
长江中游干流鱼类群落构建机制分析
2020-10-15 09:44杨婷越刘焕章
水生生物学报 2020年5期
杨婷越 俞 丹 高 欣 刘焕章
(1. 中国科学院水生生物研究所水生生物多样性与保护重点实验室,武汉 430072; 2. 中国科学院大学,北京 100049)
群落构成机制(Community assembly mechanism)一直是生态学研究的热点问题之一, 它对于解释物种共存(Species coexistence)至关重要[1]。近年来, 有关群落构成机制的多种理论被提出[2—4], 而现代生态学则将群落的构成机制划分为3种类型:中性作用、竞争作用以及环境过滤作用[5,6]。Hubbell[7]提出的中性作用认为群落中的物种在个体水平上是等价的, 物种间不表现为规律性的关联。竞争作用认为群落的资源有限, 物种间强烈的竞争使得生态位发生分化, 群落中的物种得以共存[8,9]。环境过滤作用认为由于局域生境环境条件的独特性, 使仅有适应该条件的物种才能共存[5,10]。
近年来, 随着系统发育群落结构(Phylogenetic community structure)分析被引入群落生态学, 我们可以有效地检验群落构成和生物多样性维持的各种理论假说。进化和生态相结合的群落系统发育结构(Phylogenetic community structure)研究, 可以从进化角度深入地分析群落物种组成现状, 为有效推测影响群落物种组成的不同生态学机制提供了实验解决方法[11]。该方法认为, 在功能性状保守的情况下, 若群落中生境筛选作用占据主导地位,那么相同生境将筛选出适应能力相似的近缘物种;相反, 竞争作用使生态相似的物种无法共存于同一环境中, 群落内多为亲缘关系较远的远缘物种[12]。但是, 在众多的群落构建机制研究中, 不同尺度的分析得到了不同的结果[13—15], 因此亟待多尺度的研究探讨。
长江流域面积广阔, 生境复杂多样, 鱼类物种多样性丰富, 是世界上著名的生物多样性热点地区,具有极其重要的保护和研究价值, 据不完全统计,长江流域分布有鱼类物种数400余种, 其中局限分布于长江上游的特有鱼类多达124种[16,17]。宜昌至鄱阳湖湖口为长江中游地区, 全长约955 km, 流域面积约为68×104km2[18,19]。长江中游流域底质结构非常多变, 河道蜿蜒曲折, 江心多有沙洲, 流态极为复杂, 是我国多种鱼类尤其是洄游性鱼类优越的栖息与繁殖场所[20]。近年来, 随着捕捞强度的增加,工业及生活污水的排放, 水利工程的建设等人为活动使得长江流域渔业资源、鱼类群落结构发生了一系列的显著变化[21]。本文采用系统发育群落结构的方法对长江中游地区鱼类群落结构及其构建机制进行研究, 以期为长江中游鱼类生物多样性及资源保护提供科学依据。
1 材料与方法
1.1 研究区域
本研究在长江中游共设立5个采样点(图 1)。其中2018年7—8月对枝江、荆州、汉南、湖口4个采样点进行鱼类资源调查, 11—12月对宜昌采样点进行调查。调查方式为自主捕捞或者雇请渔民进行捕捞, 捕捞工具包括定置刺网、流刺网、地笼等,同时走访采样点附近周边水产市场补充部分日常采样中无法采集到的种类。现场进行物种鉴定并记录各采样点鱼类组成数据。物种鉴定参照《中国动物志·硬骨鱼纲·鲤形目(中卷)》、《中国动物志·硬骨鱼纲·鲤形目(下卷)》、《中国动物志·硬骨鱼纲·鲇形目》、《中国动物志·硬骨鱼纲·鲟形目海鲢目鲱形目鼠鱚目》、《中国动物志·硬骨鱼纲·鲈形目(五)·虾虎鱼亚目》和《中国内陆鱼类物种与分布》[22—27]。个体较小的鱼类标本整条采用95%酒精直接固定, 个体较大的标本剪取适量鱼体背部肌肉或鳍条用95%酒精固定, 残体用10%福尔马林固定, 所有标本保存于中国科学院水生生物研究所水生生物博物馆。
1.2 鱼类基因组DNA的提取、扩增及测序
本研究采取标准的高盐抽提法提取鱼类基因组DNA。Cytb基因的扩增引物为: L14724(5′-GACTTGAAAAACCACCGTTG-3′), H15915(5′-CTCCGATCTCCGGATTACAAGAC-3′)[28]。PCR扩增体系总体积为30 μL, 其中包括: 10× easytaq Buffer 3 μL, 2.5 mmol/L dNTPs 1.5 μL,TaqDNA聚合酶0.25 μL, 正反向引物各1 μL, 模板DNA 3 μL,双蒸水20.25 μL。PCR反应程序为: 94℃预变性5min, 94℃变性30s, 54℃退火45s, 72℃延伸1min,扩增35个循环, 72℃延伸10min。PCR产物经1%的琼脂糖凝胶电泳检测后, 产物送商业公司完成纯化及双向测序。
1.3 序列处理及分析
使用Clustal X1.8软件[29]对DNA序列进行比对,参照测序峰图在SEAVIEW软件中辅以手工校正[30]。采用MEGA5.0软件将其翻译成相应的氨基酸序列,以避免测序等原因造成的人为错误[31]。
1.4 分子系统发育树的构建
采用MEGA5.0软件基于Kimura 2-papamter模型计算物种间遗传距离[31]。采用邻接法(Neighborjoining, NJ)构建分子系统发育树。系统树各分支的置信度采用自引导(Bootstrap)以1000次重复抽样进行检验。
1.5 群落等级聚类分析
本研究根据不同种类在各采样点的出现情况建立 1/0(是/否)矩阵进行分析, 其中横坐标为物种,纵坐标为样点。采用软件 PRIMER 5.0, 以 Bray-Curtis 相似性系数为基础构建不同采样点的相似性矩阵, 采用等级聚类(未加权组平均法, 即 UPGMA)的分类方法构建聚类分析图[32]。
1.6 群落系统发育结构指数计算
采用基于系统发育树枝长距离矩阵的平均成对系统发育距离(Mean Pairwise Distance,MPD)以及标准化用于衡量观测到的系统发育多样性与零模型随机生成的预期值之间差异程度的净亲缘关系指数(Net Relatedness Index,NRI)评估不同空间尺度的鱼类群落系统发育结构[33]。使用R软件中的picante软件包计算群落系统发育结构指数, 具体公式如下:
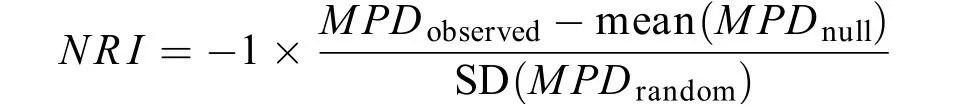
式中,MPDobserved为平均系统发育距离的观察值,mean(MPDnull)是系统发育树上物种随机分配(Taxa shuffle)运行999次进行1000次迭代产生的每个群落999个随机MPD值的平均值, SD(MPDrandom)则是随机MPD值的标准差。零模型选用限制性独立交换算法(Independent swap)。如果量化后的群落系统发育结构指数显著大于零模型, 则表明该群落系统发育结构聚集(物种之间的亲缘关系比预期的近);反之, 则表明该群落系统发育结构发散(物种之间的亲缘关系比预期的远)[34]。
2 结果
2.1 长江中游鱼类群落物种组成
本研究在长江中游5个采样点共采集到鱼类79种, 隶属于6目14科55属, 其中鲤形目55种, 鲇形目11种, 鲱形目2种, 鲈形目9种, 合鳃鱼目1种, 鲟形目1种(表 1)。宜昌共采集到物种25种, 总计3266尾,总重737.582 kg, 其中鲤形目17种, 鲇形目5种, 鲈形目2种, 鲟形目1种。枝江共采集到鱼类36种, 总计2002尾, 总重92.372 kg, 其中鲤形目23种, 鲇形目8种, 鲈形目4种, 鲱形目1种。荆州共采集到鱼类41种, 总计1825尾, 总重117.259 kg, 其中鲤形目28种, 鲇形目6种, 鲈形目7种。汉南共采集到鱼类38种, 总计2193尾, 总重40.385 kg, 其中鲤形目26种,鲇形目4种, 鲈形目6种, 鲱形目2种。湖口共采集到鱼类56种, 总计1065尾, 总重197.052 kg, 其中鲤形目42种, 鲇形目8种, 鲈形目4种, 合鳃鱼目1种, 鲱形目1种。
2.2 空间聚类分析
依据不同鱼类物种在各采样点的出现情况进行聚类, 聚类分析结果显示, 在65%的相似性水平上, 所有样点可以划分为3个Group(图 2), 其中GroupⅠ仅包括宜昌(S1)1个采样点; Group Ⅱ包括枝江(S2)及荆州(S3)2个采样点; Group Ⅲ包括汉南(S4)及湖口(S5)2个采样点。在55%的相似性水平上, 所有样点可以划分为2个Group(图 2), 其中Group A仅包括宜昌(S1)1个采样点; Group B包括枝江(S2)、荆州(S3)、汉南(S4)及湖口(S5)4个采样点。聚类分析的结果与采样点的空间分布相符合。
2.3 不同尺度系统发育群落结构
由于本研究采集的物种为辐鳍鱼类, 所以采用辐鳍鱼类的姐妹群肉鳍鱼类矛尾鱼(Latimeria chalumnae)为外类群(GenBank登录号NC001804),构建长江中游79个物种基于线粒体Cytb基因的分子系统发育树(图 3)。
对不同尺度的长江中游5个采样点鱼类群落构建机制进行探究, 发现各尺度鱼类群落构建机制均不表现为随机作用。以地区采样点尺度(Local scale)对长江中游鱼类群落构建机制进行分析, 发现长江中游4个采样点(即宜昌、枝江、汉南、湖口)的NRI>0, 鱼类群落系统发育结构与随机群落相比更为聚集, 鱼类群落表现为环境因子过滤作用作为建群机制, 荆州的NRI<0, 鱼类群落系统发育结构与随机群落相比更为发散, 群落内物种间竞争作用主导群落构建。
以区域尺度(Regional scale)对长江中游鱼类群落构建机制进行分析, 在65%的相似性水平上, 即划分为3个Group时, 仅GroupⅠ(即宜昌)采样点NRI>0, 鱼类群落系统发育结构表现为聚集, 环境因子过滤作用主导群落构建, 其余2个Group(即枝江+荆州、汉南+湖口)均为NRI<0, 鱼类群落系统发育表现为发散, 物种间的竞争排斥作用主导群落构建;在55%的相似性水平上, 即划分为2个Group时,Group A(即宜昌)NRI>0, 鱼类群落系统发育表现为聚集, 环境过滤作用主导群落构建, Group B(即枝江+荆州+汉南+湖口)NRI<0, 鱼类群落系统发育表现为发散, 物种间的竞争排斥作用主导群落构建(表 2)。

表1 长江中游5个采样点鱼类群落物种组成Tab. 1 Species composition of fish community in 5 sampling sites in the middle reaches of the Yangtze River

续表 1
3 讨论
3.1 长江中游鱼类群落物种组成
长江中游地区五个江段采集的鱼类自上游向下游表现为鱼类物种数逐渐增加(除汉南外)的现象, 其中鲤科鱼类均占半数以上, 汉南采样点(38种)略少于荆州(41种)可能是由于存在采样误差,这与Rahel和Hubert[35]提出的自上游到下游鱼类物种多样性增加相一致。其中相较于其余4个采样点宜昌采集到的物种最少, 渔获物种类仅为25种, 且结构较为单一。由于三峡大坝建成蓄水后, 引起河道形态、底质类型及水文条件的改变, 使得长江干流坝下江段鱼类群落结构发生重大变化, 物种多样性降低[36]。
相较于其他4个采样点, 湖口采集到的鱼类种类最多为56种, 鄱阳湖是长江中下游仅存的两个通江湖泊之一, 与长江物质及能量交换频繁, 加上静水、流水生境的互补作用, 形成独特的复合生态系统, 湖口是长江干流与鄱阳湖通江水道的交叉口,其生境相对更为复杂, 静水、流水生境的互补作用使得湖口生物多样性相对较多, 鱼类资源更为丰富[37]。
依据采集到的鱼类物种有无进行聚类, 聚类分析的结果与采样点的空间分布相符合, 推测是由于长江中游自上而下水环境逐渐发生变化, 地理距离相对较近的采样点水流流速、水温、溶解氧等水环境条件较为相似, 鱼类群落中物种组成也更为接近,使得聚类分析结果符合采样点的空间分布。
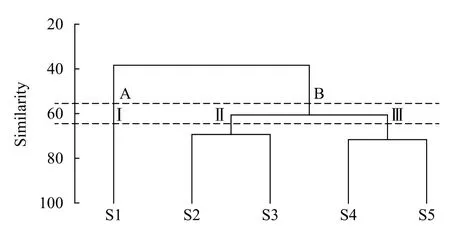
图2 长江中游5个采样点鱼类群落结构空间格局的等级聚类图Fig. 2 Dendrogram of hierarchical clustering based on fish data of 5 sampling sites in the middle reaches of the Yangtze River
3.2 长江中游不同尺度鱼类群落构建机制
地区采样点尺度(Local scale)鱼类群落构建机制长江中游地区5个采样点的鱼类群落系统发育结构分析结果表明, 仅荆州采样点鱼类群落系统发育结构发散, 群落内物种间的竞争排斥作用主导群落构建。荆州江段环境广泛多样, 竞争排斥作用的建群机制使得群落内容纳的多为亲缘关系较远的远缘物种, 许多远缘物种食性相似, 食物来源存在重叠, 如采集到的蒙古鲌(Culter mongolicus)、翘嘴鲌(Culter alburnus)等鲌类与光泽黄颡鱼(Pelteobaggrus nitidus)、鳜(Siniperca chuatsi)等肉食性鱼类多以小型鱼类、虾、水生昆虫为食[22,24,26], 食物来源的重叠使物种间表现了较强的竞争作用。其余4个采样点鱼类群落系统发育结构聚集, 表现为环境过滤作用主导群落构建。群落生境是物种定居于某个区域的首要限制因素, 4个采样点区域空间具有生境异质性, 环境条件如水流流速、水温、溶解氧含量等各不相同,具有特殊性。
区域尺度(Regional scale)鱼类群落构建机制在区域尺度下, 当聚类为3组时, Group Ⅰ(宜昌)鱼类群落系统发育结构聚集, 表现为环境过滤作用主导群落构建;Group Ⅱ(枝江+荆州)及GroupⅢ(汉南+湖口)鱼类群落系统发育结构均转变为发散, 物种间的竞争排斥作用主导群落构建。当尺度进一步扩大即聚类为2组时, Group A(宜昌)鱼类群落系统发育结构始终为聚集, 表现为环境过滤作用主导群落构建。Group B(枝江+荆州+汉南+湖口)鱼类群落系统发育结构则变为发散, 表现为物种间的竞争作用主导群落构建。
然而无论从地区采样点尺度或区域尺度进行鱼类群落系统发育结构分析, 长江中游宜昌江段鱼类群落均表现为系统发育结构聚集, 表明宜昌江段具有强烈的、特殊的环境因子, 致使该位点始终为环境过滤作用主导群落构建。宜昌江段是长江中上游的分界点, 上游河床比降大, 水流湍急, 中游河床比降锐减, 水流较平缓, 宜昌江段其环境条件相对于其他4个采样点更为特殊, 使得该采样点群落构建机制始终为环境过滤作用[18]。在不同的研究尺度下, 荆州采样点由于环境抑制性相对较为均衡,其鱼类群落系统发育结构始终不变表现为发散, 物种间竞争作用主导群落构建。这反映出荆州江段环境的广泛多样, 容纳了较多的远缘物种。其余江段(枝江、汉南、湖口)在地区采样点尺度下, 由于区域空间生境具有异质性, 各采样点江段特殊的环境因子使得仅适应该环境条件的物种得以共存, 群落内多为近缘物种, 各江段群落的构建机制均表现为环境因子过滤作用; 随着研究尺度的扩大即在区域尺度下, 环境多样性增加, 群落内容纳的远缘物种增加, 从而导致鱼类群落构建机制发生转变, 由环境因子过滤作用转变为物种间竞争作用。
已有研究发现群落构建成因会由于尺度的不同而存在差异, 体现出空间尺度在群落构建机制研究中的重要性[13,38,39]。Hubert等[38]分析了印度洋及太平洋3个岛屿、14个采样点珊瑚礁鱼类的群落构建机制。结果发现, 在地区采样点尺度下(Local scale), 14个采样点内部, 群落构成机制为中性随机作用;在区域尺度下(Regional scale), 不同岛屿珊瑚礁鱼类群落构建机制为竞争排斥作用。然而对于陆生植物群落来说, 不同的研究尺度, 群落的构建机制表现不同于鱼类群落。Kembel和Hubbell[39]在研究巴拿马热带雨林植物群落时, 发现在小尺度上, 群落的构成机制为竞争作用, 谱系结构发散, 随着研究尺度的增加, 环境变量逐渐增多, 生境过滤则成为群落构建的主要成因。

图3 基于线粒体Cyt b序列构建的长江中游5个采样点鱼类物种NJ系统发育树Fig. 3 Neighbor-joining (NJ) tree of fish community based on Cyt b gene from 5 sampling sites of the middle reaches of the Yangtze River
本研究对长江中游干流不同尺度鱼类群落结构构建机制进行分析, 发现在地区采样点尺度下,荆州江段鱼类群落构建机制为竞争作用, 其余4个采样点均表现为环境过滤作用主导群落构建; 在区域尺度下, 宜昌江段鱼类群落构建机制始终为环境过滤作用, 荆州江段鱼类群落构建机制始终为竞争作用, 其余3个采样点(枝江、汉南、湖口)扩大尺度后鱼类群落构建机制发生转变, 由环境过滤作用转变为竞争排斥作用主导群落构建。与Rahel等[35]的研究结果类似, 存在部分一致性。这反映出水生动物与陆生植物在不同的研究尺度上群落构建机制存在差异。陆生植物群落环境因素对其物种组成有着非常重要的作用, 小空间尺度下的生境较为均质, 资源相对有限, 物种间的竞争作用主导群落构建[11], 随着研究尺度的增加, 群落内温度、水分、风力、光照和土壤等诸多生态因子增加, 环境因子过滤作用成为群落构建的主导因素[12,40]。但在水体环境中, 随着研究尺度的增加, 水环境的多样性使得群落内容纳了更多的远缘物种, 群落构成的生态机制表现为竞争作用。因此, 不同的生境,不同的生物类群其群落构建机制在不同尺度上表现不同。此外, 本研究显示, 在长江中游干流的宜昌江段与其他江段有明显的差别。因此, 在未来的保护工作中应予以关注, 区别对待。
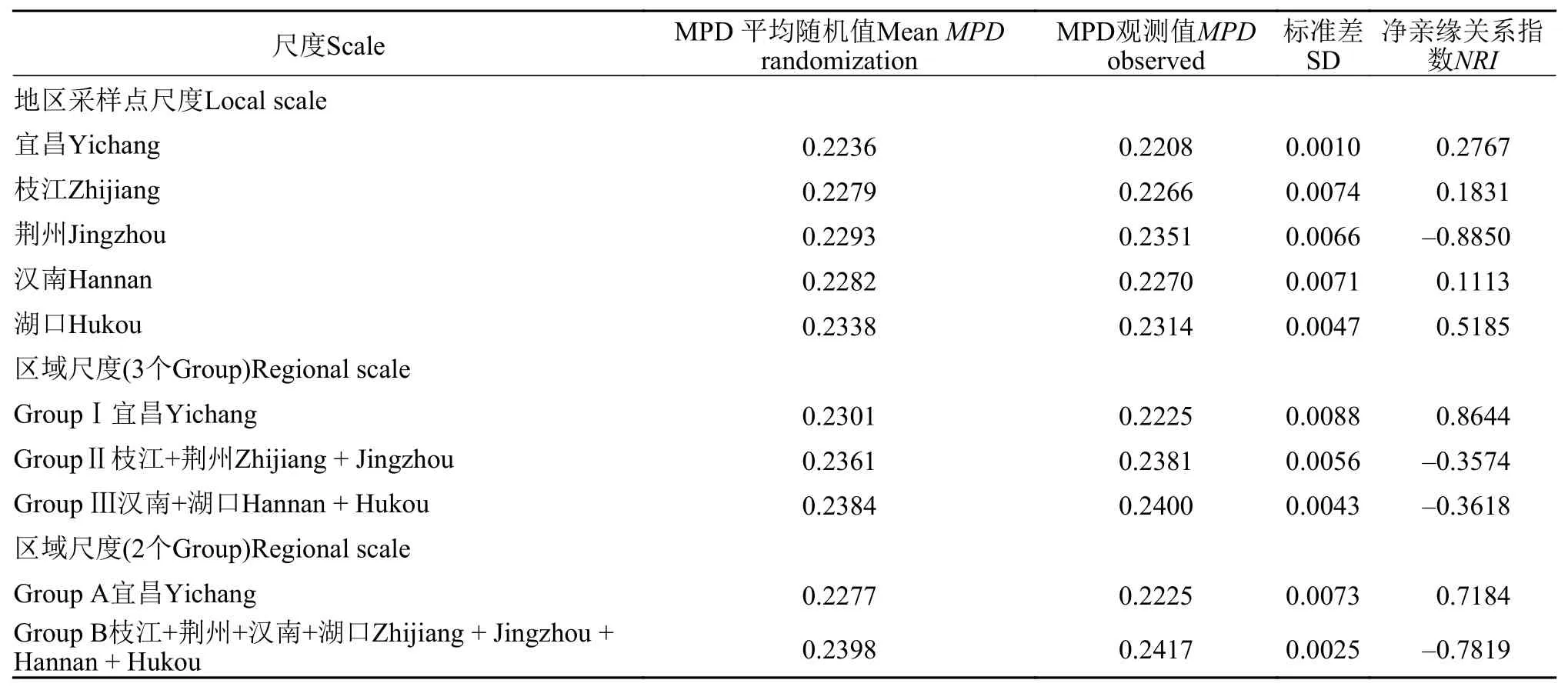
表2 长江中游鱼类群落不同尺度下平均成对分类系统发育距离(MPD)和净亲缘关系指数(NRI)Tab. 2 Mean pairwise phylogenetic distance (MPD) and net relatedness index (NRI) with different spatial scales of fish community in the middle reaches of the Yangtze River
致谢:
感谢中国科学院水生生物研究所但胜国、张智、陈林、倪述、唐瑞和高嘉昕在采样方面给予的帮助。
猜你喜欢
当代水产(2022年8期)2022-09-20
海洋信息技术与应用(2022年1期)2022-06-05
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
中国水利(2020年23期)2020-12-24
儿童时代·幸福宝宝(2020年9期)2020-09-08
河南科学(2020年3期)2020-06-02
科学导报(2020年27期)2020-05-13
