
海洋桡足类作为生物活饵料的研究进展
2020-10-05 04:58:50程浩楠王淑红程方平
海洋渔业 2020年4期
程浩楠,王淑红,程方平
(1.集美大学水产学院,观赏水族福建省高校工程研究中心,福建厦门 361021;2.中国科学院海洋研究所海洋生态与环境科学重点实验室,山东青岛 266071)
桡足类是海洋中常见的后生动物(Metazoa)[1],隶属于节肢动物门(Arthropoda),甲壳动物亚门(Subphylurm Crustaceans),颚足纲(Maxillopoda),桡足亚纲(Copepoda),是一种体型较小的甲壳动物。目前已经记录的桡足类大约有11 500种[2-3],占浮游生物总量的70%以上[4],其成体大小通常在1~5 mm之间。与传统生物活饵料轮虫和丰年虫不同,桡足类富含二十碳五烯酸(EPA,C20∶5n 3)、二十二碳六烯酸(DHA,C22∶6n 3)、游离氨基酸和一些其他必需微量营养素以及消化酶[5-6],这些成分尤其是不饱和脂肪酸是很多海洋生物特别是海洋鱼类早期幼体正常发育的重要保证。桡足类特殊的游泳方式还有助于诱发幼鱼强烈的摄食反应[7-9]。体型较小的桡足类无节幼虫可作为石斑鱼科(Epinephelinae)、笛鲷科(Lutjanidae)[10-11]以及锈红刺尻鱼(Centropygeferrugata)[12]等小型海水神仙鱼的开口饵料。这些产浮性卵的海水鱼仔鱼,开口时口裂通常小于传统生物活饵料[12-14],有些种类虽然可以摄食一些较小的传统生物活饵料(如:超小型轮虫、俄罗斯小卤虫等),但无法很好地消化吸收[15],而桡足类可作为这些种类的优质活饵,提高其幼体存活率。
因此,本文针对目前桡足类作为生物活饵料的研究进展,从种类选择、藻类饵料供应、培养条件(温度、盐度和光照)等方面进行了综述,以期为后续开发桡足类高密度集约培养技术提供参考。
1 桡足类养殖发展面临的问题
1.1 目标养殖种的筛选
鱼类生产性育苗过程中需要大量且稳定的生物活饵料供应,尽管桡足类具有诸多优点,但其作为活饵料在海水鱼类养殖生产中的使用仍然不太普遍。主要原因在于:一、桡足类高密度培养难度大,人工饲育条件下繁殖力较低[1];二、活体桡足类的保存和运输困难;三、对于饵料生物,滞育卵将大大降低保种、运输的难度,然而仅有近岸生活的少数桡足类种类产生滞育卵[16],滞育卵的产生条件、数量以及保存和孵化条件等都远未及卤虫卵成熟,致使桡足类用于生物活饵料具有很大局限性。因此,目前所利用的桡足类基本源自野生捕捞或虾塘粗放培养,极少有稳定的纯种培养。
然而,野外捕捞不仅难以保证饵料的质量和规格,还有可能引入对鱼类幼体不利的种类。研究发现,有的畸异水蚤(Anomaloceraspp.)会捕食箭虫和幼鱼;一种歪水蚤(Tortanusspp.)可捕食体长在16~21 mm的鲱(Clupeapallassi)幼鱼;有些剑水蚤(Cyclopsspp.)能咬破鱼卵卵膜,进入卵的内部,用口器吞噬胚体,有时在一个卵内可以发现数只剑水蚤,导致鱼卵孵化率极大降低,给养殖生产造成损失[17]。因此,充分了解不同种类桡足类的生物学特性、筛选具有人工培养潜力的桡足类种类非常必要。
1.2 发展集约养殖技术
发展桡足类集约化培养技术,是桡足类作为生物活饵料应用的关键。桡足类集约化培养是指通过人工设备模拟自然界环境,进行连续、高密度的桡足类培养,产出种类、质量和数量都可控的桡足类,保障鱼类育苗生产的稳定性。海洋桡足类集约化培养对于解决开口饵料粒径极小的鱼类人工繁育问题具有重要意义,如胄刺尻鱼(Centropygeloriculus)等小型海水神仙鱼,其人工繁育的瓶颈即为缺乏合适的桡足类无节幼虫作为开口饵料。对于提升其他海洋生物苗种质量,制备富含不饱和脂肪酸的桡足类冰鲜饵料也具有广阔的开发前景。然而,目前桡足类集约养殖技术尚处于起步阶段,几十年来,仅有一些零星报道,尚无大规模成功运用先例。
2 桡足类人工繁育研究进展
2.1 种类选择
中国水域分布的桡足类可分为9个目:哲水蚤目(Calanoida)、剑水蚤目(Cyclopida)、猛水蚤目(Harpacticoida)、宽足水蚤目(Platycopioida)、怪水蚤目(Monstrilloida)、管口虱目(Siphonostomatoida)、鞘口水蚤目(Poecilostomatoida)、摩门虱目(Mormonilloida)和小虱目(Misophrioida)[18]。其中哲水蚤类完全营浮游生活;剑水蚤大部分营浮游生活,少数则营寄生或共栖生活;猛水蚤主要营底栖生活,仅有很少的种类及幼体营浮游生活。后6个目则完全营寄生或阶段性寄生生活[17]。
自然界中,桡足类广泛分布于不同的海域及半咸水的河口,由于生活习性及地理区域不同,桡足类形成了不同的生态类群(ecological group)。我国四大海区(南海、东海、渤海、黄海)每个海区的优势生态类群不尽相同。例如,低温低盐类群的真刺唇角水蚤(Labidoceraeuchaeta)和双刺纺锤水蚤(Acartiabifilosa)等分布在近岸低盐海域;角锚哲水蚤(Rhincalanuscornutus)和筛哲水蚤(Undinuladarwinii)等外海高温高盐类群分布在南海和受暖流影响的东海区以及台湾海峡。此外,还有中华哲水蚤(Calanussinicus)和小拟哲水蚤(Paracalaunsparvus)等一些广温广盐性的种类,季节性地分布于各个海域[17]。
综合分析不同水文环境海域的桡足类优势种类及环境因子,可以筛选一些桡足类进行人工培养,探索并优化其培养条件。同时,还需要考虑桡足类的生态学特征,分析最适合的培养条件,如营浮游还是底栖生活、食性、产卵行为、个体大小以及抗逆性等,以完善大规模人工养殖技术。在生产过程中,还应根据养殖对象对生物饵料的具体需求进行定向选择。例如,强额孔雀水蚤(Parvocalanuscrassirostris)的无节幼虫体长仅为80μm左右,可作为某些口裂较小幼鱼的开口饵料。
目前,用于人工活饵料培养的桡足类主要涉及哲水蚤目、剑水蚤目及猛水蚤目中的种类。它们大多营自由生活,分布广泛,抗逆性较强。具有大规模培养潜力的桡足类在自然界中一般为近岸分布,对于温度和盐度变化具有较高的耐受力,其食性偏向于植食性和杂食性,便于人工饲喂。主要为中小型桡足类,具有较短的生活史,可以达到较高的培养密度。纺锤水蚤属、胸刺水蚤属、拟哲水蚤科、伪镖水蚤属和个别猛水蚤都有大规模人工培养的成功范例(表1)。
2.2 人工繁育桡足类的优化:驯化和选育
自然界的桡足类种群引进室内培养后,生存环境发生变化,必然导致一些个体产生应激反应,出现活力和繁殖力下降等一系列降低产量和质量的不利现象,因此就需要对新引入人工繁育体系的桡足类进行驯化。驯化是在实验诱导下,使某个物种原有遗传性的行为、形态或者生理特性发生改变的过程[19-20]。驯化过程中,一些种类只需要在人工条件下连续培养几个世代即可提升其繁殖力和环境耐受力,不需要选育。然而驯化也有可能导致一些良性性状丢失,特别是在家系遗传留存过少和近亲交配时[21-22]。此外,驯化的缺点也可能会产生世代累积效应[23]。
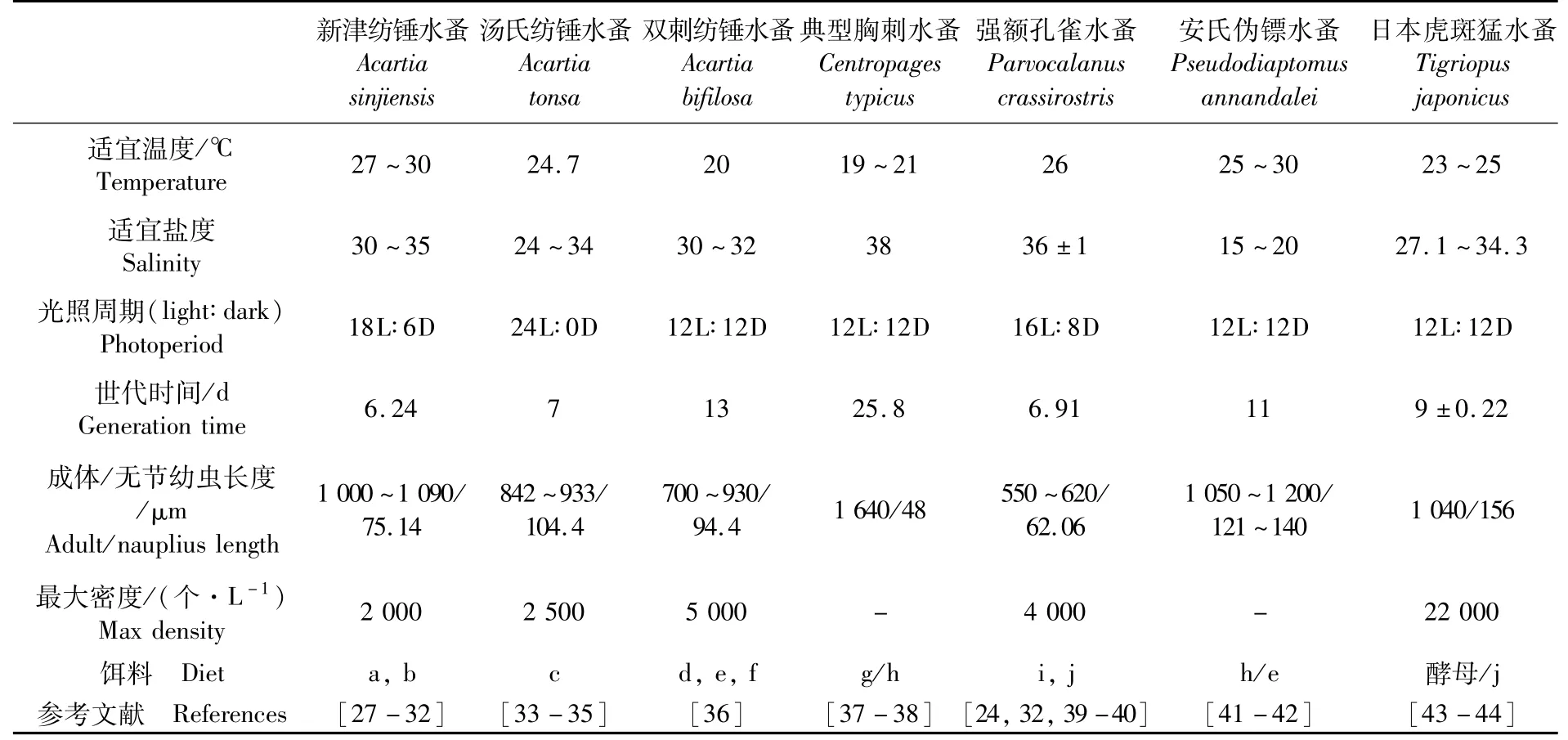
表1 具有大规模培养潜力的桡足类Tab.1 Several kinds of potential copepod species for mass culture
相比鱼虾等大型水产动物,桡足类的世代周期较短。在人工选择压力下,通过一定时间的选育可以大大改善人工繁育桡足类的种质资源,为大规模培养提供更优质的苗种,提高产品品质。目前,仅有少量对桡足类进行人工选育的报道。ALAJMI等[24]对比了在实验室连续培养约80个世代的强额孔雀水蚤与野生种群,发现实验室培养的种群繁殖力和摄食率都远高于野生种,同时死亡率也远低于野生种。ALAJMI等[25]将强额孔雀水蚤同一子代中产卵量在前30%的雌性个体挑出,在连续选育5个世代后,第5代雌性的产卵量对比随机交配的子代显著提高20.5%。但LEE等[26]对矮小拟镖水蚤(Paracyclopinanana)进行选育后,实验种群的繁殖力却未出现明显提升。
虽然不同种类桡足类选育效果不同,但在实际生产过程中非常有必要进行选育以提高种质。在人工繁育过程中,可以通过正向选育强化优良性状,也可以补充野生群体以避免因近亲繁殖而导致的不良性状的累积。对于短世代周期的桡足类,科学的选育实验设计将非常有利于定向选育。
2.3 桡足类培养条件的优化
2.3.1 饵料供应
合适的饵料供应是决定桡足类生长、繁殖和自身生理状态的关键因素。滤食性桡足类通常以单胞藻为饵料,投喂的单胞藻主要包括金藻、硅藻和绿藻等。捕食性桡足类则需要投放一些小型浮游动物,如原生动物和甲壳动物的早期幼虫[17],也可用轮虫、酵母菌、虾片等配合饲料投喂。
在选择藻类作为桡足类饵料时需考虑不同桡足类不同发育时期的适口性,选择大小合适的藻类作为饵料。李捷等[45]分别单独使用4种不同粒径的藻以及混合藻对中华哲水蚤(Calanus sinicus)和双刺纺锤水蚤(Acartiabifilosa)进行投喂实验。结果表明,4个单种饵料中粒径较大的两种能够保证桡足类幼体有着较高的存活率,但由于粒径过大不适于无节幼虫摄食,导致无节幼虫在开口期出现较高的死亡率。上述研究表明,在养殖过程中,应根据投喂对象的发育特点提供适宜的饵料,还可进行多种不同大小的藻类混合喂养。
单胞藻的营养成分对于桡足类大规模培养非常重要[28]。繁殖力是桡足类养殖过程中影响种群补充维持养殖密度的关键参数,饵料种类以及投喂浓度是影响桡足类繁殖力的先决条件。CAMUS等[28]用大溪地等鞭金藻(Isochrysissp.)、周氏扁藻(Tetraselmischuii)、巴夫藻(Pavlovasp.)和牟氏角毛藻(Chaetocerosmuelleri)4种藻,设计了10组不同的饵料配比,投喂拟矮隆水蚤(Bestiolinasimilis),观察了不同藻类配比投喂3 d后的繁殖力。结果表明:金藻、扁藻、巴夫藻(比例为1∶1∶1)喂养的6只雌性个体产卵总数最多,与其他几种配比喂养的雌性桡足类有显著差异,说明不同的单胞藻饵料配比显著影响桡足类的繁殖力。
饵料浓度对于桡足类的繁殖力也有较大影响。KLINE和LAIDLEY[46]分别用密度为100×103、200×103、300×103、400×103、500×103个·mL-1的牟氏角毛藻和等鞭藻(Tisochrysis lutea)(1∶1)的混合饵料投喂种群密度为4个·mL-1的强额孔雀水蚤,结果表明不同饵料浓度对强额孔雀水蚤的存活率无显著影响,但是用密度为500×103个·mL-1藻液投喂的桡足类,其繁殖力几乎高出用密度为100×103个·mL-1藻液投喂的6倍。
值得关注的是,虽然“硅藻—桡足类—鱼类”被认为是海洋生态系统中的经典食物链而备受重视[47],但近20年以来,现场和实验室研究都表明高浓度硅藻对桡足类的产卵和孵化过程具有抑制作用[48-50]。BAN等[51]对37种桡足类 硅藻组合进行实验分析,发现高浓度硅藻(103~105个·mL-1)在36种组合都存在对产卵率(7种组合)、孵化率(11种组合)或二者同时(18种组合)的抑制效应。UYE[52]使用2种微藻投喂太平洋哲水蚤(Calanuspacificus),投喂微小原甲藻(Prorocentrumminium)的对照组整个实验期间卵成功孵化的概率约为96.6%,投喂一种硅藻(Cheatocerosdifficilis)的试验组前5 d卵的孵化率大于84.0%,7 d后孵化率骤降到低于27.3%;IANORA等[53]的实验表明,给克氏纺锤水蚤(Acartia clausi)投 喂 圆 海 链 藻(Thalassiosira rotula)后,其产卵量和寿命都低于投喂微小原甲藻的一半。李捷等[54]的研究也发现与球等鞭金藻(Isochrysis glbana)相比,三角褐指藻(Phaeodactylumtricornutum)对中华哲水蚤的产卵和孵化存在明显的抑制作用。上述研究使得硅藻作为桡足类主要且高效食物来源的地位受到质疑,在人工培养条件下可以使用其他类群的藻类代替硅藻,或者按照一定比例将其他类群藻类与硅藻混合喂养,减轻过多硅藻带来的负面影响[55]。
藻类作为饵料,其质量也影响到桡足类作为生物活饵料的质量,桡足类身体成分中的一些氨基酸和脂肪酸与其食用藻类中这些物质的含量呈现一定的相关性。有研究表明,给中华哲水蚤投喂DHA占总脂肪酸含量30.22%的中肋骨条藻(Skeletonemacostatum)28 d后,DHA含量较自然水体中的提升了约10%;若投喂DHA含量只有0.58%的海洋原甲藻(Prorocentrum micans),其DHA含量只提升约3%[56]。在直接使用10 mmol·L-1的L 赖氨酸、L 亮氨酸、L 异亮氨酸、L 缬氨酸溶液分别对巴拿马异剑水蚤(Apocyclops panamensis)强化3 h后,其体内对应氨基酸含量都有至少2倍以上的提升[57]。因此,通过合理选择微藻饵料或人工添加营养素可以有效提升桡足类体内一些营养成分的含量。
2.3.2 培养温度
温度是影响桡足类分布的重要制约因素,同时也是桡足类生长的重要影响因素。温度对于不同种类桡足类的影响不尽相同,从而决定了它们在自然界中的时空分布模式及生活史[58],可参考桡足类在自然界中的分布和生态习性以初步确定其人工繁育的温度范围。如纽氏伪哲水蚤(Pseudocalanusnewmani)作为冷水种桡足类,水体中的种群数量在温度较高的季节极低,到了冬季才会大量繁殖,根据LEE等[59]的研究,20℃是纽氏伪哲水蚤的温度上限,胚胎发育以及产卵率均受到抑制。桡足幼体2期(copepoditeⅡ)后其体长随着温度的降低而增加,但是其生长速率在15℃以内随着温度的上升线性增加。而MILIONE和ZENG[30]对热带哲水蚤新津纺锤水蚤(A.sinjiensis)的研究表明,在10℃、15℃、20℃、25℃、28℃、30℃的不同温度中培养,随着温度的升高新津纺锤水蚤的种群增长和孵化率都显著提高,在30℃的时候达到最高,这与冷水种纽氏伪哲水蚤的情况大相径庭。HOLSTE和PECK[33]的研究表明,在22.9℃时,以隐藻(Rhodomonassp.)培养的汤氏纺锤水蚤(A.tonsa)平均每只雌性每天可产出50个卵,随着温度升高单个雌性每日平均产卵量随之下降,在34℃时由于雌性全部死亡,产卵量为0。
温度还会对桡足类种群结构产生一定影响,NOGUEIRA等[60]的研究表明,在17℃时纺锤水蚤(A.grani)的种群中有70%是无节幼虫,在28℃时无节幼虫只占种群的5%左右。由此可见,进行人工培养时,应根据所选种类的生物学特点以及培养的目标,确认最佳培养温度。
2.3.3 盐度
盐度作为大规模培养的另外一个重要环境参数,也对桡足类有着一定的影响和制约作用。OHS等[61]研究了盐度为10、15、20、25、30、35的培养环境中对伪镖水蚤(Pseudodiaptomus pelagicus)生理过程及生活史,发现盐度在35和10时无节幼虫存活率分别为59.0%和81.0%,且随着盐度的改变,其种群首次性成熟的时间也随之改变,在盐度为15~30时首次性成熟时间为6.3 d左右,盐度为10时则需约7.5 d,差异较明显[61]。MILIONE和ZENG[30]的研究发现在15~50盐度范围内,新津纺锤水蚤在30盐度下具有最高的种群密度增长率。HOLSTE等[33]研究发现当盐度为0时,汤氏纺锤水蚤卵的孵化率只有11.4%,在盐度为25时孵化率达到峰值84.5%,且在盐度增加至34时孵化率几乎不变。不同地区的天然海水盐度有所差异,在培养不同桡足类时可根据该种类适应盐度及时进行调整,也可尝试进行适当的盐度驯化。
2.3.4 光照
在自然界中,光照通过周期和强度影响桡足类饵料的生长和自身节律,进而影响桡足类种群密度增长。一个完整的光周期为24 h,在全球尺度上,一天24 h中白天和黑夜的占比有较大差异。光照时间对于大多数桡足类赖以为生的微藻生长起着至关重要的作用,而培养系统中微藻的数量又可能成为制约桡足类种群密度增长的因素。然而,人工饲养条件下通常会给予足量或者过量的微藻饵料并且随时补充,这就使光周期间接影响桡足类的作用减弱或消失。
但目前室内实验证明,光谱、光强和周期对桡足类摄食率、产卵率、孵化率、发育过程等方面也有重要影响。CAMUS和ZENG[29]在光照强度为1 250 lux的条件下,研究了光周期(light∶dark)为0 h∶24 h、6 h∶18 h、12 h∶12 h、18 h∶6 h、24 h∶0 h的条件下新津纺锤水蚤的产卵率、孵化率、无节幼虫发育等一系列参数的变化。发现在为期8 d的产卵监测中,具有较长光照时间的两组(24 h∶0 h,18 h∶6 h)具有最高的产卵量和孵化率,且从卵发育至各个时期的时间均明显缩短。不同波长的光照也会影响桡足类摄食,TAO等[62]发现中华哲水蚤在红、橙、黄光下的排粪率显著高于黑暗条件下,且连续9 d的产卵实验中,在强度分别为0.78 mW·m-2、1.58 mW·m-2、2.53 mW·m-2的全光谱照射下的产卵率始终高于黑暗条件下,但在光照强度为2.53 mW·m-2时,表现出较低的孵化率。还有研究发现光周期还能诱 导 桡 足 类 产 滞 育 卵[63-66],CHINNERY和WILLIAMS[64]发现在光周期(light∶dark)为13 h∶11 h、12 h∶12 h时,双刺纺锤水蚤卵中滞育卵的比例分别为90%和10%,两者差异显著。HAIRSTON和KEARNS[67]发现在光照时长从11.75 h提高到13.25 h时,所产卵的滞育百分比从8.7%上升到98.0%。因此,光照条件对桡足类的生理活动和繁殖过程的影响是复杂的,其机理可应用到桡足类人工繁育方面。
2.3.5 培养密度
种群密度是人工培养过程中最重要的养殖参数之一。自然界中桡足类作为一个重要的次级生产者,其种群的增长受到食物、捕食者和生存空间等各种因素的限制,常常处于较低水平。在人工养殖条件下,食物、温盐等条件优化之后,不同种类种群密度能达到的峰值也不尽相同。比起其他传统生物活饵料,轮虫(Brachinousspp.)在常规培养方式下的密度通常只能维持在100~200个·mL-1,但在新研发的培养技术下其培养密度可以达到16万个·mL-1[68]。然而受种种制约,桡足类很难开发高密度人工培养技术,这也是它作为生物活饵料的最大瓶颈。
桡足类的摄食方式分为滤食、摄食碎屑和捕食等3种方式,在高密度培养下桡足类个体之间的竞争与天然环境相比更为激烈。在人工饲养条件下常常观察到它们摄食同种幼体的现象,如:汤氏纺锤水蚤[29]、海岛哲水蚤(Calanus helgolandicus)[69-71]、长 角 宽 水 蚤(Temora Longicornis)[72]和 虎 斑 猛 水 蚤(Tigriopus fulvus)[73]都有摄食无节幼虫的现象,进而影响种群的补充。这可能是桡足类培养中种群密度无法持续升高的重要因素。OHMAN和HIRCHE[74]发现飞马哲水蚤(Calanusfinmarchicus)雌性密度在达到16 000个·m-2时卵具有较高的死亡率,且观察到这种哲水蚤会摄食自己的卵和与卵粒径相似的浮游生物。
桡足类繁殖率在种群密度增长到一定程度时不再升高甚至降低,还可能跟内源性自动调节有关。有报道认为,虽然滤食性的强额孔雀水蚤在培养到一定密度时也会产生种群数量负增长,但目前并没有该种类会摄食幼体的直接证据[46],该研究的结果显示,强额孔雀水蚤在培养密度为2~8个·mL-1时,平均每个个体产生无节幼虫和卵的数目在3 d内持续下降。当密度达到8个·mL-1时,其个体几乎丧失繁殖力。同一研究还显示,每日收取强额孔雀水蚤卵和幼体的组,繁殖率明显高于3天收取一次的组,此现象可能就与强额孔雀水蚤摄食其卵和幼体有关,具体机制尚需进一步研究。
因此,在培育过程中,对于滤食性种类应注意做好成体和幼体的分离工作,尽量减少同类残食现象。同时应关注桡足类培养密度,通过驯化、选育逐步提高其密度,以提高单位水体的饵料产出。
2.3.6 培养环境水质影响
人工培养环境相对自然水体体积较小,进行养殖活动时常常积累一定水平的污染物,如含氮、碳、磷、硫等的代谢废物。这些代谢产物在氧气充足的条件下对水生生物的影响程度不大,但是当其浓度较高达到阈值时,则会产生毒害作用。如氮以分子氨态或亚硝酸盐氨态存在且达到一定浓度时,就会对养殖水生生物产生很强的神经性毒害[75]。目前有关此类污染物对于桡足类存活率影响方面的报道较少,现有报道观点不尽一致。KLINE等[46]发现强额孔雀水蚤培养水体中的N NH3浓度从0提升至0.4 mg·L-1后,对其无节幼虫存活率没有影响。KURIHARA和ISHIMATSU[76]的研究发现,在水中CO2浓度为2 000 mg·L-1时,马纺锤水蚤(Acartiatsuensis)的存活率、幼体发育及体长等均与CO2浓度为380 mg·L-1时无显著差异,但卵的孵化率却比后者降低约10%~18%,差异显著,这可能是CO2改变了养殖水体的酸碱度所致的[77]。
水中溶解氧(DO)的降低也会对桡足类产生影响,LUTZ等[78]的研究发现哈默胸刺水蚤(Centropageshamatus)的卵在DO<0.03 mg·L-1的海水中孵化会受到抑制,但移入海水(0.13 mg·L-1>DO>0.03 mg·L-1)11 d后的孵化率能达到4%~87%,但前5 d卵的孵化率为0%,说明溶解氧较低会导致卵发育迟缓。汤氏纺锤水蚤的卵在0.13 mg·L-1>DO>0.09 mg·L-1的水中孵化率低于10%,在DO<0.09 mg·L-1水中孵化率为0%。
水体中的颗粒有机碳和溶解有机碳也可能对桡足类产生影响。有机碳可能引起整个养殖环境的微生物菌群发生失衡[79-81],从而影响到桡足类的生理生态活动。在饵料投喂过量的水体中,原生动物容易出现暴发,一些原生动物如钟虫(Vorticellaspp.)等可能会粘附在桡足类体表,导致桡足类的死亡[82]。因此在桡足类大规模培养时应注意水质的监测与维护。
2.4 产滞育卵的桡足类用于人工繁育的潜在优势
在温度较高或较低以及食物不充足、光周期、盐度或者其他一些条件改变较大时,部分桡足类会产生滞育卵[16],目前报道的约有51个种可以产出滞育卵,它们分别来自纺锤水蚤科、胸刺水蚤科、角水蚤科、宽水蚤科和歪水蚤科等[83]。MARCUS和MURRAY[84]将实验室产出的小批哈默胸刺水蚤的滞育卵孵化,并成功用于数种鱼类的育苗。但目前滞育卵的研究大多只停留在实验室阶段,将桡足类滞育卵应用于水产养殖中的实例较少。探索自然条件下桡足类卵的滞育条件,以及通过人工手段诱导一些在水产养殖中具有潜力的桡足类产生滞育卵,具有重要的研究意义和实际应用价值。滞育卵易于保存、运输,可大大降低桡足类作为活饵料保种、运输、交易的难度。
3 桡足类集约化培养技术的发展
桡足类集约化培养能将养殖效率最大化,早在20世纪末一些学者[7,85-86]就曾报道过桡足类循环水培养系统,但其构成都较为简单,都是由养殖和过滤等基本单元组成。徐东晖[87]根据培养系统的自动化程度和水体养殖方式,将桡足类集约化培养系统分为:
1)开放式非自动化培养系统,采用敞口容器设计的用于培养猛水蚤Tisbeholothuriae的非自动化装置,培养密度可达115.8个·mL-1[88];
2)封闭式非自动化培养系统,采用封闭的200~450 L水箱连续培养了约70个世代的汤氏纺锤水蚤,成体密度维持在50~100个·L-1[89];
3)封闭循环式非自动化培养系统,采用封闭水箱的自动化培养装置连续培养猛水蚤Amphiascoidesatopus,可日产桡足类干重约5 g[85],国内学者[90]也研发出了一款桡足类循环水养殖设备,在61 d的培养周期中,该装置可以使中华哲水蚤的成体数量由2~3个·L-1稳定至19.7个·L-1,产卵量由14.7个·L-1稳定至85.7个·L-1,而且可以分离卵和成体,防止卵被误食;
4)封闭循环式自动化培养系统,首先是一些简单的自动化设备组成的封闭循环培养系统,连续培养猛水蚤,最高日产无节幼虫约8×105个[7]。后续出现了一款大型桡足类自动连续培养设备,其中引入了大量的自动化控制设计,通过电磁阀和液位开关自动完成幼体收集和水循环过程[91]。此套系统优点在于其自动化程度很高,可减少大量人力投入,但缺点在于其中未加入温控装置,导致其生产过程稳定性不够。CAROTENUTO等[92]则在循环水养殖系统中加入了温度控制系统,使得整套系统的生产稳定性大大增加。
集约化、自动化的养殖设备能够通过可控的系统使养殖水体得到有效净化,提高整个系统连续产出的能力,同时可以定时定量收集需要的不同发育阶段的桡足类幼虫或卵,相较于传统养殖模式能够大大提高养殖效率,增加生产的稳定性,是桡足类集约化培养中不可或缺的一环,具有很好的应用前景。
4 小结
桡足类作为多种海洋生物幼体的天然开口饵料,具有体型小、富含不饱和脂肪酸、易消化等诸多优点,但目前少有集约化稳定供应,限制了其在生产中的应用。桡足类对饵料需求高,多数桡足类必须使用新鲜的微藻喂养;养殖密度难以提升,产量低;种群控制机制不明确,人工培养不能稳定供应等因素是制约桡足类集约化培养技术发展的主要原因。培养种类的选择、培养条件的探索和优化,以及自动化和智能化集约培养设备的研发应是今后主要的研究方向。
猜你喜欢
生物学通报(2021年9期)2021-07-01 03:24:44
辽宁化工(2021年5期)2021-06-03 05:15:50
——以“不同咖啡因饮料对水蚤心率的影响”为例
中学生物学(2021年2期)2021-04-30 12:04:06
现代电生理学杂志(2019年3期)2019-02-21 01:28:35
河北渔业(2019年1期)2019-02-13 01:35:34
小天使·二年级语数英综合(2018年9期)2018-10-16 11:21:22
第二课堂(初中版)(2018年3期)2018-06-12 06:13:52
河南科技(2015年21期)2015-03-26 21:21:21
湖南农业(2015年7期)2015-03-20 01:48:37
海洋与湖沼(2014年4期)2014-03-09 06:57:32
