
NaHS对菘蓝吲哚生物碱合成途径中相关基因表达的影响
2020-10-12 07:36贾红磊刘岚钰刘华欣
陕西科技大学学报 2020年5期
贾红磊,刘岚钰,刘华欣
(陕西科技大学 环境科学与工程学院,陕西 西安 710021)
0 引言
菘蓝是常用中药板蓝根与大青叶的主要来源植物,靛蓝、靛玉红是同属于双吲哚生物碱类的活性物质,二者互为同分异构体.药理研究表明其具有抗微生物、提高免疫力 、抗炎、抗病毒和解热等多种作用[1].21世纪以来,对于菘蓝代谢相关通路,与关键性信号分析有所研究,有关次生代谢物研究较少.目前,广泛认定的靛蓝、靛玉红是菘蓝体内起主要药效作用的次生代谢物成分[2],其中主要萜类部分的合成主要依赖于细胞质中的甲羟戊酸(MVA)途径和质体中的2-C-甲基-D-赤藓糖醇-4-磷酸(MEP)途径,其中MVA途径存在于各类高等植物中,莽草酸在莽草酸激酶(Shikimate kinase,SK)、邻氨基苯甲酸合酶(Anthranilate synthase,AS)等酶的一系列作用下生成合成吲哚[3,4].BcSK是编码SK的基因,SK在大肠杆菌和酵母菌中研究颇多[5,6].AS分为邻氨基苯甲酸合酶亚基合酶(Anthranilate Synthaseαsubunit,ASA),邻氨基苯甲酸合酶亚基合酶(Anthranilate Synthase subunit,ASB),其中,ASA可将分支酸催化为邻氨基苯甲酸,ASB将谷氨酰胺催化裂解为游离铵根离子并将铵根离子传递给亚基,这其中BcASA1,BcASA2,BcASB是编码ASA和ASB的基因,进一步发生催化作用经MVA途径生成吲哚[7].脱氧木酮糖-5-磷酸还原酶(1-deoxy-D-xylulose-5-phosphate reductoisomerase,DXR)是MEP途径上的关键酶,BcDXR是编码DXR的基因,DXR首先在拟南芥中被发现,是目前药用植物的研究热点[8].
H2S在动植物体内是广泛存在,植物中H2S的产生主要是由d-半胱氨酸脱巯基酶 (d-cysteine desulfhydrase,DCD) 和LCD催化半胱氨酸分解产生的[9,10],近期也有越来越多的研究表明了H2S在动植物体内可作为第二信使,能够改善植物叶片光合作用,并用来调节植物对非生物胁迫的应答[11-14].其H2S信号分子在植物生理过程中发挥着非常关键的作用,参与着调控植物多种生理过程.Lin等[15]研究结果表明,H2S供体NaHS可以诱导黄瓜幼苗不定根的产生.Fang等[16]研究发现,NaHS处理番茄幼苗可提高其根系中硫化氢合成酶活性和内源H2S的含量.所以运用NaHS处理植物可以进一步分析菘蓝耐药基因相关表达量变化,创新性的从分子学角度定量分析也是具有一定研究意义的.
因此,本文利用实时定量PCR技术对菘蓝叶子和根中BcSK、BcDXR、BcASA1、BcASA2、BcASB的相对表达情况进行分析,探究NaHS与空白组CK处理下对菘蓝幼苗体内相关基因表达的调控作用以及合成前体MEP和MVA途径上相关酶合成基因表达的影响.
1 实验部分
1.1 植物培养
本文所用的菘蓝幼苗均采用水培方式进行培养,一般研究中氮硫元素的配施对于植物中靛蓝和靛玉红的含量大致呈现为高低硫抑制,适宜硫浓度促进的趋势[17],H2S外源供体是NaHS,由贾红磊等[18]研究可知在150μmol/L NaHS处理组下的菘蓝幼苗植株高度有显著性上升,可以缓解Cd胁迫导致的幼苗植株生长抑制作用,所以本实验处理组浓度设定为150μmol/L ,在经过NaHS处理和空白对照ck组处理后后,分别收集处理3 h、6 h、9 h时菘蓝幼苗叶子及根,收样后经液氮速冻后置于-80 ℃冰箱备用.
1.2 幼苗组织总RNA提取
幼苗叶子和根中总RNA的提取采用北京聚合美生物科技公司的植物通用RNA快速提取盒.样品cDNA的合成采用北京聚合美生物科技公司的M5 First Strand cDNA Synthesis kit试剂盒,2μg总RNA建立50μL体系,程序设置为45 ℃,15 min,85 ℃,15 min,反应后即为cDNA样品,将cDNA样品置于-20 ℃冰箱保存.
1.3 实时荧光定量表达分析
选择菘蓝Actin基因作为内参基因,以菘蓝总RNA反转录得到的cDNA模板,并进行RT-qPCR扩增.通过相关文献获得靛蓝和靛玉红合成前体吲哚合成途径上的关键酶SK、DXR、AS的引物序列[19,20].其中所用内参及目的基因引物均由上海生工生物工程股份有限公司合成,引物序列见表1所示.程序设定如下:(95 ℃,60 s)for 1 cycle;(95 ℃,15 s;48 ℃,15 s;72 ℃,30 s) for 40 cycle.每个cDNA样品重复三次实验,使用2-ΔΔCt方法[21]计算相对表达值.
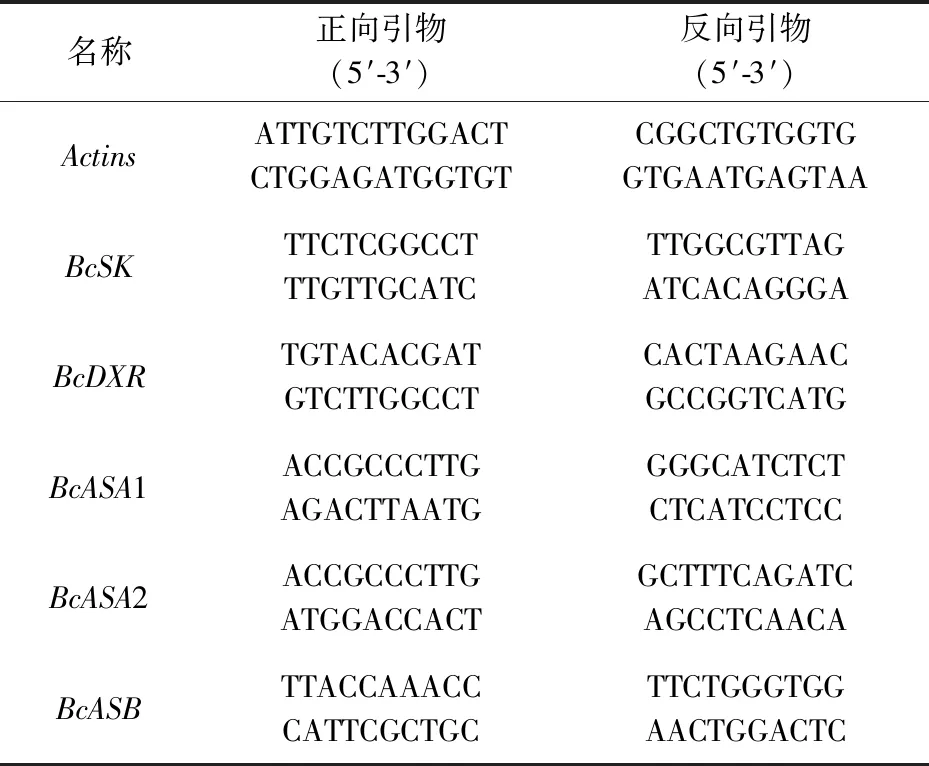
表1 Real-time RT-qPCR引物
2 数据统计与分析
用实时荧光定量表达的分析方法,探究菘蓝幼苗的不同药效基因BcSK、BcDXR、BcASA1、BcASA2、BcASB皆受到外源信号分子NaHS的调控,对吲哚生物碱合成途径中的关键基因进行定量分析,将会更清晰理解药效物质的合成途径并奠定基础,由此来推测吲哚类生物碱次生代谢过程以及调控的变化量,本实验在菘蓝幼苗中的各基因的相对表达情况运用SPSS17.0软件进行显著性分析,所有实验数据均为三次实验均值.
2.1 BcSK在菘蓝幼苗中的表达情况
SK是莽草酸途径上第五步的反应催化酶,也是莽草酸途径中第一个关键酶.BcSK基因在菘蓝幼苗叶片和根部中的表情况如图1所示.与对照CK组相比,在150μmol/L NaHS处理下,BcSK基因的相对表达量在3 h和9 h出现明显上调,在3 h时BcSK相对表达量出现峰值,在6 h时BcSK基因的相对表达量出现明显下调.其中3 h处理下叶片中BcSK基因的相对表达量显著性上调了1094.1%,根中BcSK基因的相对表达量显著性地上调了228.4%.150 μmol/L NaHS处理9 h时,叶片中BcSK基因的相对表达量显著上调了230.9%,根中BcSK基因的相对表达量显著上调了95.3%.由于在6 h时BcSK基因的相对表达量出现明显下调,其中叶片中BcSK基因的相对表达量下调了83.9%,而根部的BcSK基因的相对表达量下调了31.2%.根据以上结果显示,不同时间NaHS处理对于BcSK基因在菘蓝幼体内相对表达量的影响是有差别的.

(a)NaHS处理对菘蓝幼苗中BcSK基因在叶片中表达情况的影响
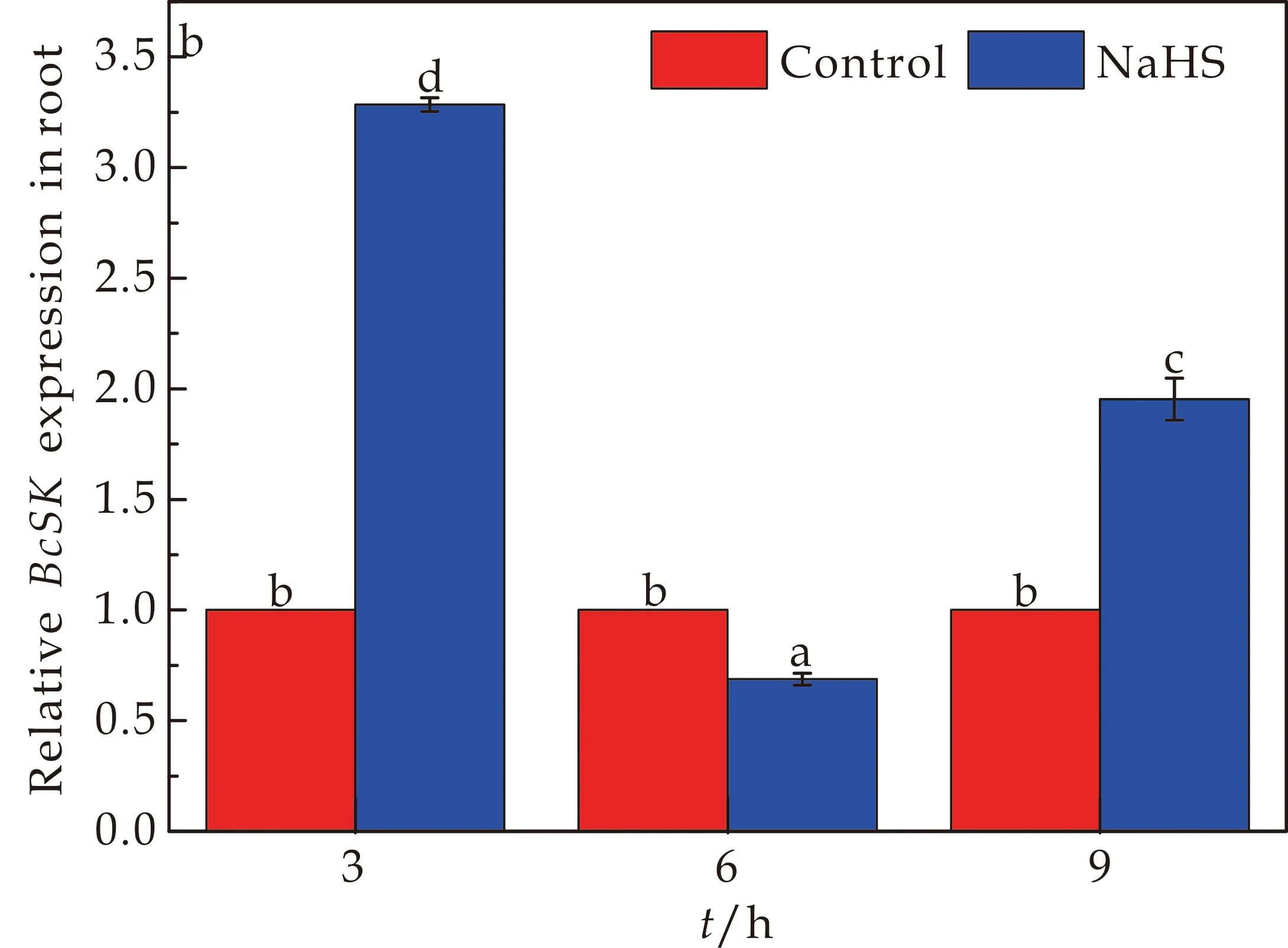
(b)NaHS处理对菘蓝幼苗中BcSK基因在根中表达情况的影响图1 NaHS处理对菘蓝幼苗中BcSK基因表达情况的影响
2.2 BcDXR在菘蓝幼苗中的表达情况
DXR作为合成吲哚类生物碱萜类部分MEP途径上第一个关节酶,对调节吲哚类生物碱的合成有着重要的作用.BcDXR基因在菘蓝幼苗叶片和根中的表达情况如图2所示.与对照CK组相比,150μmol/L NaHS处理下,BcDXR基因的相对表达量在3 h处理情况下叶片中BcDXR基因的相对表达量显著上调了1 789.9%,根中上调了307.0%,在9 h处理条件下,叶片中BcDXR基因的相对表达量上调了51.1%,而根中上调了161.5%.然而,在6 h时 NaHS处理下,叶片中BcDXR基因的相对表达量是上调了50.3%,而在根部地下部分BcDXR基因的相对表达量是下调了15.32%.由以上结果可得短时间内BcDXR基因的相对表达量在根与叶片中主要表现为上调作用,而在6 h处理下叶片与根分别表现出了上调和下调不同作用.

(a)NaHS处理对菘蓝幼苗中BcDXR基因在叶片中表达情况的影响
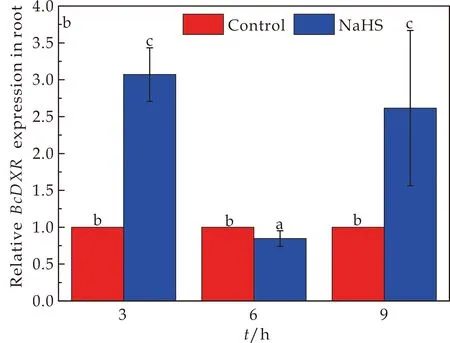
(b)NaHS处理对菘蓝幼苗中BcDXR基因在根中表达情况的影响图2 NaHS处理对菘蓝幼苗中BcDXR基因表达情况的影响
2.3 BcASA1在菘蓝幼苗中的表达情况
ASA是编码邻氨基苯甲酸合酶亚基的基因,本篇选取关键基因BcASA1和BcASA2来进行相对荧光定量分析.其中BcASA1基因在菘蓝幼苗叶片和根部中的表达情况如图3所示.与CK处理组相比,在150μmol/L NaHS对BcASA1基因的相对表达量在不同处理时间表现出不同的调节作用.在3 h时NaHS处理下的叶片中BcASA1基因的相对表达量是显著上调了276.3%,而在根部BcASA1基因的相对表达量下调了24.8%,在6 h叶片中BcASA1基因的相对表达量上调了14%,而在根部BcASA1基因的相对表达量是下调了26.8%,在9 h叶片中BcASA1基因的相对表达量下调了20.9%,在根部BcASA1基因的相对表达量则上调了173.3%.对于BcASA1基因来说,其在NaHS处理下不同时间段的叶片和根部BcASA1基因的相对表达含量趋势是大不相同的,3 h和6 h时在叶片表现为上调,在根部均表现为下调.但在9 h时叶片中的BcASA1基因的相对表达量是显著上调的.
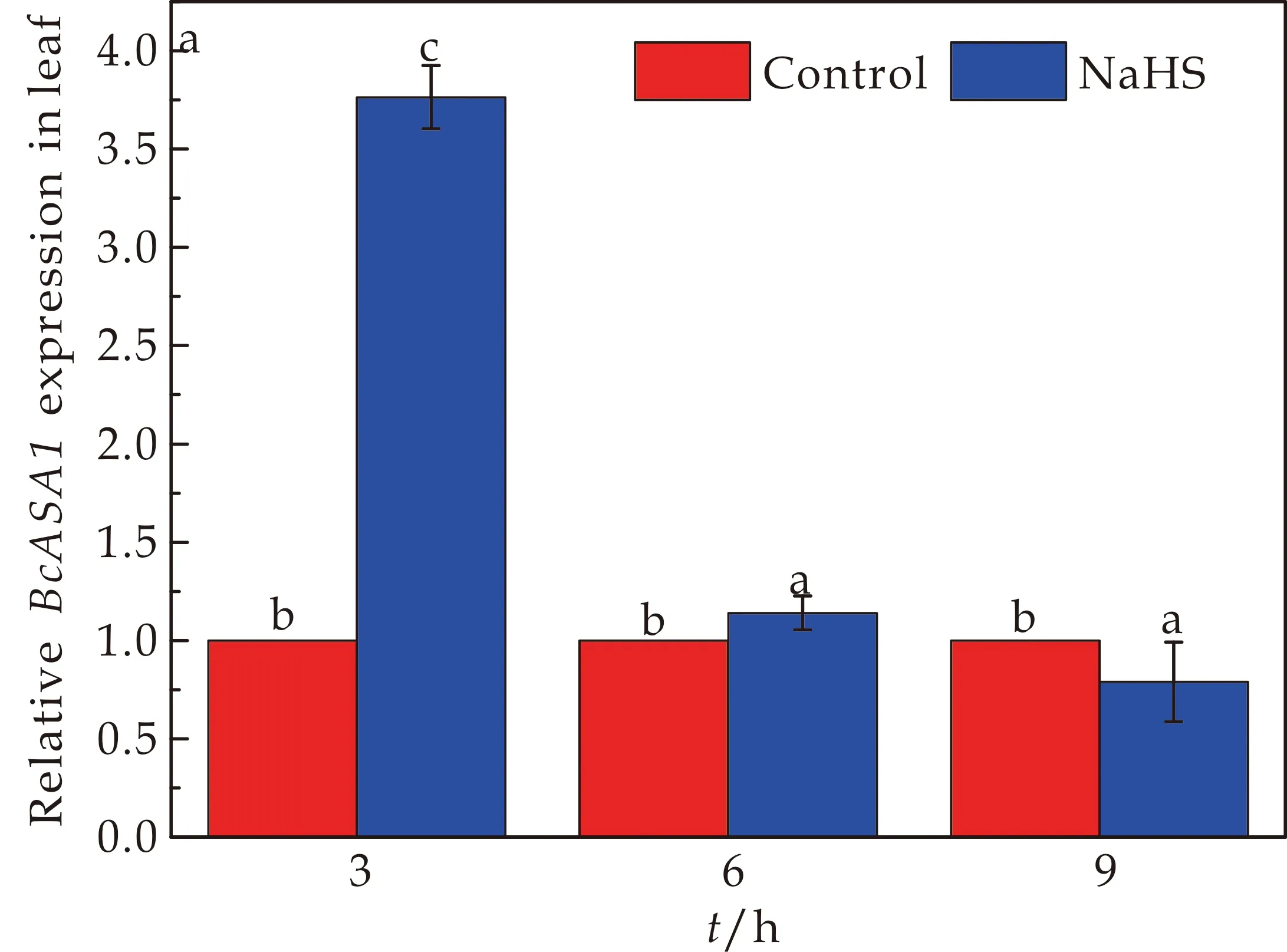
(a)NaHS处理对菘蓝幼苗中BcASA1基因在叶片中表达情况的影响
2.4 BcASA2在菘蓝幼苗中的表达情况
BcASA2基因在菘蓝幼苗叶片和根部中的表达情况如图4所示.与CK处理组相比,在150μmol/L NaHS处理下,叶片与根中BcASA2基因的相对表达量在3 h都出现了明显上调,而在6 h时表现出明显下调,在9 h处理时叶片与根分别表现出不同的趋势.其中,3 h时150μmol/L NaHS处理下叶片中的BcASA2基因的相对表达量上调了252.3%,根部相对表达量上调了141.0%;在6 h时叶片当中的BcASA2基因的相对表达量下调了82.0%,而在根部其相对表达量并无显著变化,在9 h时叶片中的BcASA2基因的相对表达量上调了47.2%,而在根部其相对表达量下调了59.5%.以上结果分析可知,较短时间处理下,叶片和根当中BcASA2基因的相对表达量均表现为明显上调,但长时间的处理下该基因的相对表达情况不一致.

(a)NaHS处理对菘蓝幼苗中BcASA2基因在叶片中表达情况的影响
2.5 BcASB在菘蓝幼苗中的表达情况
ASB是编码邻氨基苯甲酸合酶亚基的基因.BcASB基因在菘蓝幼苗叶片和根部中的表情况如图5所示.与对照组相比,150μmol/L NaHS处理下,叶片中BcASB基因的相对表达量在3 h、6 h、9 h的不同时间均出现下调,而根中BcASB基因的相对表达量在3 h、9 h时出现上调,6 h时出现下调.其中3 h时,NaHS处理下叶片当中的BcASB基因的相对表达量显著下调了70.9%,而在根部当中BcASB基因的相对表达量是上调了16.9%的;6 h时NaHS处理下叶片中BcASB基因无明显变化,根部的BcASB基因的相对表达量显著上调了41.3%;9 h时处理下叶片当中的BcASB基因的相对表达量显著下调了38.2%,在根部中的BcASB基因的相对表达量明显上调了84.2%.上述分析结果可知,在NaHS处理下叶片中的BcASB基因的相对表达量显著下调,长时间的处理下显著上调根中BcASB基因的相对表达量.
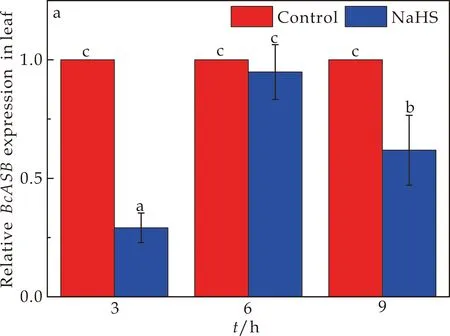
(a)NaHS处理对菘蓝幼苗中BcASB基因在叶片中表达情况的影响
3 结论
本文主要对菘蓝幼苗叶片和根中BcSK、BcDXR、BcASA1、BcASA2、BcASB基因进行相对荧光定量分析.经过不同时间NaHS处理后,BcSK、BcDXR、BcASA1、BcASA2、BcASB基因会表现出不同的调节作用.其中在3 h处理下,这五种基因在菘蓝的叶片和根中大都表现出显著上调作用;在6 h处理下,BcSK基因的相对表达量在叶片和根均表现下调作用,而BcDXR基因的相对表达量在6 h时的叶片与根会表现出上调和下调的不同作用;在9 h时,叶片中的BcASA1基因和根中BcASB基因的相对表达量均是显著上调的,而不同时间处理的叶片中的BcASB基因的相对表达量均表现下调作用.由实验结果表明NaHS以S2-形式的硫素在菘蓝幼苗的相关药效成分合成途径中能够起到非常重要的调控作用.
大多数学者的研究[22-24]多集中在植物次生代谢产物靛蓝和靛玉红的含量测定,国外有研究[25-27]菘蓝根部基因表达的RT-qPCR以及如何获得参与活性成分生物合成的候选基因.而本文在NaHS处理下使植物中的硫元素资源更多转向了次生代谢方向,这也是菘蓝对不利环境的一种适应机制.
因此,利用NaHS来处理可能会诱导菘蓝体内相关基因的合成,从而提高靛蓝和靛玉红的含量,以促进相关代谢产物的合成.已有报道证明BcASA1、BcASA2、BcASB等相关基因表达上调对靛蓝、靛玉红等关键物质的含量具有正相关的影响[20],一方面硫元素作为酶反应中心的重要元素,它的含量可以调节吲哚类生物碱合成相关酶的活性[28],另一方面氮硫配施对菘蓝的靛蓝、靛玉红合成含量具有极显著的影响(P<0.05)[29],会随着硫素浓度的增加呈先上升后下降趋势,也更有利的说明NaHS处理可能激发次生代谢,从而影响靛蓝、靛玉红的含量积累.
猜你喜欢
中国典型病例大全(2022年10期)2022-05-10
今日农业(2021年10期)2021-11-27
中国应急管理科学(2021年9期)2021-03-16
诗潮(2021年1期)2021-03-15
西藏文学(2020年5期)2020-09-21
人物画报(2020年29期)2020-03-14
人物画报(2020年36期)2020-03-13
快乐语文(2019年35期)2020-01-06
中国诗歌(2019年6期)2019-11-15
腹腔镜外科杂志(2016年11期)2016-06-01
