
不同负荷运动训练对大鼠肝脏及骨骼肌线粒体呼吸链功能的影响
2020-10-09 12:55:18黄彩云
上海体育学院学报 2020年10期
李 洁,黄彩云,王 艳
(1.西北师范大学体育学院,甘肃兰州730070;2.甘肃中医药大学体育健康学院,甘肃兰州730099)
随着现代竞技体育的发展,竞技运动能力的竞争日趋激烈。在训练中运动员被要求不断承受超负荷刺激,以使机体产生新的适应,从而达到提高运动能力的目的。但机体对过强刺激不能产生良好的适应,甚至可能导致损伤;过弱的刺激则会大大延长适应的发生过程或无法产生运动训练效果,只有在生理范围内适宜的刺激才能加快适应过程。因此,适宜的运动负荷是关系到运动训练安全和有效的重要因素之一。线粒体是真核细胞内三羧酸循环、呼吸及合成高能磷酸化合物最旺盛的部位和主要场所。自20 世纪60 年代以来,人们一直密切关注运动与线粒体的关系,有关运动与线粒体呼吸链功能方面的研究已有报道,如:大鼠递增负荷力竭性运动导致肝组织线粒体呼吸链复合体酶(C)Ⅰ损害,呼吸链底物堆积[1];大鼠耗竭游泳后,心肌和股四头肌线粒体脂质过氧化水平升高,心磷脂含量下降,细胞色素C 氧化酶活性降低[2]。本课题组前期的研究[3-4]也发现,大强度间歇疲劳运动及中等强度持续疲劳运动对大鼠肝脏及骨骼肌线粒体呼吸链酶活性均有不同程度的影响。经8周大强度运动训练,力竭运动后大鼠肝脏线粒体CⅠ、CⅡ和CⅣ活性均显著提高[5],骨骼肌线粒体CⅠ~Ⅲ活性均显著升高,CⅣ活性显著下降[6]。以上研究说明,力竭运动及不同强度疲劳运动对机体组织线粒体呼吸链功能产生影响,耐力训练对线粒体呼吸链功能亦可产生影响,且存在组织差异性。从以往研究可以看出,运动模式多样、运动强度或负荷不一导致研究结果系统性、可比性较差。因此,本文采用低负荷、中等负荷、高负荷及极高负荷运动训练,进行不同负荷运动训练对大鼠肝脏和骨骼肌线粒体呼吸链功能影响的比较研究,力求为运动训练提供科学方案。
1 材料与方法
1.1 实验对象及分组
选取雄性健康2 月龄Wistar 大鼠60 只,体质量180 g 左右(购于甘肃中医药大学动物实验室,动物生产许可证:甘SCXK 2011-001)。经适应性喂养和跑台适应性运动训练后,保留50 只大鼠进行正式实验。将大鼠随机分为5 组(n=10):对照(C)组、低负荷运动训练(LT)组、中等负荷运动训练(MT)组、高负荷运动训练(HT)组和极高负荷运动训练(ST)组。
1.2 主要药品和仪器
烟酰胺腺嘌呤二核苷酸(nicotinamide adenine dinucleotide,NADH)、细胞色素C(Cyt C)、辅酶Q0、抗酶素A、鱼藤酮、牛血清白蛋白、考马斯亮蓝G250均为Sigma 公司产品,2,6-dichloroindophenol(DCPIP)、sodium salt hydrate、dodecyl-β-D-maltoside(β-裂解剂)均为Fluka 公司产品,其余药品为国产分析纯。DSPT-202 动物跑台为中国杭州钱江科工贸公司制造,Universal 320R 低温高速离心机为德国制造,UVmini-1240岛津紫外可见光分光光度计为日本岛津精密科学仪器有限公司制造。
1.3 训练方案
参照文献[7-9]方法,各运动训练组大鼠进行递增负荷运动训练。LT 组跑台坡度为5%,速度由8 m/min 递增至 15 m/min,MT、HT 和 ST 组跑台坡度分别为5%、10%和20%,速度均由18 m/min 递增至27 m/min,各组运动时间均由20 min 递增至60 min。每天(18:00—20:00)训练1次,每周训练6 d,共训练8周。C 组常规饲养不进行运动训练。具体训练方案如表1所示。训练前、后称体质量。
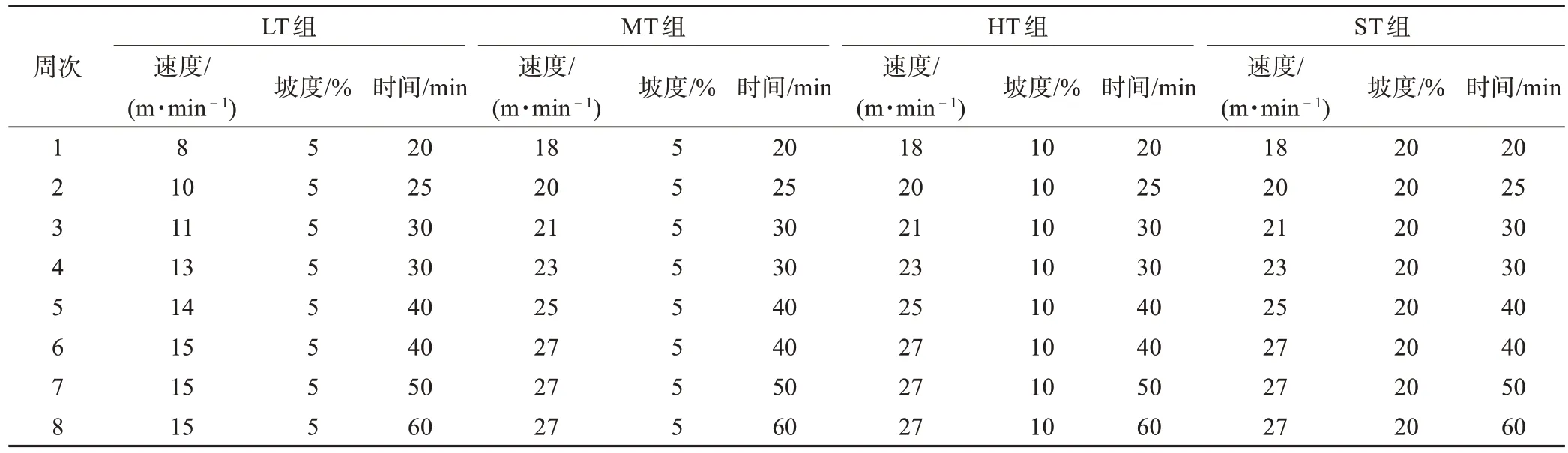
表1 训练方案Table 1 Training program
1.4 取材及线粒体提取
按每100 g体质量1 mL的剂量,腹腔注射0.3%的戊巴比妥钠麻醉大鼠,之后于冰盘上迅速取出肝脏及左、右肢股四头肌,分别置于冷生理盐水中清洗积血,滤纸吸干用锡纸包好,置于液氮中冷冻,-20℃低温保存待用。按文献[10]方法进行肝脏组织匀浆和提取线粒体,按文献[11]方法进行肌肉组织匀浆及提取线粒体。
1.5 指标测定
(1)线粒体蛋白质含量:以牛血清白蛋白为标准,采用考马斯亮蓝法进行测定。
(2)NADH、DCPIP、Cyt C 消光系数:分别制作NADH、DCPIP 和Cyt C 的浓度梯度,在一定波长的光波下,用光径10 mm 的比色杯比色,记录消光值,做出标准曲线,标准曲线的K值为各物质的消光系数。
(3)线粒体CⅠ~Ⅳ活性:参照文献[12]的方法,将10~20 μg 线粒体蛋白加入终体积为2 mL 的缓冲液中,以蒸馏水作空白管,校正光密度到0 点,分别测定340、600、550 nm 处3 min 吸光度(A)值的变化。复合体酶活性(μmol·mg-1·min-1)=[(△A/min×反应体积)/吸光系数]/线粒体蛋白含量,其中△A指1 min 内A值变化。
1.6 统计学处理
所有实验数据以均数±标准差表示,组间数据用SPSS 21.0 软件进行单因素方差分析,P<0.05 表示差异具有统计学意义。
2 结果
2.1 体质量比较
训练前,各组大鼠体质量之间无显著性差异,8周后,与C组相比,LT组大鼠体质量下降5.53%、MT组下降2.63%、HT 组下降11.95%,但与训练前相比均无显著性差异,ST组显著下降38%(P<0.05)。不同负荷训练组间,ST组与LT组、MT组、HT组相比均显著下降(P<0.05),分别下降23.53%、25.63%和18.88%,LT 组、MT组和HT组间无显著性差异。大鼠体质量由低到高顺序为:ST组<HT组<LT组<MT组<C组(表2)。

表2 大鼠体质量变化Table 2 Body weight change in rats
2.2 NADH、DCPIP、Cyt C消光系数
通过在反应体系中建立NDAH、DCPIP 和Cyt C浓度梯度,分别在 340、600 和 550 nm 处读数,做出标准曲线,得NADH 消光系数为7.371,DCPIP 消光系数为17.17,Cyt C消光系数为9.01,用于酶活性的计算。
2.3 线粒体蛋白质含量
以牛血清白蛋白为标准,采用考马斯亮蓝法测定各组大鼠肝脏和骨骼肌组织中提取的线粒体蛋白质含量,用于酶活性的计算。其中,C、LT、MT、HT、ST组肝脏线粒体蛋白质含量分别为1.855、1.920、1.810、2.040、1.883 mg/mL,骨骼肌组织线粒体蛋白质含量分别为0.883、1.259、1.505、0.914、0.584 mg/mL。
2.4 肝脏线粒体CⅠ~Ⅳ活性比较
肝脏线粒体 CⅠ活性,与 C 组相比,LT 组和HT 组显著升高(P<0.01),分别升高88.45%和72.78%,ST组显著下降31.08%(P<0.05),MT 组下降10.23%,但无显著性差异。与LT 组相比,MT 组和ST 组显著下降(P<0.01),分别下降52.36%和63.42%,HT 组下降8.32%,但无显著性差异。与MT组相比,HT组显著升高92.46%(P<0.01),ST 组显著下降23.22%(P<0.05)。与HT组相比,ST组显著下降60.11%(P<0.01)。
肝脏线粒体CⅡ活性,与C 组相比,LT 组和ST 组显著下降(P<0.01),分别下降27.52%和35.93%,MT组和HT组分别上升0.82%和5.95%,但均无显著性差异。与LT 组相比,MT 组和HT 组显著升高(P<0.05),分别升高39.09%和29.75%,ST 组下降11.61%,但无显著性差异。与MT 组相比,ST 组显著下降36.46%(P<0.01),HT 组下降6.72%,但无显著性差异。与HT组相比,ST组显著下降31.88%(P<0.01)。
肝脏线粒体CⅢ活性,与C 组相比,MT 组显著升高21.77%(P<0.05),HT组显著下降32.32%(P<0.05),ST 组升高9.03%,LT 组下降3.54%,但均无显著性差异。与LT 组相比,MT 组显著升高26.24%(P<0.05),ST 组升高13.03%,HT 组下降18.53%,但均无显著性差异。与MT 组相比,HT 组显著降低35.47%(P<0.01),ST 组降低10.47%,但无显著性差异。与HT 组相比,ST组显著升高38.74%(P<0.01)。
肝脏线粒体CⅣ活性,与C组相比,MT组和HT组显著下降(P<0.05),分别下降21.44%和23.89%,LT组和ST 组分别升高14.55%和0.77%,但均无显著性差异。与LT 组相比,MT 组和HT 组显著下降(P<0.01),分别下降31.42%和33.56%,ST组下降12.03%,但无显著性差异。与MT 组相比,ST 组显著升高28.26%(P<0.05),HT组下降3.12%,但均无显著性差异。与HT组相比,ST组显著升高32.39%(P<0.01)。
肝脏线粒体CⅠ~Ⅳ活性从高到低顺序,CⅠ为:LT 组>HT 组>C 组>MT 组>ST 组。CⅡ为:C 组>HT 组>MT 组>LT 组>ST 组。CⅢ为:ST 组>LT 组>MT 组>C 组>HT 组。CⅣ为:LT 组>ST 组>C 组>HT组>MT组(表3)。
2.5 骨骼肌线粒体CⅠ~Ⅳ活性比较
骨骼肌线粒体CⅠ活性,与C 组相比,ST 组显著升高71.65%(P<0.01),LT 组和MT 组均显著下降,分别下降24.18%(P<0.05)和44.51%(P<0.01),HT 组升高1.84%,但无显著性差异。与LT 组相比,HT 组和ST 组均显著升高,分别升高 34.32%(P<0.05)和126.4%(P<0.01),MT 组显著下降26.85%(P<0.05)。与MT 组相比,HT 组和ST 组显著升高(P<0.01),分别升高83.62%和209.48%。与HT 组相比,ST 组显著升高68.54%(P<0.01)。
骨骼肌线粒体CⅡ活性,与C 组相比,ST 组显著升高 51.17%(P<0.01),MT 组显著下降 26.25%(P<0.05),LT 组升高6.02%,HT 组下降17.22%,但均无显著性差异。与LT 组相比,ST 组显著升高42.58%(P<0.01),MT 组和 HT 组显著下降(P<0.05),分别下降30.44%和21.92%。与MT 组相比,ST 组显著升高104.99%(P<0.01),HT 组升高12.24%,但无显著性差异。与HT组相比,ST组显著升高82.63%(P<0.01)。

表3 大鼠肝脏线粒体呼吸链复合体酶活性比较Table 3 Comparison of the activity of respiratory chain complexes in rat liver mitochondria
骨骼肌线粒体CⅢ活性,与C组相比,ST组显著升高37.43%(P<0.01),LT 组显著下降34.12%(P<0.01),MT组和HT组分别下降12.95%和6.33%,但均无显著性差异。与LT组相比,MT组、HT组和ST组均显著升高,分别升高 32.14%(P<0.05),42.18%(P<0.01)和108.61%(P<0.01)。与 MT 组相比,ST 组显著升高57.87%(P<0.01),HT 组升高7.6%,但无显著性差异。与HT组相比,ST组显著升高46.72%(P<0.01)。
骨骼肌线粒体CⅣ活性,与C组相比,LT组和MT组显著下降(P<0.01),分别下降41.57%和38.58%,HT 组升高14.85%,但无显著性差异,ST 组显著升高22.39%(P<0.05)。与 LT 组相比,HT 组和 ST 组显著升高45.72%(P<0.05)和 109.45%(P<0.01),MT 组升高5.11%,但无显著性差异。与MT 组相比,HT 组和ST 组显著升高38.64%(P<0.05)和99.27%(P<0.01)。与HT组相比,ST组显著升高43.73%(P<0.01)。
骨骼肌线粒体CⅠ~Ⅳ活性从高到低顺序,CⅠ为:ST 组>HT 组>C 组>LT 组>MT 组。CⅡ为:ST组>LT 组>C 组>HT 组>MT 组。CⅢ为:ST 组>C组>HT 组>MT 组>LT 组。CⅣ为:ST 组>C 组>HT组>MT组>LT组(表4)。
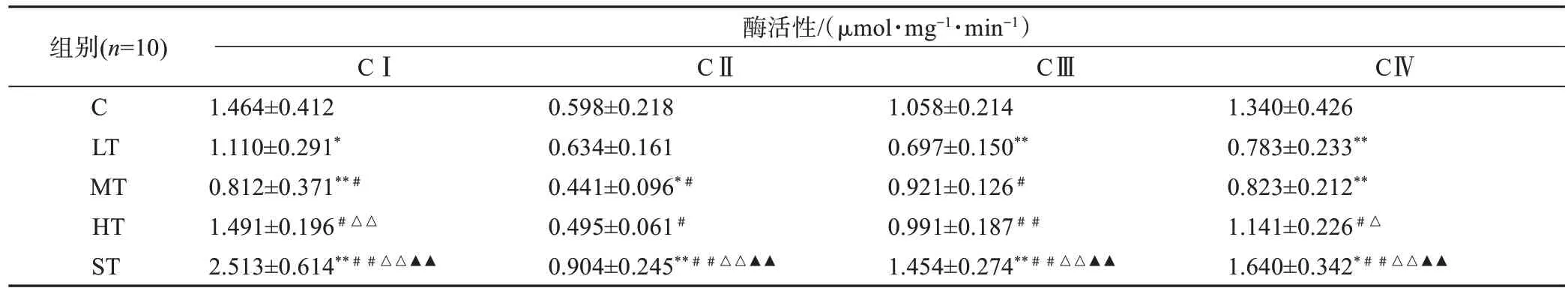
表4 大鼠骨骼肌线粒体呼吸链复合体酶活性比较Table 4 Comparison of the activity of respiratory chain complexes in rat skeletal muscle mitochondria
3 讨论
3.1 不同负荷运动训练对大鼠体质量的影响
体质量可反映机体的营养和肌肉发育程度,是评价机体生长发育及健康状况的重要指标。在运动训练中,通过体质量还可了解运动负荷的大小、训练对机体的影响程度以及机体对训练的适应状况等。已有研究[13]表明,大鼠进行大强度耐力训练7周,力竭运动后即刻,训练组体质量显著低于安静组。在过度训练中,从第5 周开始至训练结束,大鼠体质量呈负增长,而安静对照组在相应时期内体质量呈正增长[14]。在本文实验条件下也出现类似结果,在极高负荷运动训练中大鼠体质量显著下降,而低、中等和高负荷运动训练对大鼠体质量影响不显著。以上研究说明,只有运动负荷达到一定值时才会明显影响大鼠的体质量,这可能是大负荷运动训练导致机体代谢已有平衡被打破,重新建立新的平衡所致。
3.2 不同负荷运动训练对大鼠肝脏及骨骼肌线粒体呼吸链复合体酶活性的影响
已有研究[15]表明,某些呼吸链复合体酶活性的变化有利于运动中细胞稳态的维持。长期规律的运动可改善骨骼肌细胞线粒体呼吸链复合体酶的活性,不同运动方式和持续时间会产生不同的变化效果[16]。
(1)低负荷运动训练8 周可提高大鼠肝脏线粒体NADH 电子传递链起始酶(CⅠ)活性,但由于FADH2电子传递链起始酶(CⅡ)活性下降,从而抵消了NADH 电子传递链起始酶活性的升高效应,致使其后的CⅢ和CⅣ活性无显著性变化。呼吸链不同部位的复合体酶活性变化可能与运动负荷有关。大鼠骨骼肌线粒体CⅠ和CⅡ无显著性变化,CⅢ和CⅣ活性显著下降,致使线粒体呼吸链功能未能得到改善,反而有所下降。究其因,可能是低负荷运动主要动员股四头肌中的慢肌纤维参与募集,而快肌纤维没有或募集较少。低负荷运动可能打破了慢肌纤维原有的能量代谢平衡,经8周运动训练后仍无法良好适应。提示,通过低负荷运动训练改善慢肌纤维的线粒体呼吸链功能,可能需要更长的训练周期。本文结果还说明,肝脏组织线粒体呼吸链起始酶对低负荷运动训练的应激反应较骨骼肌组织更敏感。
(2)中等负荷运动训练8 周后,大鼠肝脏线粒体CⅢ活性显著升高,CⅠ和CⅡ活性无显著性变化,CⅣ活性显著降低。已有研究认为,线粒体呼吸链的限速步骤是在细胞色素b-c1(CⅢ)。提示,中等负荷运动训练可提高肝脏线粒体呼吸链的限速步骤,有望提高呼吸链功能。然而,呼吸链末端氧化酶(CⅣ)活性出现显著降低,从而使呼吸链功能未能得到改善。对骨骼肌的相关研究[17]已有报道,有氧运动2 周可使大鼠骨骼肌CⅠ的活性增强,4周后CⅠ的活性仍在增强,其作用机制可能与新生过氧化氢分子的减少有关;6 周后,CⅠ的活性下降。耐力运动在短期(4 周)内可在一定程度上增加大鼠股直肌线粒体CI的活性,而长期(6~8周)运动只能使其活性维持在较高水平,并不能有所突破;在短期(4 周)内还可提高CⅣ活性,6周时显著提高,8周后有下降趋势;耐力训练无法使训练效果得到长期的突破,对于不断提高运动成绩不够理想,可能需要按周期阶梯式增加耐力训练强度,才能使线粒体功能得到提升[18]。本文中8 周中等负荷运动后,大鼠骨骼肌线粒体CⅠ和CⅣ活性显著下降,CⅡ和CⅢ活性无显著性变化。这可能与在中等负荷运动中主要动员慢肌纤维参与收缩有关。对于提高慢肌纤维线粒体呼吸链功能而言,中等负荷运动刺激可能还较低,使骨骼肌无法产生良好的适应。本文结果还显示,中等负荷运动训练后,肝脏线粒体CⅢ活性显著升高,骨骼肌线粒体CⅢ活性无显著性变化,说明肝脏线粒体呼吸链限速步骤对中等负荷运动的应激反应较骨骼肌敏感。另外,骨骼肌线粒体呼吸链氧化酶(CⅣ)变化趋势与肝脏一致,这可能与中等负荷运动训练后机体有氧代谢增强,导致电子漏增多,过氧化损伤增加有关。
(3)高负荷运动训练8周后,肝脏线粒体CⅠ活性显著升高,CⅡ活性无显著性变化,CⅢ和CⅣ活性显著降低。提示:仅提高运动训练负荷无法达到改善肝脏线粒体呼吸链功能的目的;呼吸链起始酶活性升高,并不意味其后的酶活性也一定会升高,其机制还有待进一步研究。高负荷运动训练8 周后,骨骼肌线粒体CⅠ~Ⅳ活性均无显著性变化。究其因,可能与在高负荷运动训练中,股四头肌慢肌纤维募集减少,主要募集快肌纤维参与收缩有关。欲募集更多快肌纤维参与收缩以及进一步提高骨骼肌线粒体呼吸链功能,可能还需加大运动训练负荷。比较高负荷运动训练对肝脏和骨骼肌线粒体呼吸链功能的影响,可以看出,高负荷运动训练对不同器官线粒体呼吸链功能的影响不同,这与不同器官对运动负荷的敏感性不同有关,对骨骼肌而言,可能还与肌纤维的募集类型和比例有关。
(4)极高负荷运动训练8 周后,大鼠肝脏线粒体CⅠ和CⅡ活性显著下降,CⅢ和CⅣ活性无显著性变化。提示,极高负荷的运动训练会导致肝脏线粒体呼吸链起始酶损伤,对改善呼吸链功能无帮助。大鼠骨骼肌线粒体CⅠ~Ⅳ活性均显著提高,说明极高负荷运动训练可有效提高骨骼肌线粒体呼吸链功能,从而达到提高机体有氧工作能力的目的。其机制可能为,极高负荷运动时募集大量快肌纤维参与收缩,长期运动训练使快肌纤维有氧工作能力得到适应性提高。已有研究可作为佐证,定期高强度耐力训练可增强运动员股外侧肌线粒体氧化碳水化合物的最大能力,并诱导线粒体CⅠ水平的特异性适应[15]。
(5)不同负荷运动训练后,肝脏线粒体CⅠ~Ⅳ活性的变化规律为:CⅠ活性在低和高负荷条件下显著升高,极高负荷条件下显著下降,中等负荷条件下无显著性变化。CⅡ活性在低和极高负荷条件下显著下降,中等和高负荷条件下无显著性变化。CⅢ活性在中等负荷条件下显著升高,高负荷条件下显著降低,低和极高负荷条件下无显著性变化。CⅣ活性在中等和高负荷条件下显著下降,低和极高负荷条件下无显著性变化。由此可见,运动训练对肝脏线粒体呼吸链功能的影响较复杂,影响呼吸链各部位酶活性的因素可能也不同。已有研究发现,线粒体可能成为运动过劳性伤害的重要靶部位[19],大强度游泳运动导致小鼠肝脏尤其是线粒体的结构和功能损伤,线粒体呼吸控制比(RCR)显著降低,通过补充姜黄素能显著缓解其损伤[20]。这与本文中高负荷运动使肝脏线粒体CⅢ和CⅣ活性显著降低,极高负荷运动使肝脏线粒体CⅠ和CⅡ活性显著下降的结果类似。这说明,单靠运动训练提高肝脏线粒体呼吸链功能的效果不佳,可能还需复合线粒体营养物质[21],其干预机制及相关信号通路还有待进一步研究。
(6)不同负荷运动训练后,骨骼肌线粒体CⅠ~Ⅳ活性的变化规律为:CⅠ活性在极高负荷条件下显著升高,高负荷条件下无显著性变化,低、中等负荷条件下显著下降;CⅡ活性在极高负荷条件下显著升高,高、中等负荷条件下显著下降,低负荷条件下无显著性变化;CⅢ活性在极高负荷条件下显著升高,高、中等负荷条件下无显著性变化,低负荷条件下显著下降;CⅣ活性在极高负荷条件下显著升高,低、中等负荷条件下显著下降,高负荷条件下无显著性变化。就本文研究方案而言,通过运动训练改善骨骼肌线粒体呼吸链功能是可行的,且极高负荷运动训练效果最佳。已有研究[22]表明,高强度间歇训练能诱导骨骼肌代谢和性能适应,尽管总体运动量较低,但与传统的耐力训练相似;8 周高强度间歇性训练及间歇+耐力训练使大鼠股直肌线粒体CⅠ和CⅣ活性显著提高,且间歇+耐力训练效果较好[18]。本文研究结果提示,欲发展骨骼肌线粒体呼吸链功能,需极高负荷的运动训练,或采取高强度间歇训练复合中等强度耐力训练。这可能与运动训练使肌红蛋白(myoglobin,Mb)增加,Mb 能增强CⅣ活性以改善线粒体呼吸能力[23],呼吸链复合体质量新分配给超复合体(supercomplexes,SCs)[24],使运动后骨骼肌线粒体 SCs 含量增加有关[25]。其机制还有待进一步研究。
以上研究结果还提示,运动训练对机体运动能力的影响是复杂的,竞技运动能力可能是有极限的。
4 结论
运动训练对线粒体能量代谢的影响具有组织差异性,低、中等、高及极高负荷运动训练对改善肝脏线粒体能量代谢的效果不佳,极高负荷运动训练在提高骨骼肌线粒体能量代谢方面效果较好。
猜你喜欢
海洋通报(2021年1期)2021-07-23 01:55:14
生物学通报(2021年4期)2021-03-16 05:41:26
江苏通信(2020年5期)2020-11-13 15:22:06
广东通信技术(2020年9期)2020-09-22 09:58:54
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
自动化与信息工程(2015年4期)2015-12-23 08:49:49
医学研究杂志(2015年5期)2015-06-10 06:43:26
癌变·畸变·突变(2014年1期)2014-03-01 04:39:36
实用老年医学(2013年7期)2013-03-11 18:39:07
