
大白菜干烧心病原菌Pantoea ananatis的生物学特性及全基因组序列分析
2020-10-20 06:48张烨张庶李化银王凤德高建伟
山东农业科学 2020年9期
张烨,张庶,李化银,王凤德,高建伟
(1.山东省农业科学院蔬菜花卉研究所/山东省设施蔬菜生物学重点实验室/国家蔬菜改良中心山东分中心,山东 济南 250100;2.黄山学院,安徽 黄山 245041)
大白菜(Brassica rapa L.ssp.pekinensis)原产于中国,是我国的特产蔬菜。大白菜产量高、生产成本低、种植方法简单、耐运输、耐储藏,且味美、营养丰富,深受人们喜爱,在蔬菜产业中具有重要地位。据统计,我国大白菜播种面积每年高达266.7万公顷,产值约600亿元以上,占全国蔬菜播种总面积的15%左右[1]。同时,大白菜是城乡居民的主要食用蔬菜,在保障全国蔬菜均衡供应、平抑菜价和农民增收等方面做出了突出贡献。
自20世纪70年代以来,大白菜干烧心病发生严重,对大白菜产量和品质造成了严重的影响,减少了菜农收入[2,3]。干烧心病又称顶烧病、心腐病和缘腐病,前人在水果和蔬菜上的研究表明,缺钙时其生长点及附近组织的生长会受到影响,表现出顶烧或心腐的症状[4-11]。因此,以往通常认为大白菜干烧心病是一种生理性病害,主要由植株体内生理性缺钙引起,表现为结球前期内部幼叶的边缘呈水渍状、半透明,随后脱水萎蔫,叶色变浅、发黄,顶边干枯,叶肉呈黄白色的干纸状[12];如在结球后期发病,则植株外观正常,但叶球内部的心叶边缘部分变干、黄化、焦枯,出现干纸状的带状病斑或不规则病斑,发病部位与健康部位界限较为清晰。大白菜干烧心病多发生在莲座期或包心期,但当生长环境不适宜或者该品种对干烧心病的抗性较弱时,苗期也会发生干烧心病[12];另外,贮藏期间,由于生理活动仍缓慢进行且钙元素供应停止,大白菜干烧心病可持续发展,并且在贮藏条件不好的情况下大白菜植株极易受病菌侵染,使叶球出现“夹叶烂”的现象,最终导致整个植株腐烂。
根据传统描述的大白菜干烧心病症特点,本课题组经多年田间调查发现,除缺钙引起的大白菜干烧心病外,细菌感染也会导致大白菜干烧心[13],因此,将其界定为细菌性干烧心和缺素(钙元素等)性干烧心两种类型。早期细菌性干烧心病叶边缘呈湿腐状,而缺钙引起的干烧心病叶边缘则成干枯状,这是两种类型大白菜干烧心病的主要区别。目前,对大白菜干烧心病的研究主要集中于生理性缺钙引起的病害,而对细菌性干烧心病研究较少;且人们对两种类型的干烧心病往往区分不清,仅采用常规田间栽培管理措施,难以实现对大白菜干烧心病的有效防治。
2019年,我们首次报道了泛菌Pantoea ananatis能导致大白菜细菌性干烧心病的发生[13]。本研究即对分离得到的P.ananatis Y2菌株进行生物学特征及其引起的大白菜干烧心病发病规律调查,并对P.ananatis的全基因组进行测序,分析基因组特征。本研究将有助于人们全面揭示大白菜干烧心病发生的分子机理,对利用现代生物技术手段结合传统育种方式培育大白菜高产、优质、抗病品种具有重要意义。
1 材料与方法
1.1 试验材料
本试验在山东省农业科学院蔬菜花卉研究所试验基地进行,所用品种为春大王。将大白菜种子在仅含有蒸馏水的培养皿上萌发后转入基质(营养土∶蛭石=2∶1)中,在25℃/22℃、14 h/10 h(光/暗)以及相对湿度50%的条件下培养至完全展开第七片叶,用于细菌接种。
1.2 病原菌分离
从田间取回大白菜感病植株,用刀片在大白菜叶片病健交界处切取1 cm2左右的方块,在超净台进行组织表面消毒后放入无菌三角瓶中,加入70%乙醇消毒30 s,期间不断摇晃三角瓶;倒出70%乙醇后,再加入1%次氯酸钠(现用现配)消毒5 min;用灭菌蒸馏水冲洗5遍后,用灭菌滤纸吸干多余水分,并转移至无菌研钵中。向研钵中加入1 mL灭菌水,用研磨棒将组织研磨充分,静置10 min左右,使组织中的细菌进入水中,形成悬浮液。将细菌悬浮液分别进行10、100、1 000、10 000倍稀释,分别取10μL涂布在NB培养基上(pH 7.0,含1.5%琼脂),30℃倒置培养2~3 d,对平板上出现的主要菌落进行纯化。
1.3 病原菌P.ananatis致病性的测定
在NB液体培养基中接种纯化的菌落,30℃条件下振荡培养过夜,常温下5 000 r/min离心10 min后,用灭菌蒸馏水重悬菌体,并调整菌液浓度为1×106cfu/mL。用1 mL无针头无菌注射器吸取重悬菌液并注射进大白菜叶片背面,以无菌蒸馏水作为对照。设置3次重复,每个重复处理10棵大白菜幼苗。接种后,观察大白菜叶片的发病情况,完成柯赫氏法则的验证,对分离得到的致病菌再次确认。
1.4 病原菌P.ananatis的生物学特性
1.4.1 形态学观察 参照《常见细菌系统鉴定手册》进行革兰氏染色,使用光学显微镜观察菌体形态。
1.4.2 生理生化鉴定 依照《伯杰氏系统细菌学手册》(Bergey’sManual of Detenninative Bacteriology)对P.ananatis进行生理生化鉴定,包括革兰氏染色、糖发酵试验、氧化酶反应、V-P试验、柠檬酸盐利用试验。
1.5 P.ananatis基因组测序分析
1.5.1 DNA提取 采用天根生化科技(北京)有限公司的细菌基因组DNA提取试剂盒(DP302)进行。
1.5.2 测序文库构建及测序 提取的细菌DNA使用琼脂糖凝胶电泳检测,使用Qubit进行定量。使用Covaris超声破碎仪从DNA上随机打断成长度约为350 bp的片段,并对该片段进行末端修复、加A、加测序接头、纯合与PCR扩增等完成文库制备。使用Illumina NovaSeq PE150进行测序。
1.5.3 生物信息学分析 (1)原始数据处理:使用readfq(Version 10)对原始数据进行过滤,包括去除含低质量碱基的reads、含N碱基超过10 bp的reads、含Adapter的reads、可能来源于宿主的reads和duplication污染。
(2)样品组装:使用SOAP denovo、SPAdes和AbySS对预处理后的CleanData进行组装[14-17],使用CISA[18]软件进行整合,初步组装结果使用gapclose(Version1.12)进行优化和补洞,并过滤掉500 bp以下的片段。
(3)基因组组分分析:使用GeneMarkS(Version 4.17)软件对基因组编码基因预测[19]。使用RepeatMasker(Version open-4.0.5)软件进行散在重复序列预测[20],使用TRF(Tandem Repeats Finder,Version 4.07b)搜寻DNA序列中的串联重复序列[21]。通过tRNAscan-SE软件(Version 1.3.1)对tRNA进行预测[22],通过与近缘参考序列的rRNA库与rRNAmmer软件(Version 1.2)预测rRNA[23],使用Rfam database对sRNA进行注释[24,25],使用cmsearch程序(Version 1.1rc4)确定最终的sRNA。使用IslandPath-DIOMB软件(Version 0.2)预测基因岛[26]。使用phiSpy软件(Version 2.3)预测样品基因组上的前噬菌体[27]。使用CRISPRdigger(Version 1.0)对样品基因组进行CRISPR预测[28]。
(4)功能注释:使用通用功能数据库与特定功能数据库进行注释。通用功能数据库包括GeneOntology数据库[29]、KEGG(Kyoto Encyclopedia of Genes and Genomes)数据库[30,31]和COG(Cluster of Orthologous Groups of Proteins)数据库[32];特定功能数据库包括TCDB(Transporter Classification Database)数据库[33]、碳水化合物活性酶(Carbohydrate-Active EnZymes Database,CAZy)数据库[34]和病原与宿主互作数据库等。
2 结果与分析
2.1 P.ananatis的致病性鉴定
2018年6月,在山东省农业科学院蔬菜花卉研究所试验基地对大白菜进行春季调查时发现了由P.ananatis引起的细菌性干烧心症状(图1A),与生理性缺钙引起的大白菜干烧心症状(图1B)明显不同。叶边缘腐烂还是干枯是细菌性与生理性缺钙引起的大白菜干烧心病的主要区别。
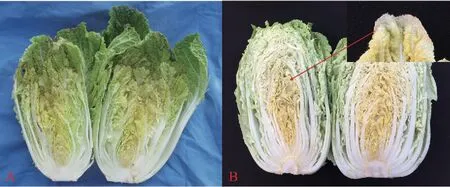
图1 Pantoea ananatis与生理性缺钙引起的大白菜干烧心病症状对比
将分离、纯化得到的P.ananatis Y2菌株回接至健康的大白菜叶片(以接无菌水为对照),10天后从回接部位发现坏死病灶(图2);25天后,发病症状与田间自然条件下发病时一致,再次分离纯化得到的细菌菌株形态学特征与接种细菌一致。

图2 病原菌回接10天大白菜病斑
2.2 P.ananatis的生物学特性
2.2.1 形态学特征 P.ananatis呈圆形菌落,菌落直径0.2~0.3 mm,表面较为光滑,边缘整齐,培养初期为乳白色,后期变为深黄色(图3)。
2.2.2 生理生化特性 P.ananatis革兰氏染色阴性,单个菌体杆状,菌体大小(0.92~2.87)μm×(0.64~1.25)μm。能利用D-葡萄糖、α-乳糖、蔗糖、D-果糖和L-鼠李糖产酸但不产气,不能利用麦芽糖产酸产气,V-P试验和柠檬酸利用试验结果为阳性,氧化酶试验结果为阴性(表1)。参照《常见细菌系统鉴定手册》与《伯杰细菌手册》,该病原菌与泛菌属细菌的特征极为相似。
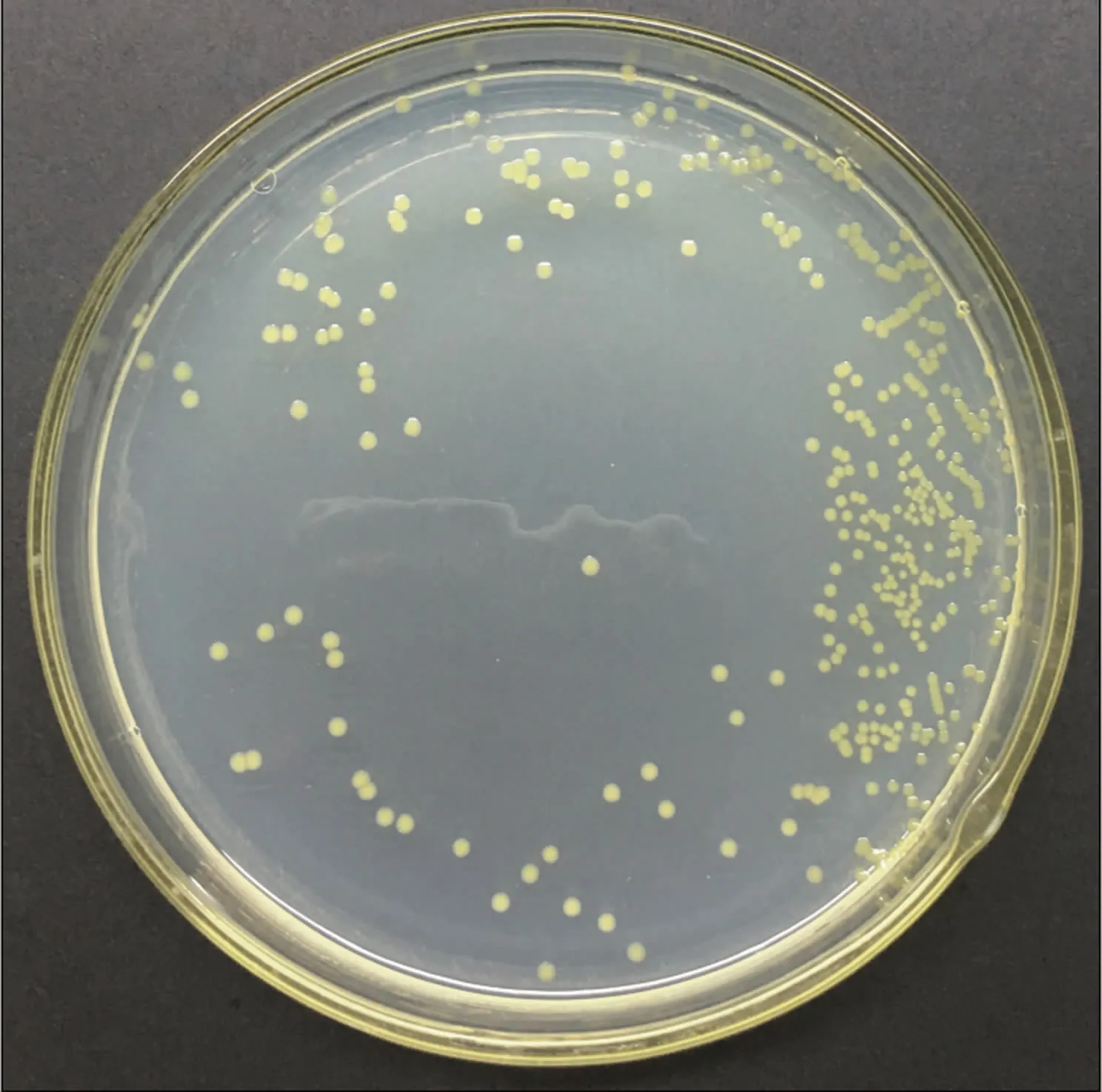
图3 细菌性大白菜干烧心病病原菌的菌落培养形态

表1 Pantoea ananatis的生理生化特性鉴定结果
2.3 P.ananatis的全基因组测序分析
2.3.1 数据处理与样品组装 使用Illumina NovaSeq PE150测序平台对P.ananatis Y2进行了全基因组测序,获得了658 Mb原始数据,其中过滤后的有效数据为600 Mb,占原数据量的91.19%。有效数据的Q20达到97.75%,Q30达到93.59%,预测基因组大小为5.26 Mb(表2)。
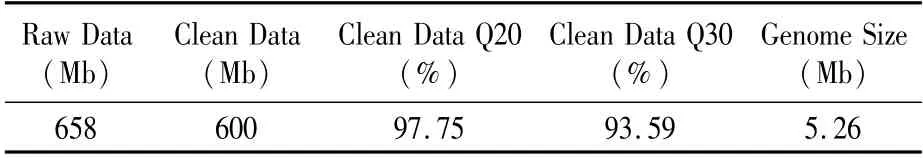
表2 基因组测序数据统计
获得的有效数据经SOAP denovo(Version 2.04)、SPAdes和AbySS软件组装后,使用CISA软件进行整合,使用gapclose(Version 1.12)等软件进行优化后,组装成32个contig(>500 bp),总长度为5 062 658 bp,G+C含量为53.54%,其中长度最长的contig包含1 350 885 bp序列,长度最短的contig包含562 bp序列(表3)。

表3 基因组组装结果contig统计
2.3.2 编码基因预测结果 对编码基因进行预测,结果(表4)显示,预测编码基因4 955个,基因总长度4 380 990 bp,平均基因长度884 bp,在全基因组序列中编码基因占86.54%。编码基因的长度从小于100 bp到大于2 000 bp均有分布,其中基因长度200~300、400~500、700~800、900~1 000 bp范围内分布的编码基因数目较多,而基因长度在0~100、1 700~1 800、1 800~1 900、1 900~2 000 bp的编码基因数目较少(图4)。

表4 编码基因预测结果

图4 P.ananatis Y2基因长度分布
2.3.3 重复序列统计结果 散在重复序列、串联重复序列是根据重复序列在基因组上的分布确定的。在P.ananatis Y2中共有109个散在重复序列,包括:长末端重复序列55个,DNA转座子15个,长散在重复序列25个和短散在重复序列14个,这些散在重复序列长度范围901~3 742 bp,在基因组中所占比例从0.0178%到0.0739%,平均长度范围64~86 bp(表5)。
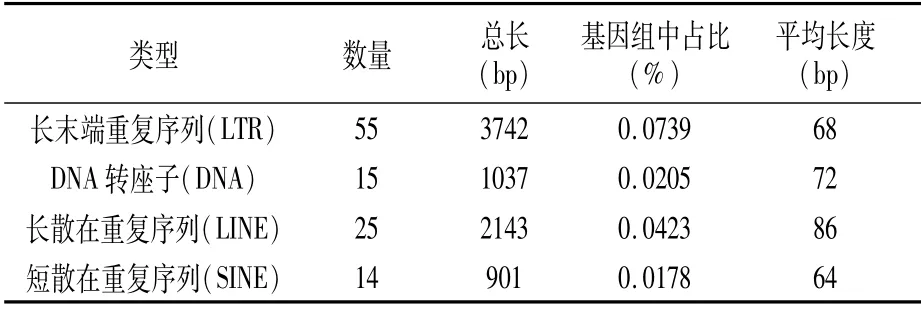
表5 散在重复序列统计结果
在P.ananatis Y2中共鉴定到串联重复序列223个,包括:串联重复序列135个、小卫星DNA 81个和微卫星DNA 7个,串联重复序列总长度范围是486~17 135 bp,在基因组中所占比例为0.0096%~0.3385%(表6)。
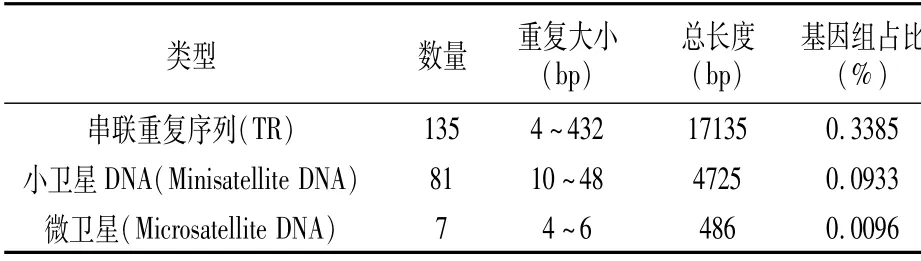
表6 串联重复序列统计结果
2.3.4 非编码RNA(ncRNA)统计结果 对于微生物而言,非编码RNA中研究较为普遍的包括sRNA、rRNA、tRNA等。在P.ananatis Y2中,共鉴定到ncRNA 111个,其中,tRNA数目最多,鉴定到71个,sRNA次之,鉴定到32个,rRNA(5S)鉴定到6个,rRNA(16S)和rRNA(23S)数目最少,各鉴定到1个(表7)。这些ncRNA的平均长度从78 bp到2 549 bp,总长度从660 bp到5 572 bp(表7)。
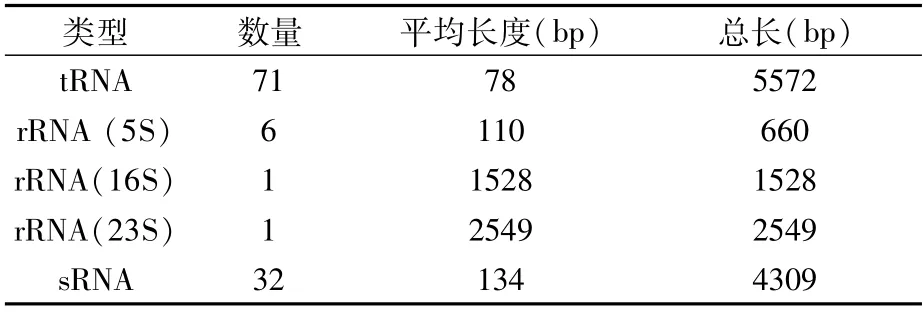
表7 ncRNA去冗余后的统计结果
2.3.5 基因岛、前噬菌体和CRISPR统计结果
在P.ananatis Y2中,鉴定到15个基因岛(GIs),其总长度为185 078 bp,平均长度为12 339 bp(表8)。对基因岛基因分布进行统计,其中长度小于15 kb的基因岛中的基因分布见图5。在P.ananatis Y2中,鉴定到6个前噬菌体,总长度为169 319 bp,平均长度为28 219 bp(表8)。在P.ananatis Y2中,仅鉴定到1个CRISPR,长度为95 bp(表8)。
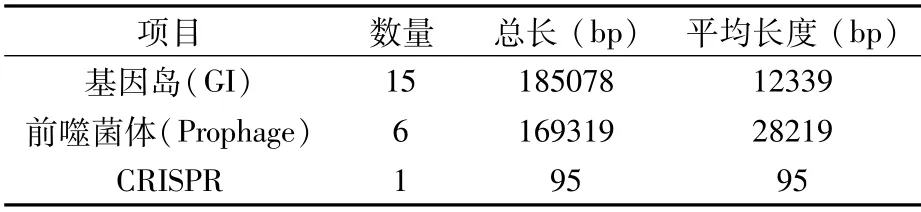
表8 基因岛、前噬菌体和CRISPR预测结果统计
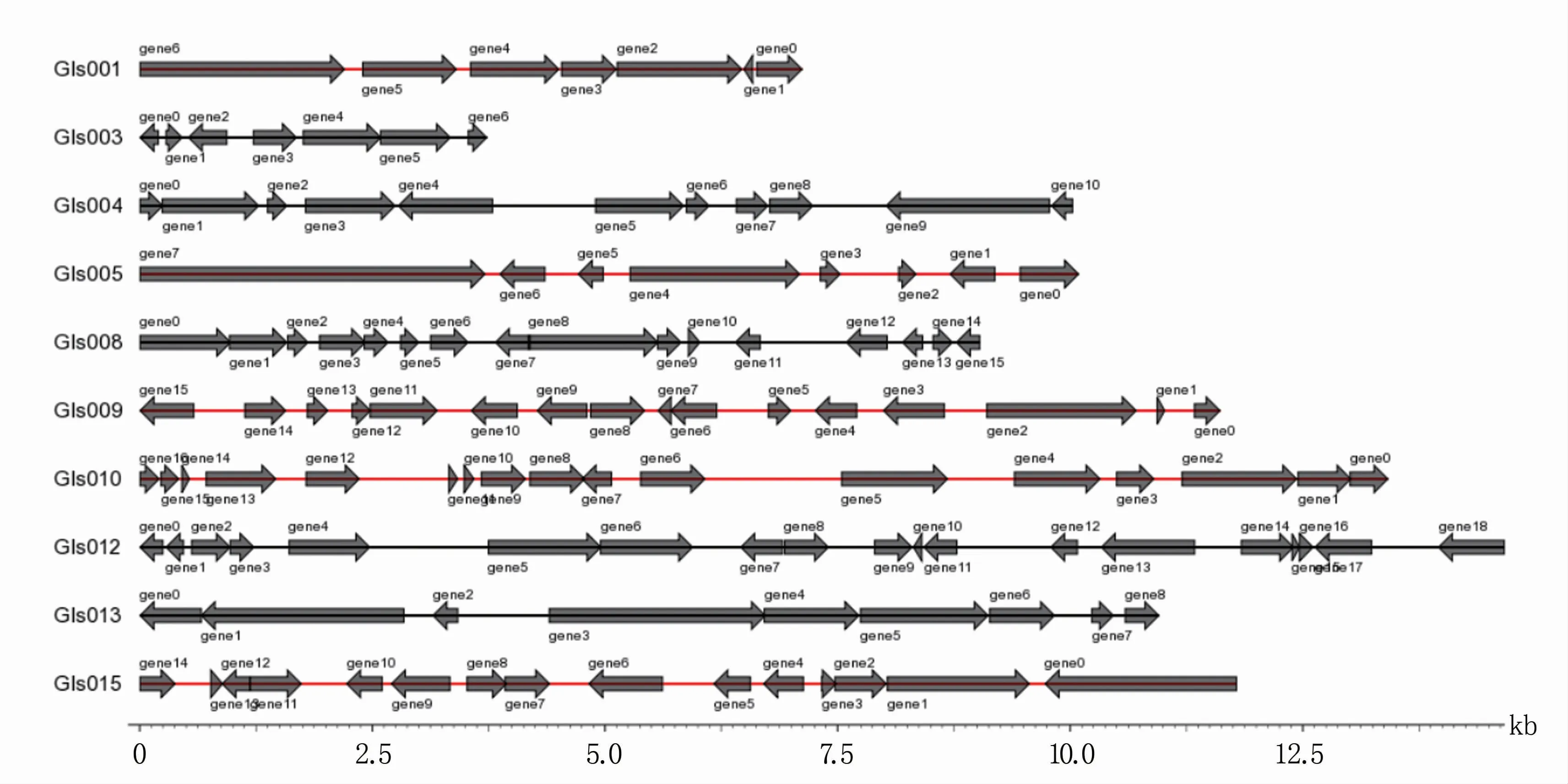
图5 小于15 kb的P.ananatis Y2基因岛中的基因分布
2.4 P.ananatis的基因组功能注释
2.4.1 GO功能注释 P.ananatis Y2鉴定到的编码基因的分子功能可以分类为11个条目,其中富集基因数目最多的两个条目是binding和catalytic activity(图6)。依细胞组分分类,编码基因可富集到10个条目中,cell和cell part富集的基因数目最多(图6)。在生物过程类别中,编码基因富集在24个条目中,富集基因数目最多的条目是cellular process和metabolic process(图6)。
2.4.2 KEGG注释 从P.ananatis Y2鉴定到的编码基因在多个KEGG条目中均有富集,在cellular processes类别中,富集基因数目最多的两个条目是cellular community-prokaryotes和cell motility;在environmental information processing类别中,都富集在两个条目,分别是signal transduction和membrane transport;在genetic information processing类别中,编码基因被富集到4个条目中,富集基因数目最多的条目是translation;在human diseases类别中,编码基因被富集到11个条目中,基因数目最多的两个条目是drug resistance:antimicrobial和infectious diseases:bacterial;编码基因在metabolism类别中富集的条目最多,有12个条目,富集基因数目最多的是global and overview maps,其 次 是carbohydrate metabolism 和amino acid metabolism。在organismal systems分类中,编码基因被富集为7个条目,包括endocrine system、environmental adaptation、immune system 和aging等(图7)。
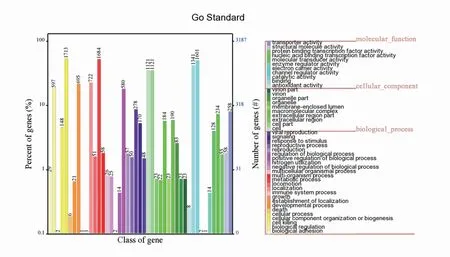
图6 P.ananatis Y2基因功能注释GO功能分类
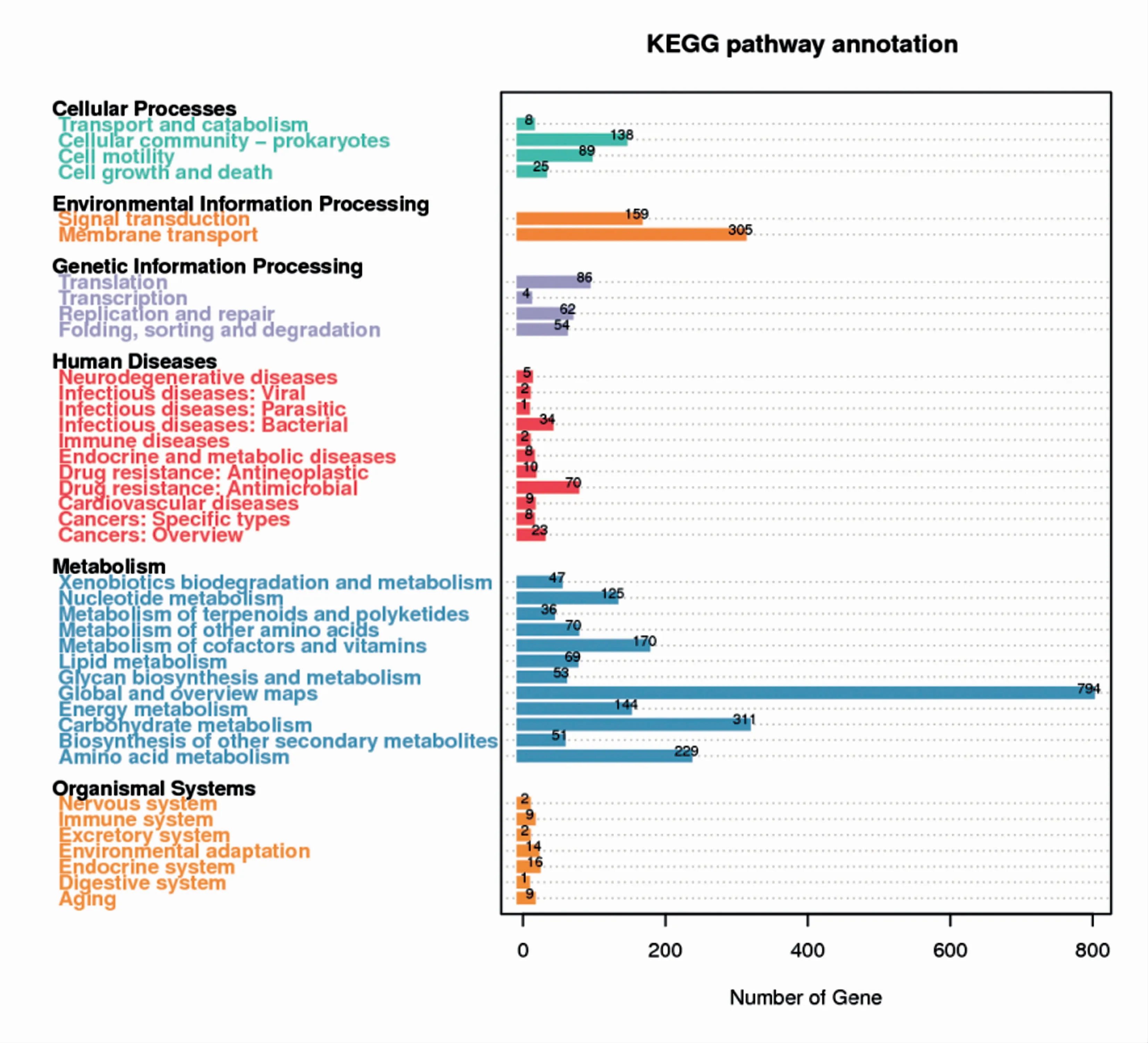
图7 P.ananatis Y2基因功能注释KEGG代谢通路分类
2.4.3 COG注释 P.ananatis Y2编码基因在COG数据库按照功能一共可以分为23类,其中富集基因数目最多的4个条目分别是:carbohydrate transport and metabolism、amino acid transport and metabolism、general function prediction only和transcription(图8)。

图8 P.ananatis Y2基因功能注释COG功能分类
2.4.4 TCDB注释 TCDB转运蛋白分类数据库是膜转运蛋白包括离子通道的分类系统,以5个级别进行分类。P.ananatis Y2编码基因在TCDB转运蛋白分类数据库中的第一级分类共富集在7个条目中,富集基因数目最多的条目是primary active transporters,其次是electrochemical protential-driven transporters(图9)。
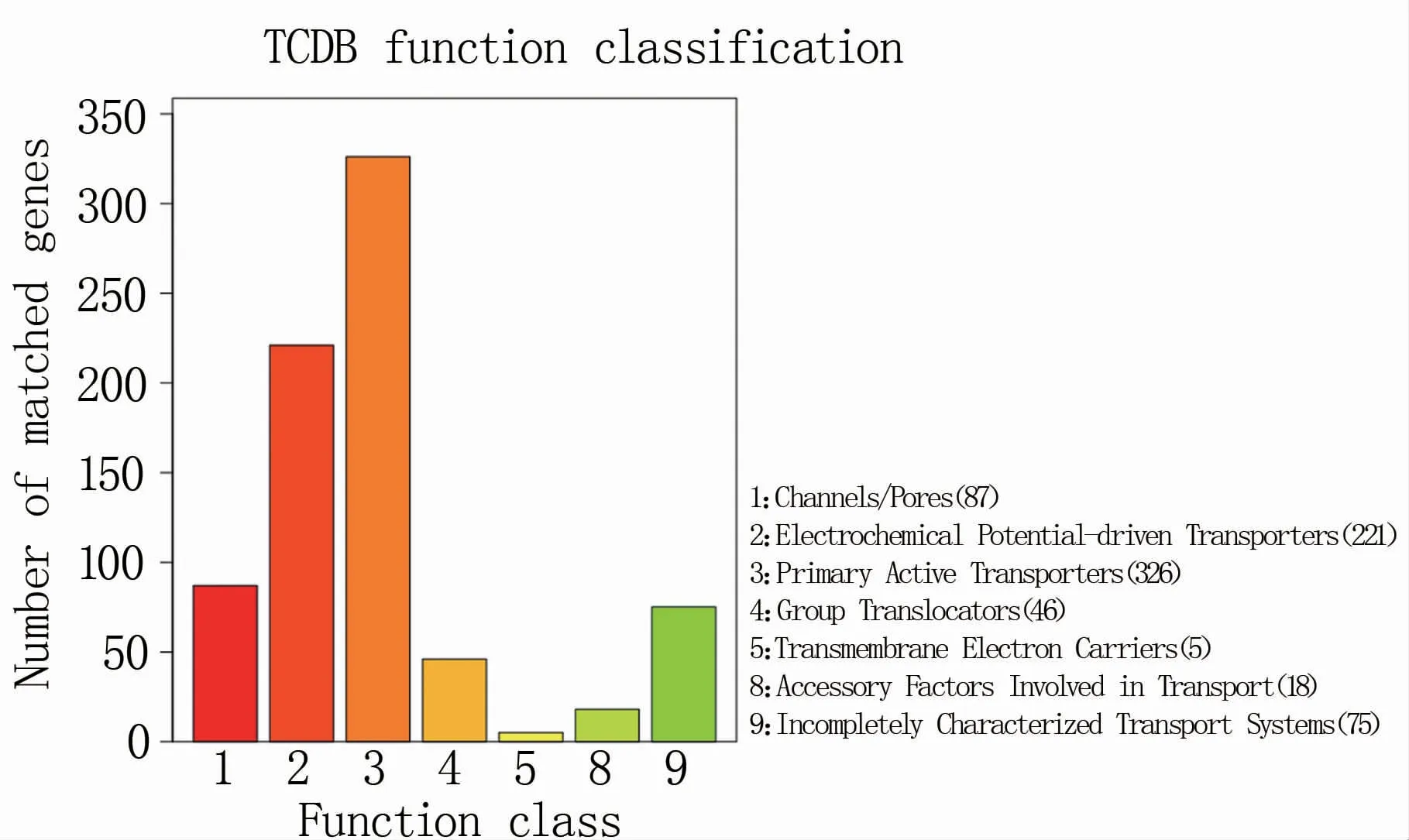
图9 P.ananatis Y2基因功能注释TCDB功能分类
2.4.5 分泌蛋白预测 分泌蛋白是指在细胞内合成的可在信号肽引导下穿过细胞膜分泌到细胞外起作用的蛋白质。分泌蛋白中有许多是生命活动所需的重要酶类。分泌蛋白的N端是由15~30个氨基酸组成的信号肽,对分泌蛋白的分泌起主导作用。使用SignalP(Version 4.1)和TMHMM(Version 2.0c)可对信号肽和跨膜结构进行预测,以确认预测蛋白序列是否属于分泌蛋白。P.ananatis Y2编码基因中有386个蛋白含有信号肽,有1 147个蛋白含有跨膜结构,综合预测为分泌蛋白的数目为311个(表9)。
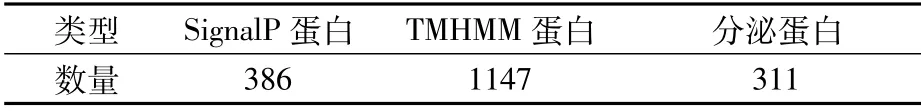
表9 分泌蛋白预测结果
2.4.6 TNSS效应蛋白预测 TNSS(type N secretion systems)即N型分泌系统,目前确定的有7种,为Ⅰ~Ⅶ型。病原菌通过将TNSS效应蛋白分泌至胞外或宿主细胞中控制免疫应答反应及细胞衰亡,从而引起病理反应。其中,T3SS(Ⅲ型分泌系统)通常用来从分子水平研究革兰氏阴性菌的感染机制和毒力作用等,是研究得比较多的分泌系统。P.ananatis Y2的编码基因只有Ⅵ型分泌系统效应蛋白,数量为3个(表10)。而采用EffectiveT3软件(Version 1.0.1)预测其T3SS效应蛋白,发现在4 955个编码基因中,T3SS效应蛋白数目为204个(表10)。两种结果差异的具体原因还有待进一步研究。原菌与宿主互作数据库,病原菌主要为真菌、卵菌和细菌,感染的宿主包括动物、植物、真菌以及昆虫。对P.ananatis Y2编码基因在PHI数据库中匹配的基因数目进行统计,结果(图11)表明,编码基因在PHI数据库中鉴定到29个基因分布在increased virulence(hypervirulence)分类中,319个基因分布于ruduced virulence分类中,6个基因分布于lethal分类中,未见其它致病相关分类含有P.ananatis Y2的编码基因。

表10 TNSS和T3SS结果统计
2.4.7 次级代谢基因簇分析 次级代谢产物是微生物在一定的生长时期以初级代谢产物为前体合成的,对微生物的生命活动无明确功能,并非生长繁殖所必需的物质。采用antiSMASH 程序(Version 2.0.2)对P.ananatis Y2的次级代谢基因簇进行鉴定,结果表明,在P.ananatis Y2中有7条次级代谢基因簇,被分成了5类,其中siderophore和hserlactone各有2条,arylpolyene、thiopeptide和terpene各有1条(图10)。
2.4.8 病原菌与宿主互作注释分析 PHI是病
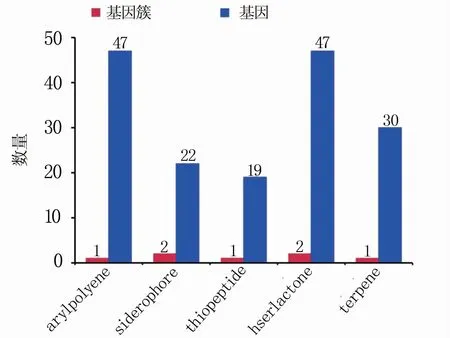
图10 P.ananatis Y2次级代谢基因簇及相应基因数量
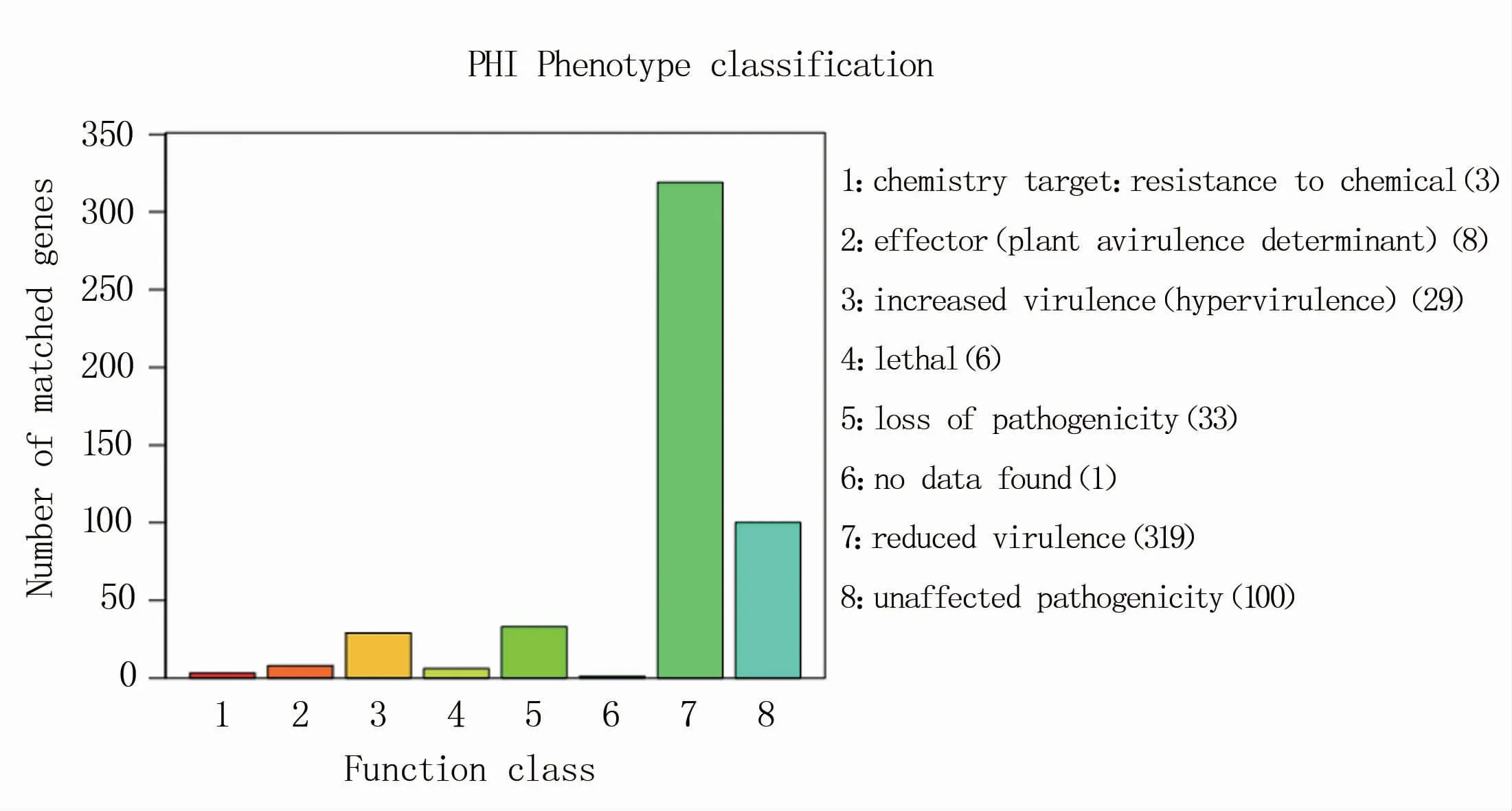
图11 P.ananatis Y2病原体PHI数据库基因分布
3 讨论与结论
2018年6月,在山东大白菜种植地区发生大面积疑似干烧心感病现象,即顶叶边缘初期水浸状,半透明,后变淡黄褐色,皱缩干枯成干边;但持续观察发现到发病后期与生理性干烧心病害的症状并不相同,没有出现叶面变干黄化或心叶边缘枯焦等现象[3],而是叶片边缘严重腐烂,因此,怀疑其并非生理性病害,可能受病原菌感染所致。我们从感病叶片的病健交界处取样,通过分子生物学鉴定,确定该病病原菌为泛菌属的P.ananatis[13]。
泛菌属细菌广泛存在于土壤、水、种子以及植物表面,甚至人与动物的尿液、汗液中也检测出此类细菌。目前,很多学者对此属细菌的致病性仍存在争议,认为泛菌属细菌是非植物致病菌,仅是植物病灶中的次生菌或腐生性细菌[35]。但越来越多的研究发现泛菌属细菌可引起多种植物病害,如P.stewartii可引起玉米细菌性枯萎病,鉴于其危害性及传播性极强,已被列为我国植物检疫对象;P.agglomerans能引起玉米枯萎病、叶疫病[36]以及玉米茎腐病[37]。近年来,国外大量报道指出多种植物的细菌性病害由P.ananatis引起,如P.ananatis可引起玉米细菌性褐腐病以及加州高粱叶斑病[38,39],并且发病面积和危害程度不断加重,引起国外大量学者的广泛关注[40-43]。然而,P.ananatis在我国引起病害的报道较少,其引起大白菜叶部腐烂尚属首次报道[13]。
本研究对从大白菜中分离鉴定到的细菌性干烧心病原菌P.ananatis Y2菌株进行生理生化鉴定和基因组测序分析,获得的组装基因组大小约为5.26 Mb,拼接获得32个contig,编码4 955个基因。将P.ananatis Y2菌株的基因组与已知的SGAir0210和Strain 1.38基因组进行比对(表11),发现其比已知的SGAir0210和Strain 1.38的基因组更大,编码的基因数也更多,但G+C含量差异不大[44,45]。P.ananatis SGAir0210是从空气中分离得到的,P.ananatis Strain 1.38是从水稻根际土壤中分离得到的,未见这两个菌株具有致病性的报道;而本试验分离得到的P.ananatis Y2菌株具有植物致病性,基因组大小的差异可能是导致其具有致病性的原因。
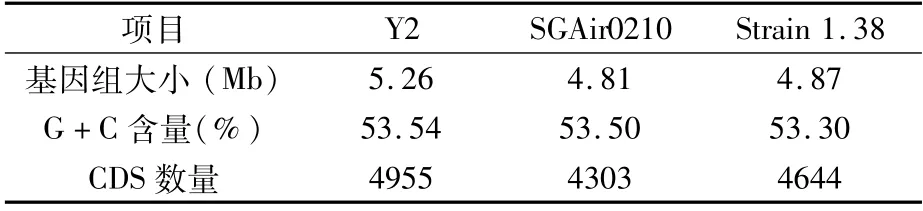
表11 不同Pantoea ananatis菌株基因组比较
在P.ananatis Y2菌株中鉴定到109个重复序列、223个串联重复序列、111个非编码RNA、5个基因岛、6个前噬菌体和1个CRISPR。在GO数据库中,P.ananatis Y2的编码基因在分子功能分类中富集到11个条目中,在细胞组分分类中富集到10个条目中,在生物过程分类中富集在24个条目中。在KEGG数据库中,P.ananatis Y2的蛋白富集到6大分类的40条途径中。在COG数据库中,P.ananatis Y2的蛋白在23个分类中有富集。同时对P.ananatis Y2菌株在膜转运蛋白、碳水化合物活性酶、分泌蛋白、分泌系统蛋白及T3SS效应蛋白、次级代谢基因簇及病原菌与宿主互作等特定功能进行了注释。尽管在P.ananatis Y2中鉴定到29个强致病相关基因,但是目前尚未见到其在大白菜叶片致病机理方面的研究,因而这29个基因对其致病力的影响还需进一步深入研究,这将有利于今后对大白菜叶片细菌性干烧心致病机理及大白菜对其抗性的研究。
猜你喜欢
昆明医科大学学报(2022年3期)2022-04-19
今日农业(2021年19期)2022-01-12
护士进修杂志(2021年22期)2021-11-23
中老年保健(2021年8期)2021-08-24
今日农业(2020年22期)2020-12-25
中国果业信息(2020年5期)2020-12-16
神州·下旬刊(2019年1期)2019-02-11
妇女之友(2015年10期)2016-04-15
阅读与作文(小学高年级版)(2016年3期)2016-03-08
小说月刊(2015年9期)2015-04-23
