持续时间也为11 d。3种无性系植株都是先花后叶型。
3.2.2叶物候特征差异比较
3月15日到3月27日为叶芽萌芽期集中时间段,最早和最晚时间相差12 d,13号、17号、18号、12号和10号无性系叶芽萌发最早,14号、7号和11号无性系叶芽萌发最晚;3月24日到4月6日是叶芽开展期,最早和最晚时间相差14 d,7号、11号、14号和19号无性系展叶开始时间最晚;3月28日到4月12日为展叶盛期,最早和最晚相差16 d;9月18日到11月21日各无性系进入叶变色期,11月15日到1月2日落叶结束。山樱无性系叶片持续时间最长,有291 d,安徽野樱无性系绿叶时间最短,只有241 d,相差50 d(表6)。
3.3 基于叶的物候期划分
用X1、X2、X3、X4、X5分别代表叶芽萌芽期、展叶初期、展叶期、叶变色期和落叶期5个物候时期,对物候期进行划分。通过相关性分析和主成分分析,得到各物候间的相关矩阵(表7)和主成分特征值、贡献率、累计贡献率(表8)以及因子载荷矩阵(表9)。
在5个物候观测项目中,X1与X2、X3呈极显著相关,相关系数达到0.934和0.856,X2与X3极显著相关,相关系数0.921,X4与X5达到显著相关水平,相关系数0.459,其余物候项目间相关关系不显著。

表6 19个无性系叶物候开始期
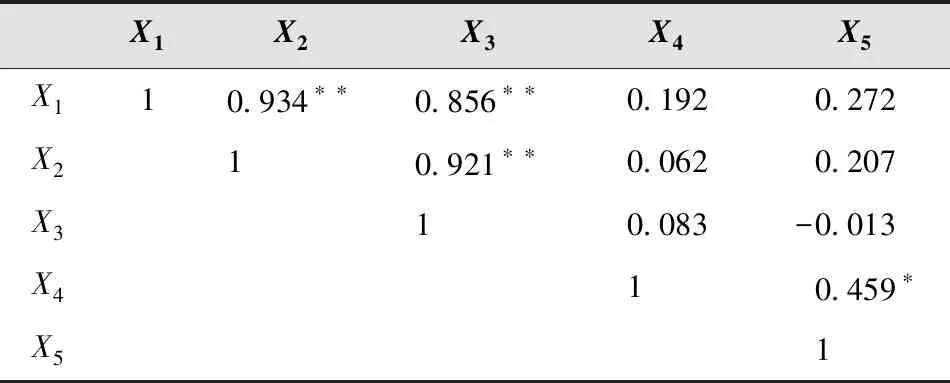
表7 物候期项目相关矩阵及显著性
如表8所示,第一和第二主成分累计贡献率达到了85.9%,基本可以解释原变量所包含的信息。因此可以将前2个主成分作为樱花无性系物候特征评价的综合指标。列出前2个主成分的函数表达式如下:
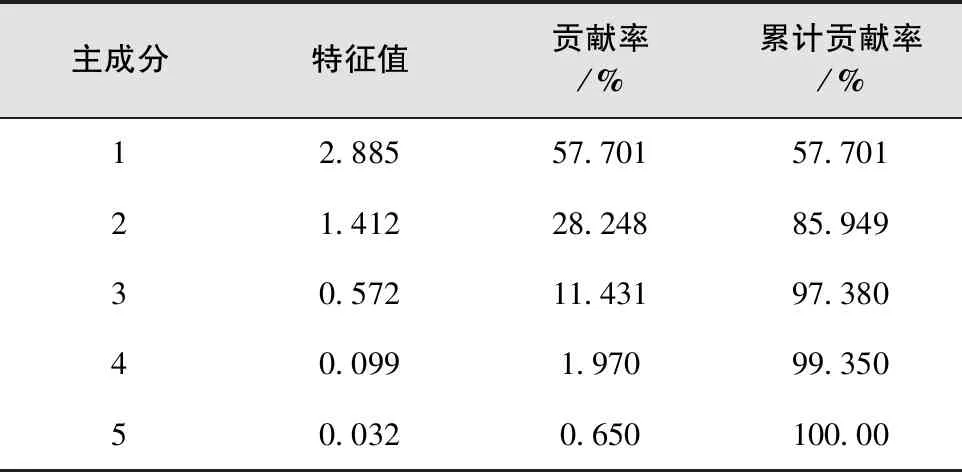
表8 主成分的特征值、贡献率、累计贡献率
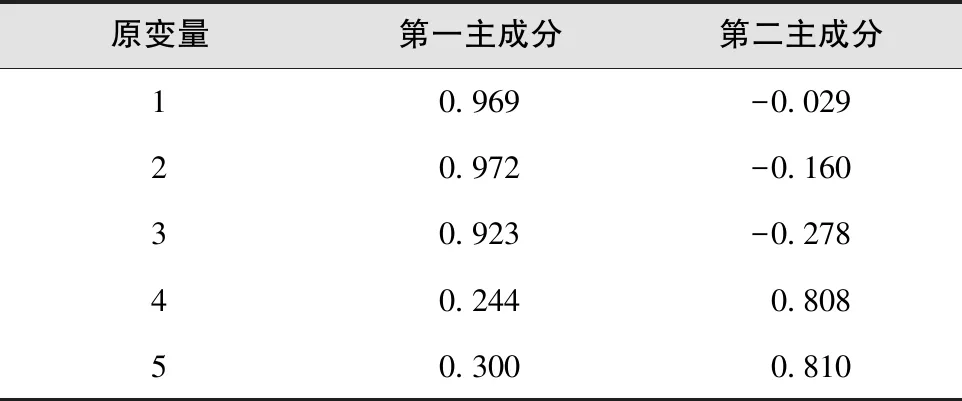
表9 因子载荷矩阵
Y1=0.57X1+0.57X2+0.54X3+0.26X4+0.18X5
Y2=-0.02X1- 0.13X2- 0.23X3+0.68X4+0.68X5
在主成分Y1中,X1(叶芽萌芽期)、X2(展叶初期)、X3(展叶期)具有最大的正系数,载荷较高,分别达到了0.969、0.972和0.923(表9),说明这3个物候期在第一主成分中起较大的正向作用;而X4(叶变色期)、X5(落叶期)2个在植物生长后期的物候期的系数比较小,说明第一主成分Y1反映的是植物生长后期的物候特点。在主成分Y2中,X4(叶变色期)和X5(落叶期)载荷值较大,分别为0.808和0.810,而X1(叶芽萌芽期)、X2(展叶初期)、X3(展叶期)的系数较小且为负数,说明第二主成分Y2反映的是植物生长前期的物候特点。
根据叶芽萌芽期(X1)、展叶初期(X2)、展叶期(X3)、叶变色期(X4)和落叶期(X5)5个物候期因子聚类分析(图1,表10)结果,将19个樱花品种无性系分为4类。第1类晚发生物候型为7、11、9、15、10五个,表现为叶芽萌芽晚,展叶初期晚,展叶期晚的物候特征;第2类早发生物候型有4、6、12、18、3、13六个,表现为叶芽萌芽早,展叶初期早,展叶期早的物候特征;第3类早结束物候型有1和2两个,表现为叶变色期和落叶期早;第4类晚结束物候型有8、16、19、5、17、14六个,表现为叶变色期晚、落叶晚。
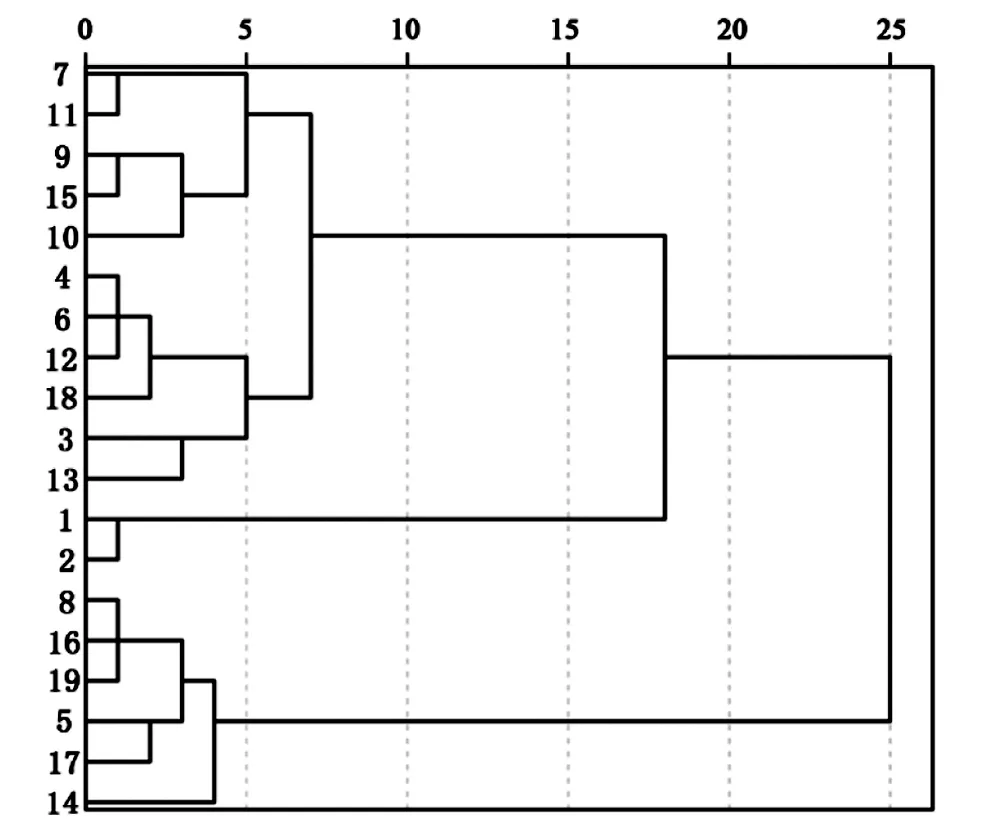
图1 叶物候聚类分析
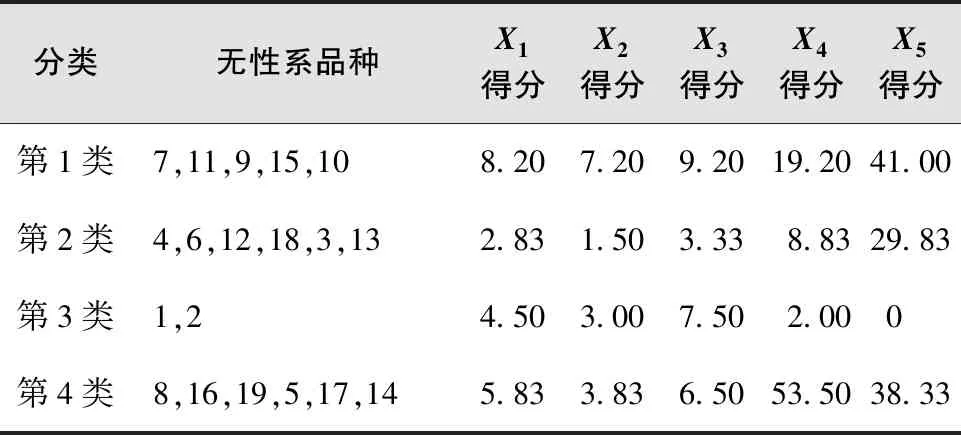
表10 聚类分析的19个无性系物候分类和物候因子得分均值
3.4 生长量月变化情况
19个樱花无性系植株在5月、6月和7月的株高生长量最大,且只有一个生长高峰期,与其他月份有显著差异,其中6号、13号、12号和4号无性系植株年生长量最大,分别达到121.45 cm、120.18 cm、114.35 cm和111.83 cm,8号和14号株高年生长量最小,仅为64.97 cm和55.94 cm(图2)。19个樱花无性系植株的地径月生长情况如图3所示,地径的生长高峰期在5月、6月和11月,一年中有2个生长高峰期。地径年生长量最大的是4号无性系,达到了13.24 mm,其次是19号,为12.76 mm。
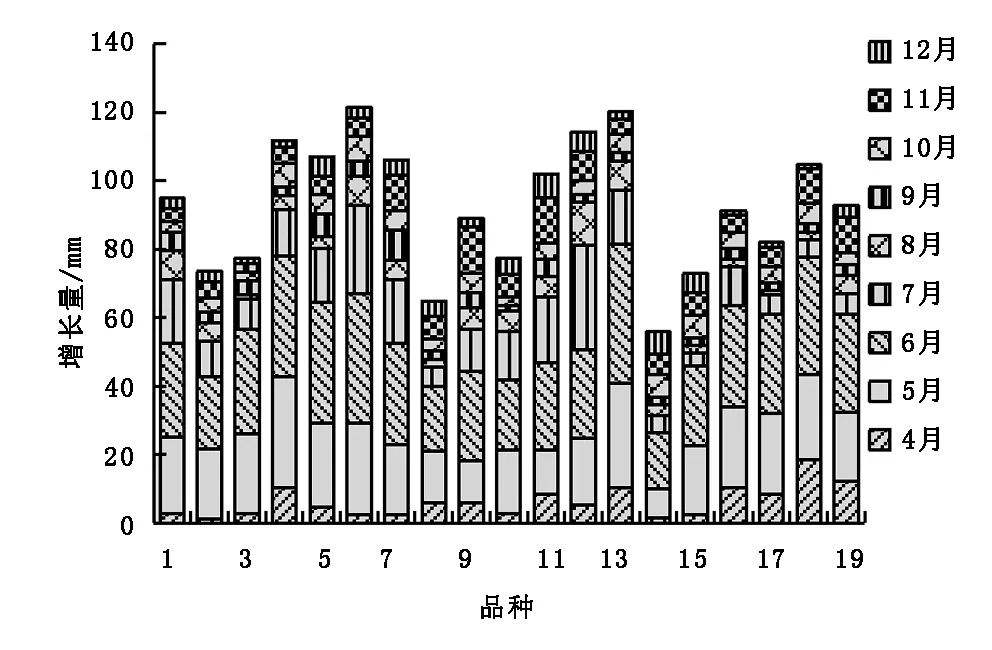
图2 株高月生长Fig.2 Monthly growth histogram of plant height
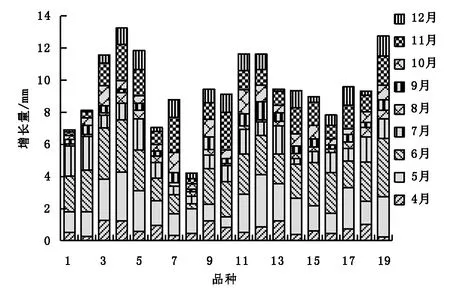
图3 地径月生长Fig.3 Monthly growth histogram of ground diameter
4 结论与讨论
对植物的生长状况进行观测记录,可以最直观地了解植物在这个地区的生长适应性。比较种间苗期生长表现,是选育优良遗传资源的有效途径。本试验通过观察19个樱花无性系在杭州地区的生长适应性、开花习性、物候特点,测量株高地径的各月生长量,发现安徽野樱、霞樱、香味山樱3种在嫁接2年后开花,较其他品种表现出早开花优势。兰兰樱、太白樱、阳春樱3种在杭州地区的6月和9月表现出了较强的抗病虫害能力,安徽野樱、吉野枝垂、绿樱和香味山樱4种的抗病虫害能力较差,容易出现病虫害。另外,夏季日灼病也是樱花常见的现象。在19个樱花无性系中,安徽野樱、吉野枝垂、绿樱、太白樱、阳春樱和越之彼岸樱在夏季出现了不同程度的日灼现象,其他无性系表现良好,日灼现象和葡萄[10]、桃树[11]等的研究发现相似,其原因是当植株长期受高温、干旱、光照等胁迫时,植物细胞自身代谢平衡难以维持,活性氧清除系统活性随之加强,体内酶活性降低甚至是失活,膜系统遭受破坏,组织损伤死亡[12]。
植物表现出来的物候是植物受生物和非生物因子(如环境因子)等共同作用而表现出来的以年为周期的自然现象[13]。通过对物候的研究,可以预测植物的花期[14-15],探索植物生长对环境因子的响应规律[16-17],了解外来植物在本地区的环境生长适应性[18-19],评判其观赏性[20]等。对19个樱花无性系的年物候进行调查分析发现,3月中下旬为萌芽集中期,11月中旬—12月为落叶期,绿叶持续时间240~280 d。以5个叶物候期为标准,对19个樱花无性系进行主成分和聚类分析后发现,用一、二主成分可以包含所有信息,第一主成分Y1反映的是植物生长后期的物候特点,第二主成分Y2反映的是植物生长前期的物候特点。聚类将19个无性系分为4类,结合主成分得分,可以将其分为晚发生物候型、早发生物候型、早结束物候型和晚结束物候型。
树木生长规律是树木遗传特性与环境条件相适应的反映,生物量的积累与植株苗高、地径的相关性极为密切[21]。研究发现,19个樱花无性系的株高生长均呈现“慢—快—慢”的生长趋势,株高速生期在一年中的5—7月,7月后温度升高,过高的温度和湿度不利于植株生长,株高生长减慢。这个生长趋势和前人在香叶树[22]、夏腊梅[23]、阿月浑子[24]等的发现一致。樱花无性系地径的年增长有2个增长高峰期,一个出现在5—6月,另一个在11月,与高山松[25]、油松[26]地径的研究发现不一致。但是李培[27]等研究了红椿地径年增长量后表明,红椿的地径增长有2个高峰期,分别出现在8月和10月,与本研究的发现相似。这可能是因为株高停止迅速增长后,地径生长失去竞争力,获得更多的生长量,从而为来年株高的增长提供基础。生长量速生阶段是苗木管理的关键时期,在田间管理中应抓住这个阶段,为苗木生长创造适宜条件,可以更高效率地提高苗木质量。
猜你喜欢
热带作物学报(2022年6期)2022-07-08 06:45:10
中国果树(2022年1期)2022-04-20 04:57:44
内蒙古林业科技(2021年2期)2021-07-08 07:33:04
生态学报(2020年21期)2020-11-24 09:09:42
山东林业科技(2019年6期)2020-01-02 09:36:30
生态学报(2019年21期)2019-12-18 07:25:30
草原(2018年10期)2018-12-21 12:28:38
西南农业学报(2017年5期)2017-06-23 08:14:35
河北林业科技(2016年5期)2016-11-08 03:12:55
西南农业学报(2016年5期)2016-05-17 05:42:22
