
云南西双版纳常见寄生植物生态化学计量特征及其与寄主的关系
2020-09-28 03:04门世争刘文耀莫雨轩张婷婷范晓阳
亚热带植物科学 2020年4期
门世争,刘文耀,莫雨轩,张婷婷,范晓阳
(1.中国科学院西双版纳热带植物园热带森林生态学重点实验室,云南 勐腊 666303;2.中国科学院大学, 北京 100049)
生态化学计量学是一门研究组成有机体所需的各种元素之间多重平衡的学科,这些元素能影响营养循环、食物网动态、生态系统生产力,揭示其元素组成对生态功能的影响是该学科的核心内容[1—3]。生态化学计量学也是综合研究碳(C)、氮(N)、磷(P)等化学元素在生物地球化学循环中的耦合关系的新方法[4]。C、N、P元素作为参与构建生物体氨基酸、蛋白质、核酸等物质的基本组成元素,其化学计量关系不仅可以分析植物元素养分来源,还为理解生物地理化学循环和生物多样性变化提供科学依据[1—2,5]。内稳性理论是生态化学计量学的核心理论之一,体现了生物在变化的生境中具有保持自身化学组分及其计量比相对稳定而不随环境剧烈变化的能力[6—7]。不同的植物和组织具有不同程度的化学计量内稳性[8]。
寄生植物是一个特殊的植物类群[9—10],根据其从寄主植物摄取养分的差异[11],可分为全寄生植物(Holoparasitic plant)和半寄生植物(Hemiparasitic plant)两大类。前者不含叶绿素,无叶片或者叶片退化,不能进行光合作用,其生长所需的水分、无机养分和有机养分等都从寄主植物获得;后者有正常的茎、叶分化,但光合水平低下,净同化获得产物少,不足以供自身生长,加之缺乏根系,需靠吸盘、吸器或吸根从寄主中获取水和无机盐[3]。寄生植物不同程度地从寄主植物摄取养分,导致寄主的生长、繁殖、生理代谢等方面受到不利的影响[4,12]。另一方面,虽然寄生植物的生物量较小,但显著影响植物群落结构、物种丰富度,甚至生物地球化学循环[10—12]。研究表明,寄生植物减弱了寄主植物的繁殖能力,降低了寄主的光合速率和竞争能力[13—14],也降低寄主植物的生物量等[15],严重的槲寄生甚至可能导致寄主死亡[16]。此外,寄生植物的凋落与分解对促进生态系统养分循环有一定的作用[17]。以往关于寄生植物的研究多聚焦在寄生植物自身的生物学特性及与寄主间物质交流方面[18—20],也有研究发现寄生植物叶片 C含量与寄主枝条呈负相关关系[21],但寄生植物从寄主获取养分及其转移机制仍然不明确[20,22]。
云南西双版纳是中国生物多样性较丰富的地区,也是全球44个生物多样性热点地区之一[23]。过去几十年来,随着橡胶林在西双版纳地区的广泛种植,大面积热带森林被橡胶园替代,导致生境破碎化,生物多样性受损严重[24]。在西双版纳地区,不同类型寄生植物常见于果园、茶园等人工林及公路边。一些学者调查了该地区桑寄生植物的物种组成[25]、多样性以及分布特征[26],但有关不同类型寄生植物的生态化学计量特征还未见报道。本文拟通过研究云南西双版纳地区常见半寄生和全寄生植物的生态化学计量特征及其与寄主之间的关系,分析不同寄生植物的化学计量内稳性指数及其变化,旨在进一步理解不同寄生植物的养分利用策略与生态适应性,为热带地区生物多样性保护与管理提供科学依据。
1 材料与方法
1.1 研究区概况
云南西双版纳位于中国西南部,地理位置21°09′~22°33′E,99°58′~101°50′N,海拔 550~2429.5 m,总面积19 690 km2。年均温21.7 ℃,年均降水量1560 mm,时空分布不匀,一年被分为旱季(11~4月)和雨季(5~10月),旱季的11月至翌年2月由于低温多雾被称为雾凉季,3~4月少雨高温则为干热季[27]。土壤C、N和P含量分别为62 mg·g-1、2.41 mg·g-1和 0.56 mg·g-1[28]。
云南西双版纳地区寄生植物资源丰富,类型多样。在西双版纳热带植物园内桑寄生植物就有3科8属17种2变种1亚种[25],常见半寄生植物是澜沧江寄生(Scurrula chingiivar.yunnanensis)、五蕊寄生(Dendrophthoe pentandra)、卵叶梨果寄生(Scurrula chingii)、云南槲寄生(Viscum yunnanense)、檀香(Santalum album)等;全寄生植物主要有金灯藤(或日本菟丝子,Cuscuta japonica)、野菰(Aeginetia indica)、大花菟丝子(Cuscuta reflexavar.reflexa)、无根藤(Cassytha filiformis)、扁枝槲寄生(Viscum articulatum)等。
1.2 样地设置与采样方法
西双版纳热带植物园是我国面积最大、收集物种最丰富、植物专类园区最多的植物园,植物园占地面积约1125 hm2,其中园区内保存有一片面积约250 hm2的原始热带雨林。园内寄生植物资源丰富,其中占优势的半寄生植物为澜沧江寄生、五蕊寄生,而全寄生植物以金灯藤分布最为广泛。因此,本研究选取这三种寄生植物作为材料。
采用样带调查法,在园区寄生植物分布较为集中的区域设置3条样线作为重复,每个目标物种在样线上设置5个采样点,于2019年3~4月(旱季)与7~8月(雨季)分别进行调查采样。采集半寄生植物时,选取寄主胸径高度以上的寄生植物收集其成熟叶片和寄生枝条,同时收集寄主的叶片与枝条。采集全寄生植物样品时,由于金灯藤众多柔软的草质茎交织成网状缠绕于群落中寄主植物的茎、叶部位,因此采集时选取全寄生植物分布密集的区域,并且采集该区域内被感染全寄生植物的寄主植物茎、叶样本。土壤样品采集自寄主植物垂直投影下的四个方位,混合成为一份完整的样品。在采样中选择寄主上相近生长时期和相似生境的寄生植物,旱季和雨季的寄主为相同物种。每种寄主设5个重复。
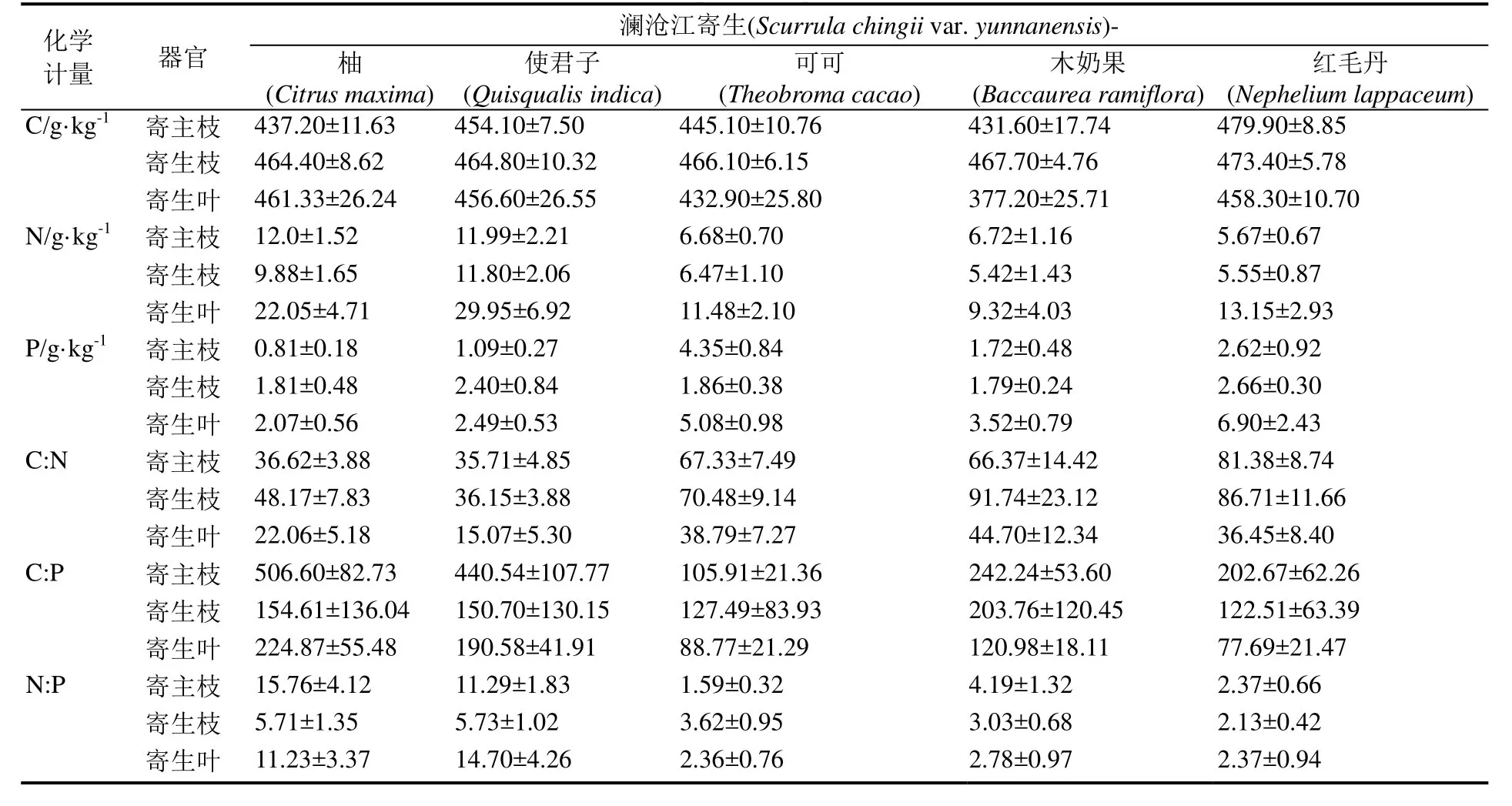
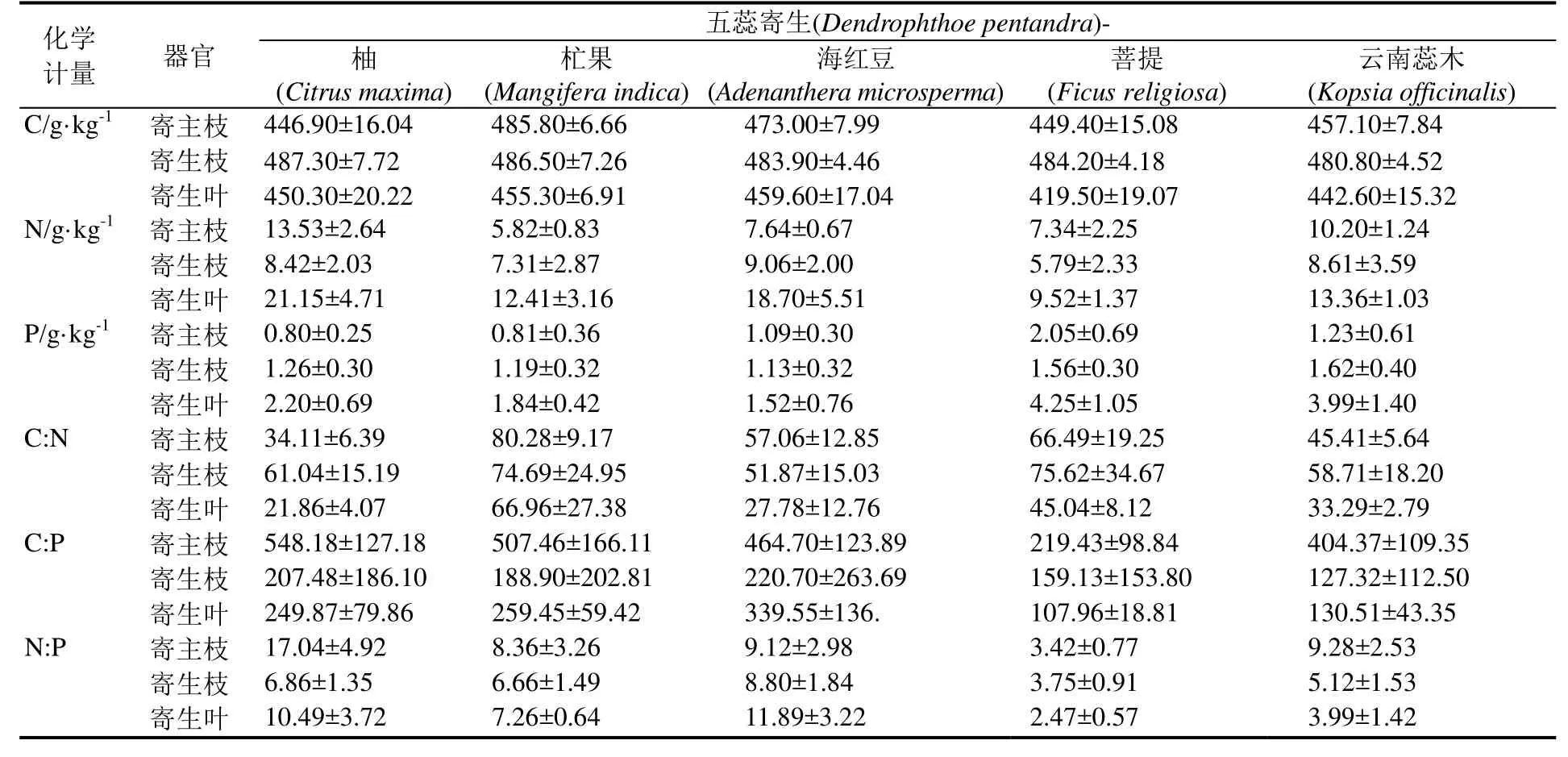
在西双版纳植物园区内,所选3种寄生植物对寄主有较强的选择性,且各自的寄主有所不同。本研究对每一种寄生植物选择寄生较多的5种寄主植物,其中半寄生植物澜沧江寄生的寄主树为柚(Citrus maxima)、使君子(Quisqualis indica)、可可(Theobroma cacao)、木奶果(Baccaurea ramiflora)和红毛丹(Nephelium lappaceum);五蕊寄生的寄主树为柚、杧果(Mangifera indica)、海红豆(Adenanthera microsperma)、菩提(Ficus religiosa)和云南蕊木(Kopsia officinalis);全寄生植物金灯藤的寄主植物主要有红背桂(Excoecaria cochinchinensis)、木薯(Manihot esculenta)、树胡椒(Piper aduncum)、臭牡丹(Clerodendrum bungei)和银合欢(Leucaena leucocephala)。
1.3 样品处理
植物样品清洁后于烘箱内杀青,然后65 ℃烘干48 h至恒重。叶片和枝条的粉碎分别采用高速万能粉碎机(FW80型,天津市泰斯特仪器有限公司,天津)和微型植物粉碎机(FZ102型,天津市泰斯特仪器有限公司,天津)。C、N含量使用碳氮分析仪 (Vario MAX CN, Elementar Analysensysteme GmbH, Hanau,Germany)测定,P含量使用电感耦合等离子体原子发射光谱仪(iCAP6300, Thermo Fisher Scientific,Waltham, USA)测定。
1.4 数据分析
通过Excel 2016软件处理数据,利用SPSS 25.0软件进行独立样本T检验和Pearson相关性分析。使用SigmaPlot 12.5软件制图。
不同寄生植物化学计量的内稳性指数采用以下模型计算[8,29]:y=cx1/H
经对数转换可将内稳性指数(H)表示为:H=lgx/(lgy–lgc)
模型中,c为常数,x为环境营养物质的供应量,y为有机体的元素含量,x和y为浓度百分比或元素含量的配比,如P%、N%或N:P等。
2 结果与分析
2.1 寄生植物C、N、P化学计量及其季节变化特征
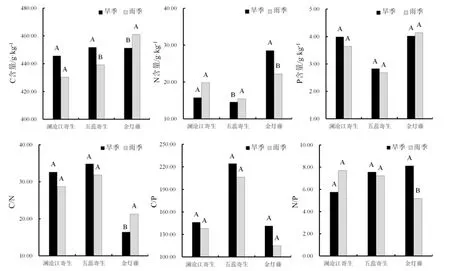
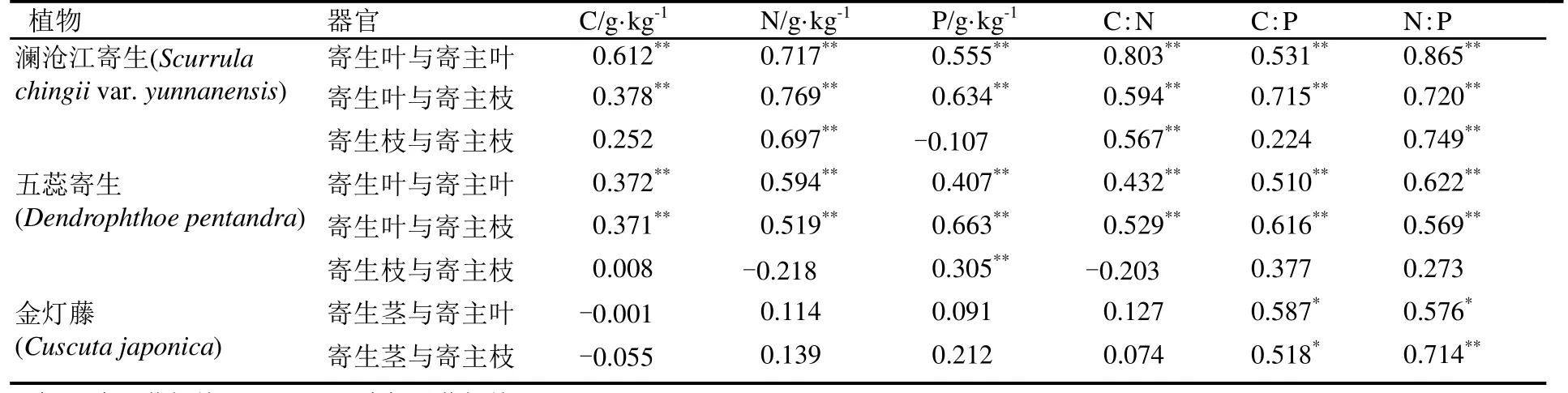
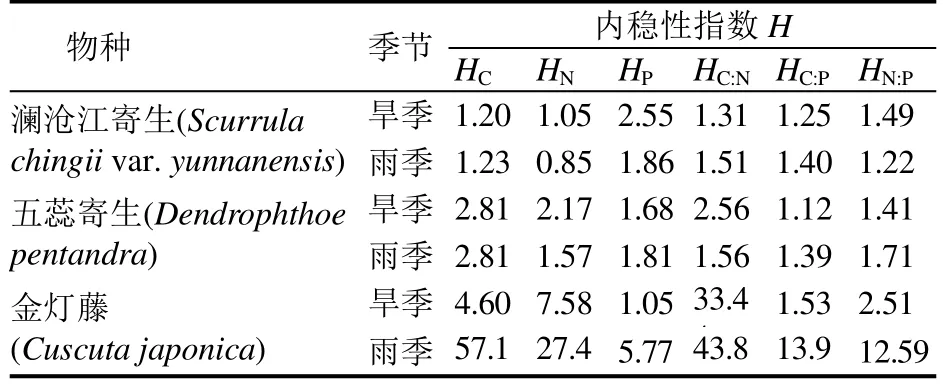
对西双版纳地区澜沧江寄生、五蕊寄生两种半寄生植物叶片和全寄生植物金灯藤茎的C、N、P进行化学计量分析。结果表明,在不同寄生类型中,全寄生植物金灯藤茎的 C、N含量均显著高于半寄生植物叶片,而半寄生植物五蕊寄生叶片的P含量显著低于半寄生植物澜沧江寄生和全寄生植物金灯藤;除 N:P没有显著差异外,半寄生植物叶片的C:N和 C:P均显著高于全寄生植物(表1)。对半寄生植物叶片和全寄生植物茎的C、N、P化学计量变异系数的分析显示,半寄生植物叶片中C元素含量的变异系数小于 N、P的变异系数,而化学计量比的变异系数呈现C:N 不同类型寄生植物的C、N、P化学计量的季节变化有所不同(图 1)。在旱季,澜沧江寄生叶片的C:P、N:P显著低于五蕊寄生;澜沧江寄生叶片和金灯藤茎之间N含量、C:N和N:P均存在显著差异;金灯藤茎的 N含量显著高于五蕊寄生,C:N、C:P也存在显著差异。在雨季,澜沧江寄生叶片 N、P含量显著高于五蕊寄生;金灯藤茎的C含量显著高于澜沧江寄生,各计量比也差异显著;金灯藤茎的C、N、P含量显著高于五蕊寄生叶片,但五蕊寄生叶片的C:N、C:P、N:P显著较高。 在半寄生植物中,五蕊寄生叶片的C含量在旱季显著高于雨季,N含量在旱季显著低于雨季,P含量和化学计量比的季节差异不显著;澜沧江寄生的C、N、P含量及其计量比的季节差异皆不显著。金灯藤C含量在旱季显著低于雨季;N含量和N:P在旱季显著高于雨季;C:N在雨季显著高于旱季;P含量和C:P在旱季与雨季之间没有显著差异。 表1 半寄生和全寄生植物C、N、P含量与化学计量及其变异系数Table 1 The contents of C, N, P and stoichiometry and their coefficient of variation in hemiparasitic and holoparasitic plants 图1 半寄生植物叶片和全寄生植物茎的化学计量及其季节变化特征Fig. 1 Stoichiometry and seasonal variation of leaves and stems of hemiparasitic plants and holoparasitic plants 两种半寄生植物叶片和全寄生植物金灯藤茎的P含量和C:N均显著高于寄主植物叶片,三种寄生植物的N含量和C:P、N:P则显著低于寄主植物叶片(表 2)。 西双版纳地区两种常见半寄生植物叶片与寄主叶、枝间的元素含量及化学计量比具有极显著的正相关关系(P<0.01),半寄生枝与寄主枝之间部分相关关系未达显著水平(P>0.05)。全寄生植物金灯藤茎与寄主植物枝、叶之间的C:P和N:P呈现显著的正相关(P<0.05),而C、N、P含量和C:N之间的相关性不显著(P>0.05) (表3)。 在寄生–寄主植物复合体中,半寄生植物寄生于寄主植物主干以外的侧枝上,则连续体内水分、无机盐、有机质等物质的运输途径为土壤–寄主主干–寄主枝条–寄生枝–寄生叶。在两种半寄生植物–寄主复合体中,虽然同一物种在不同寄主上有所差异,但寄主枝–寄生枝–寄生叶连续体的生态化学计量特征趋势具有高度相似性。澜沧江寄生和五蕊寄生中N含量及C:P、N:P在该连续体中总体呈先下降后增加的趋势;C和C:N含量总体呈现先增加后下降的趋势;P含量则在该连续体中不断富集(表4)。 表2 半寄生叶片和全寄生茎与其寄主植物叶片C、N、P化学计量的差异Table 2 Differences in stoichiometry of C, N and P in hemiparasitic leaves and holoparasitic stems and hosts’ leaves 表3 寄生植物与寄主叶、枝间生态化学计量特征的相互关系Table 3 The interaction of stoichiometry between parasitic plants leaves, host main branches and those of their host 表4 两种半寄生植物寄主枝-寄生枝-寄生叶中生态化学计量特征的变化Table 4 Changes of stoichiometric characteristics in the host branches, parasitic branches, parasitic leaves of two hemiparasitic plants (续表4) 3种寄生植物不同季节化学计量的内稳性指数(H)均大于1(表5),表明3种寄生植物能够维持内稳性,具有较强的控制养分的能力。除旱季的HP外,全寄生植物金灯藤的其他内稳性指数均大于两种半寄生植物。同一寄生植物中,澜沧江寄生的HP较大,HN较小;五蕊寄生的HC较大,HC:P较小,两种半寄生植物的内稳性指数季节变化不明显;金灯藤的HC、HC:N较高,HP较低。值得说明的是,雨季金灯藤的内稳性指数显著高于旱季。 表5 寄生植物叶片的内稳性特征及其季节变化Table 5 The homeostasis indexes of parasitic plants 本研究中半寄生植物叶片 C、N含量分别为441.75 g·kg-1和 15.96 g·kg-1,均低于全球陆生植物的平均水平[30],而半寄生植物叶片 P含量(3.28g·kg-1)高于全球[30]和全国陆地植物的平均水平[31],半寄生植物 N:P仅为 7.08,远低于全球的 13.8与全国的16.3[30—31]。与云南哀牢山亚热带山地森林常见半寄生植物椆树桑寄生(Loranthus delavayi)和柳叶钝果寄生(Taxillus delavayi)相比[21],本研究两种半寄生植物的C含量和C:N、C:P偏低,而N、P含量及N:P较高。全寄生植物金灯藤的 N(24.79 g·kg-1)与P(4.08 g·kg-1)含量均高于全球与全国水平,但其N:P(6.70)远低于全球与全国水平[30—31]。作为构成细胞的基本元素,C元素在生物体内具有含量高、变异小的特点[32—33]。本研究中,半寄生植物中C含量的变异系数小于10%,全寄生植物中C含量的变异系数为22.14%,亦验证这一点;半寄生植物中C:N变异系数小于 C:P,可能受植物体内 N、P含量变化、生物代谢等方面的影响[34]。植物叶片的养分含量随季节变化而存在较大差异[35],与年平均降水量相关性不显著[36]。我国热带雨林植被中开展的研究也未发现生态化学计量特征与降雨量之间存在相关性[37]。本研究的寄生植物C、N、P含量及其计量比大部分在旱季、雨季差异不显著。 全寄生植物金灯藤的 N、P含量显著高于半寄生植物,这与不同物种自身生物学特性导致的对养分选择吸收特性有关外[38],还与两者从寄主摄取养分的组成和数量差异相关,养分来源限制与 N、P利用效率均可能导致 N、P含量的差异。澜沧江寄生和五蕊寄生同为多年生桑寄生科植物,但两者的物候节律有所不同,生态化学计量特征的较大差异可能受生长发育阶段或物候节律等方面的影响[3]。叶片 C:N、C:P比值表明植物碳同化的能力[4],反映植物N和P的利用效率。半寄生植物具有部分合成有机物的能力,在营养元素供应缺乏的情况下往往具有较高的养分利用效率[39]。两种木本半寄生植物叶片C:N、C:P均显著高于全寄生植物草质茎,而N:P没有显著差异,这与不同功能群的相关研究基本一致[40],也反映出半寄生植物 N、P的利用效率高于全寄生植物,但又与普通高等陆生植物有较大差异,寄生植物这一独特的现象有待深入研究。 寄生植物较其寄主有更高的营养浓度[14],半寄生植物的N、P含量是寄主的2~4倍[41—44]。在本研究中,寄生植物叶片较寄主植物N含量和C:P、N:P显著偏低,P含量和C:N显著偏高。这可能是元素的单向流动或富集效应的原因[44]。半寄生植物叶片与寄主枝条的化学计量关系强于全寄生植物与寄主枝条的关系。半寄生植物叶与寄主叶之间、与寄主枝间的元素含量及化学计量比具有极显著的正相关关系,寄生枝与寄主枝C、P含量和C:P相关关系未达显著水平。全寄生植物寄生茎与寄主枝、寄生枝与寄主叶间C:P和N:P均存在显著正相关关系。前人研究表明,寄主植物中元素浓度能预测半寄生植物元素浓度[21],这一结论适用于半寄生植物,但并不适用于全寄生植物。 由于生长策略差异,不同物种的化学计量内稳性具有不同的变化趋势[35],常量元素内稳性高于微量元素[21],陆生植物N的内稳性绝对值(HN)强于P的内稳性绝对值(HP),而N:P的内稳性绝对值(HN:P)强于HN和HP[35]。在本研究中,半寄生和全寄生植物C的内稳性(HC)均大于N(HN)和P(HP)的内稳性,但在澜沧江寄生和五蕊寄生两种半寄生植物中,除五蕊寄生在旱季外,HN均小于HP,同时也不符合HN:P大于HN和HP的规律;全寄生植物金灯藤符合HN大于HP,HN:P大于HP的规律,但HN:P小于HN。植物生态化学计量内稳性除受光照强度、养分供应状况影响外,也受物种、器官(地上和地下)、生长发育阶段等因素影响[45]。在繁殖阶段HP较高是由于植物需要稳定的P供给来维持其生殖生长[29]。本研究中,旱季采样时五蕊寄生进入生殖生长,而澜沧江寄生处于营养生长阶段,但五蕊寄生表现出较低的HN:P。 内稳性高的物种往往具有更高的稳定性和优势度[8]。本研究中,金灯藤的内稳性高于半寄生植物的内稳性,在雨季更为明显。内稳性高的物种养分利用较为保守[46],在一定程度上也说明金灯藤具有较强的入侵能力[47]。因此,在未来的工作中需进一步研究不同类型寄生植物的养分利用策略与生态适应性。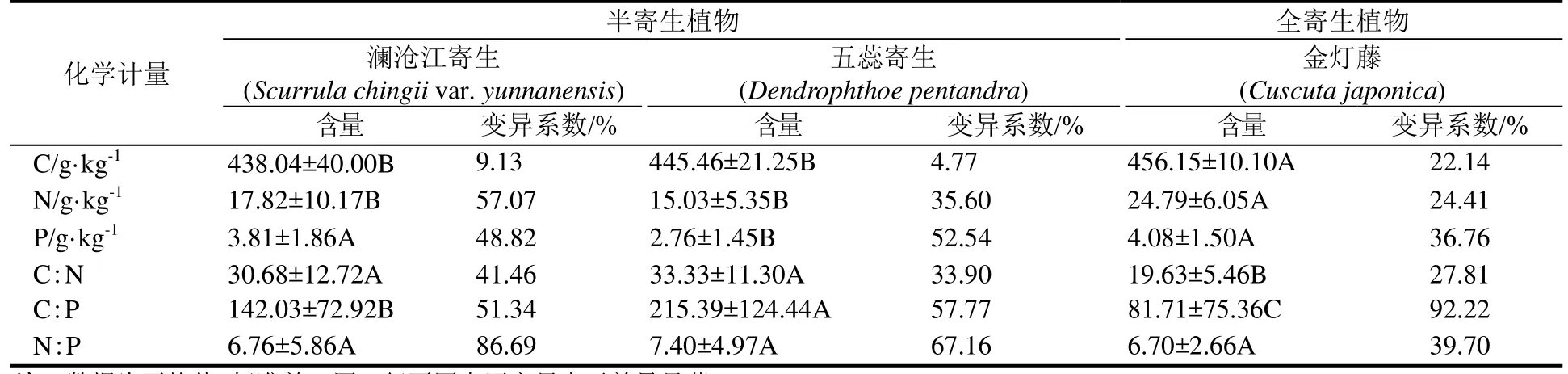
2.2 寄生植物及其寄主化学计量特征的相互关系

2.3 寄生植物的内稳性特征及其季节变化
3 讨论
猜你喜欢
落叶果树(2021年6期)2021-02-12
计测技术(2020年6期)2020-06-09
文苑(2020年12期)2020-04-13
海峡姐妹(2019年1期)2019-03-23
特别健康(2018年4期)2018-07-03
消费导刊(2017年24期)2018-01-31
江苏农业科学(2017年7期)2017-05-23
现代企业(2015年2期)2015-02-28
山东农业科学(2014年9期)2015-01-07
植物营养与肥料学报(2010年3期)2010-11-16
