
水貂FGF5 基因多态性及其与毛长性状的关联分析
2020-10-21 00:56刘琳玲宋兴超丛波刘宗岳宋姗姗杨福合王桂武
特产研究 2020年5期
刘琳玲,宋兴超,丛波,刘宗岳,宋姗姗,杨福合,王桂武
(中国农业科学院特产研究所,特种经济动物分子生物学国家重点实验室,农业农村部特种经济动物遗传育种与繁殖重点实验室,吉林 长春 130112)
貂皮是中国“东北三宝”(新三宝:人参、貂皮、鹿茸)之一,素有“皮中之王”的美称。貂皮是珍贵的裘皮,其皮板优良,轻柔结实,毛绒丰厚、细密、平齐,色泽光润,有弹性。针毛和绒毛的长度是影响貂皮毛绒质量的重要因素之一。水貂育种工作中,毛绒品质的选育目标为针毛短、平、齐,绒毛细、密,且针、绒毛比例适中,因此,毛长性状在水貂新品种选育及培育过程中尤为重要。目前,国内饲养量较多的水貂品种主要有红眼白水貂、丹麦咖啡水貂和美国短毛黑水貂等[1]。通过筛选与水貂毛长性状相关的单核苷酸多态性(single nucleotide polymorphism,SNP)位点,运用分子标记辅助选择(marker-assisted selection,MAS)方法可以进一步加快水貂品种选育进展。据研究表明,成纤维细胞生长因子5(fibroblast growth factor 5,FGF5)是成纤维细胞生长因子家族的成员之一。FGF5 基因是通过影响毛囊生长期的长短进而影响被毛长度的调控基因[2-3]。FGF5 基因的隐性突变会导致反常的长毛现象已经在人类[4]及兔[5]、猫[6]、狗[7]、鲸目动物[8]、羊驼[9]和羊[10]等动物上得到验证。丛波等[11]研究表明,美国短毛黑水貂针毛平均长度为(2.02±0.22)cm、绒毛平均长度为(1.69±0.13)cm;丹麦咖啡色水貂针毛平均长度为(2.58±0.21)cm、绒毛平均长度为(2.11±0.29)cm,丹麦咖啡水貂针毛和绒毛都显著长于美国短毛黑水貂。本研究以被毛长度存在显著差异的美国短毛黑水貂和丹麦咖啡水貂为研究对象,利用聚合酶链式反应―限制性片段长度多态性(PCR-RFLP)方法检测FGF5 基因多态性,并对FGF5基因多态性与毛长性状进行相关分析,旨在为今后研究水貂毛绒品质的分子遗传机理与分子标记辅助选择育种提供科学依据。
1 材料与方法
1.1 试验材料
在中国农业科学院特产研究所毛皮动物实验基地和大连名威貂业有限公司采集7 月龄左右、雄性水貂血样,其中,美国短毛黑水貂115 只,丹麦咖啡水貂137只。在水貂取皮期,采集心脏血液,柠檬酸钠抗凝,在﹣ 20 ℃的条件下保存。
1.2 主要试剂
全血基因组试剂盒(北京全式金生物技术有限公司);限制性内切酶 TaqⅠ(赛默飞世尔公司);Marker和Taq DNA 聚合酶(大连TaKaRa 有限公司)。
1.3 DNA 提取
采用全血基因组试剂盒提取血液DNA,利用0.7%琼脂糖凝胶电泳进行检测,紫外分光光度法测定基因组的浓度和纯度,在﹣20 ℃的条件下保存。
1.4 引物合成及 PCR 扩增
以GenBank 数据库中水貂FGF5 基因序列(登录号:JF288167.1)为参考,利用 Primer Premier 5.0 软件在其内含子1 和外显子1 区域设计1 对引物FGF51-R(5′-AGGGCTATCACTAAAGCCGC-3′)和 FGF51-S(5′-TGGGGAGAGGAACCTCGATT-3′),由吉林省库美生物科技有限公司合成。
PCR 反应程序如下:94 ℃ 预变性 5 min;94 ℃ 变性 30 s,60 ℃ 退火 30 s,72 ℃ 延伸 45 s,共 30 个循环;72 ℃延伸5 min;在4 ℃条件下保存。
1.5 SNP 分析
先用30 个美国短毛黑水貂的基因组进行PCR 扩增,扩增产物送交吉林省库美生物科技有限公司进行测序。利用DNAMAN 软件对扩增核苷酸序列进行比对分析并筛查SNP。经分析发现,在扩增产物的 376处存在多态性,该位点位于FGF5 基因内含子1 中。
1.6 PCR-RFLP 分析
利用NEBcutter V 2.0 在线酶切设计软件(http://nc2.neb.com/NEBcutter2/)查询限制性内切酶,发现Taq I 可识别FGF5 基因T376C 位点的C 等位基因。
基因型鉴定:内切酶TaqⅠ识别位点为AGTCGAT,正常情况下将621 bp 的扩增序列酶切为376 bp 和245 bp 两个片段。将能够酶切的基因型定义为CC 基因型,不能酶切的基因型为TT 基因型,杂合子则为CT基因型。
对 115 只美国短毛黑水貂和 137 只丹麦咖啡水貂的基因采用PCR-RFLP检测方法进行FGF5 基因多态性检测。
1.7 数据分析
利用PopGen 32 软件统计基因型频率和等位基因频率、纯合度和杂合度等群体遗传参数。使用SPSS 16.0统计软件进行SNP位点与毛长性状的关联分析,构建线性分析模型,具体模型如下:
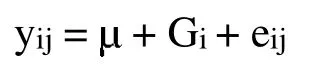
2 结果与分析
2.1 FGF5 基因的 PCR 扩增结果及测序筛选 SNP
利用引物对30 只美国短毛黑水貂的FGF5 基因进行PCR 扩增,扩增产物在500~750 bp 间,与预期结果相一致。从图1 可以看出,SNP 位点位于FGF5 基因内含子1 区域的376 bp 处,经分析处于TaqⅠ内切酶位点内。

图1 CC、TC 和TT 基因型的测序峰图Fig.1 The sequencing peak graph of CC, TC and TT genotypes
2.2 PCR-RFLP 检测分析
PCR 扩增产物长度为621 bp,采用限制性内切酶TaqⅠ对2 个水貂品种FGF5 基因的扩增产物进行酶切,酶切产物经1.5%琼脂糖凝胶电泳检测,结果见图2。由图 2 可知,FGF5 基因 T376C 位点存在 3 种基因型,分别为 TT(621 bp)、TC(621、376 和 245 bp)和 CC(376 和 245 bp)。
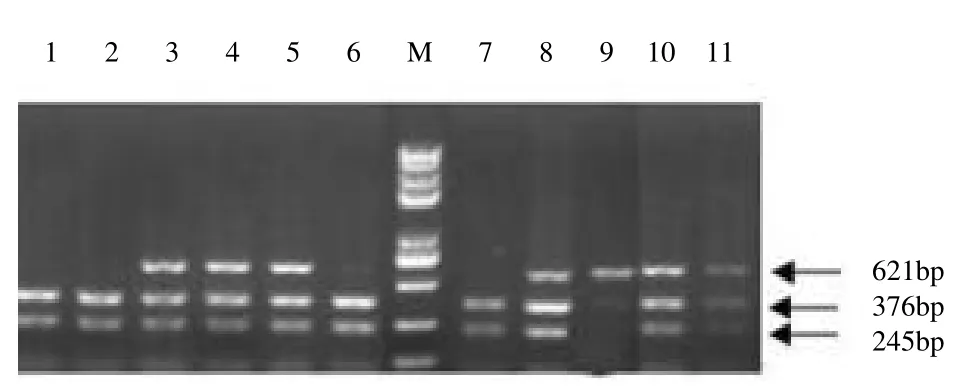
图2 FGF5 基因T376C 位点的酶切分析结果Fig.2 Results of FGF5 T376C locus digested with Ⅰenzyme
2.3 FGF5 基因型频率和等位基因频率分布
由表1 可知,美国短毛黑水貂群体中检测到3 种基因型,以 CC 型居多,TC 型次之,TT 型最少,而 C 等位基因频率高于T 等位基因频率;咖啡水貂群体中以TC 型居多,CC 型次之,TT 型最少,而 C 等位基因频率与T 等位基因频率相差不大。
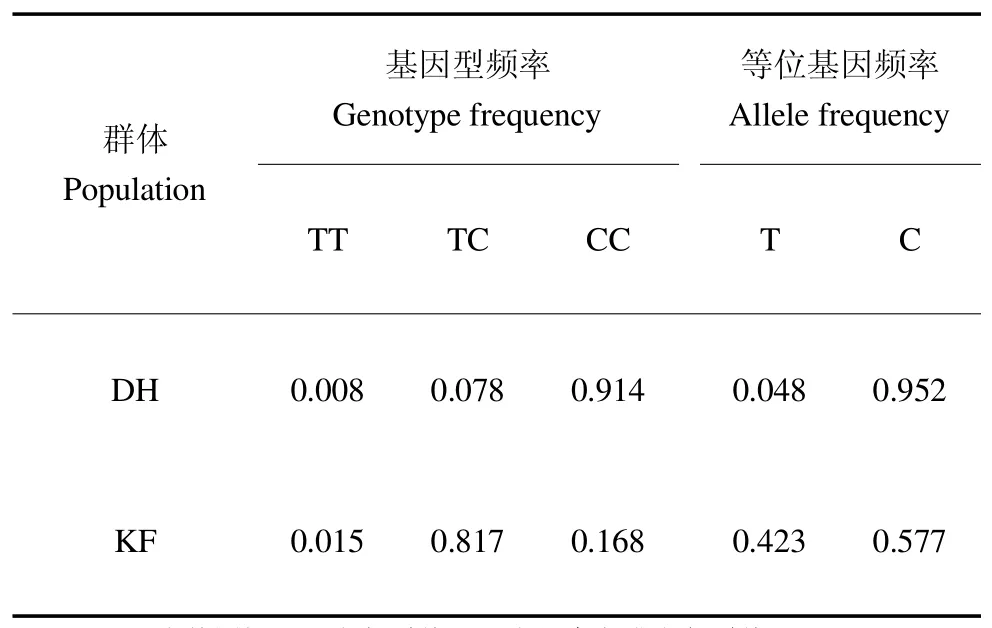
表1 FGF5 基因 T376C 位点的基因型频率和等位基因频率Table 1 Genotype frequency and allele frequency of FGF5 T376C locus
2.4 不同水貂品种 FGF5 基因 T376C 位点的遗传多样性分析
由表2 可知,T376C 多态位点在美国短毛黑水貂群体中为中度多态(0.25 <PIC <0.05),而在丹麦咖啡水貂群体中表现为高度多态(PIC >0.25)。2检测结果表明,T376C 位点在美国短毛黑水貂群体处于Hardy-Weinberg 平衡状态,在丹麦咖啡水貂群体中显著偏离Hardy-Weinberg 平衡状态。

表2 2 个水貂品种 T376C 位点遗传多态性Table 2 Genetic polymorphism of T376C locus in two mink breeds
2.5 FGF5 基因 T376C 位点多态性与水貂毛长性状的关联分析
由表3 可知,T376C 位点形成的不同基因型在两种水貂品种中差异极显著(P <0.000 1),说明该位点可能与水貂毛长性状相关。

表3 T376C 位点基因型与水貂毛长表型的关联分析Table 3 Relationship analysis between the genotype of T376C locus with hair length phenotype of mink
3 讨论
美国短毛黑水貂和丹麦咖啡水貂均为国外引进品种,是目前我国饲养量较多的水貂品种[12]。美国短毛黑水貂毛绒品质优良,针毛短、平、齐,绒毛细、密,分布均匀,皮板富有弹性,但繁殖力较低;丹麦咖啡水貂体型较大,繁殖力较高,被毛略粗糙,呈浅褐色或深褐色[13]。美国短毛黑水貂和丹麦咖啡水貂的毛绒品质差异较大,因此,本研究选择这2 个品种开展毛绒品质相关分子遗传标记研究。
FGF5 是毛囊从生长期到休止期转换的重要的影响因子,同时也被证明 FGF5 是体内诱导退行期毛囊分泌和抑制毛发生长的信号蛋白。目前,关于FGF5 基因突变导致动物毛长表型改变和FGF5 基因多态性的检测方法均有报道[14-16]。本研究分析了FGF5 基因在不同水貂品种中的多态性,不同品种水貂群体遗传学分析表明 T376C 位点在美国短毛黑水貂和丹麦咖啡水貂中的基因型频率有很大的差异。在美国短毛黑水貂群体中以CC 基因型居多,而C 等位基因频率高于T 等位基因频率;在丹麦咖啡水貂群体中以TC 基因型居多,而C 等位基因频率与T 等位基因频率相差不大;经卡方检验发现,品种间的基因型频率差异极显著(P<0.01)。丹麦咖啡水貂针毛和绒毛都显著长于美国短毛黑水貂,据此推测,该位点可能与水貂毛长性状相关。T376C 位点在美国短毛黑水貂群体中为中度多态,而在丹麦咖啡水貂群体中表现为高度多态。T376C 位点在短毛黑水貂群体处于Hardy-Weinberg 平衡状态,与刘琳玲等[17]研究结果基本一致。然而,丹麦咖啡水貂群体在该位点显著偏离Hardy-Weinberg 平衡状态,究其原因可能是因为常规选择育种对该品种水貂绒毛经济性状产生一定的影响,并且本研究样本量较少,后续尚需增加检测数量及扩大水貂品种范围以进一步验证本试验结果。
本研究基于 PCR-RFLP 技术[18]开展了 2 个品种水貂群体T376C 位点的多态性检测及其与毛长性状的相关性分析,结果表明,该位点与水貂毛长性状存在极显著相关。绒山羊FGF5 基因多态性与绒长度相关,苏博美利奴羊FGF5基因多态性与其产毛性状相关,兔的产毛量也被证实与该基因相关[5]。FGF5 基因T376C位点可能是影响水貂毛长的主控位点,为水貂毛长性状分子遗传标记研究提供了一定的分子基础[19]。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
昆明医科大学学报(2022年3期)2022-04-19
昆明医科大学学报(2022年2期)2022-03-29
中国动物检疫(2022年1期)2022-01-13
智慧健康(2021年17期)2021-07-30
昆明医科大学学报(2021年2期)2021-03-29
家畜生态学报(2020年11期)2020-12-04
环球时报(2020-06-08)2020-06-08
新课程·下旬(2018年9期)2018-11-14
青少年科技博览(中学版)(2015年10期)2015-01-11
