
东北日光温室葡萄园水热通量特征及其对气象因子的响应
2020-09-22 09:03郑思宇王铁良魏新光付诗宁
干旱地区农业研究 2020年4期
郑思宇,王铁良,魏新光,李 波,孙 君,葛 东,付诗宁
(沈阳农业大学水利学院,辽宁 沈阳 110866)
生态系统水热传输过程与植物生理生态过程、环境特征密切相关[1],研究地表水热通量特征是探明生态系统与大气间相互作用的关键[2],也是合理制定并优化灌溉指标的重要参考指标,对合理分配及高效利用水资源至关重要[3]。东北地区是我国重要的农业生产基地,也是中国日光温室的起源地,日光温室面积占设施农业面积的50%以上[4],近年来也在大力发展温室果树产业。其中,辽宁省温室葡萄栽培面积达2.19×104hm2,已经成为当地设施农业主要的生产形式与经济来源[5]。水分管理是保证葡萄优质高产的关键环节,特别对温室而言,不合理的用水方式会造成水分利用率降低,还会导致温室内病虫害发生机率增大[6],从而影响葡萄品质及产量。
目前,关于研究植物蒸散发及能量分配的研究很多,比如有玉米[7-9]、水稻[10-11]、小麦[11]等,其中,对葡萄园水热特征研究也非常多。Yunusa等[12]利用波文比系统研究了澳大利亚地区滴灌葡萄园的水热通量变化规律,发现在2月中旬,可用净辐射分别被感热通量(H)、潜热通量(λET)与土壤热通量(G)均分,而到了3月末,H占比会下降到38%。Zhang等[13]研究荒漠绿洲葡萄园发现,不同生长阶段潜热通量与净辐射比率(λET/Rn)、感热通量与净辐射比率(H/Rn)、土壤热通量与净辐射比率(G/Rn)变化范围分别为29%~38%、57%~65%、4%~7%。Heilman等[14]研究表明葡萄冠层结构会对能量分配产生影响,H/Rn占冠层能量平衡较大部分。Kool等[15]对以色列干旱地区灌溉葡萄园水热通量进行研究时发现,H与λET之间相差不大,最大值在130~300 W·m-2之间变化,这期间的H/λET的值取决于上次灌溉以来的时间长短且灌溉会导致H快速下降,当日灌溉时,H最高达到64 W·m-2,第二天达到191 W·m-2,在之后不灌溉最高达到256 W·m-2。以上对葡萄园的研究都集中于大田环境,对温室研究较少。对温室中能量平衡及能量分配的研究多集中于蔬菜。Qiu等[16]对温室辣椒进行研究发现,在不同滴灌条件下,辣椒全生育期白天H/Rn均高于λET/Rn,白天G/Rn则较低,最高仅为7.9%,此外还得出λET/Rn与叶面积指数(LAI)呈线性增加关系。龚雪文[17]在对温室番茄研究时发现在不同生育期各能量分量占比有所不同,且具有季节性变化。从以上研究可知,因为影响能量分配及植物蒸散发因素较多(土壤含水率,叶面积指数,栽培模式等),所以导致各地区植物之间水热通量差异较大。况且,温室内净辐射要低于外部环境,空气流通性较差,风速低且空气湿度较大,会导致能量分配与蒸散发更为复杂[16]。目前也少见针对东北日光温室条件下葡萄水热传输的研究。因此,本文对东北日光温室条件下,葡萄水热能量分配及其对环境因子的响应作初步探索,以期为该地区温室葡萄节水灌溉提供理论依据。
1 材料与方法
1.1 试验区概况
试验于2018年5月1日—11月4日在沈阳农业大学试验基地辽沈Ⅲ型日光节能温室(N41.82°,E123.57°)内进行,温室坐北朝南,脊高4 m,跨度8 m,长度60 m,温室设计采用单面采光抛物面式,拱形钢桁架结构,温室内无支柱。前屋面覆盖材料为PVC防老化塑料无滴膜,采用防雨被作为保温措施。供试土壤0~60 cm土层容重为1.44 g·cm-3, 田间持水率为0.223 cm3·cm-3。
供试葡萄品种为“玫瑰香”(VitisviniferaL. cv. Muscat Ham-burg),于2015年3月完成定植,于4月25日施入基肥(鸡粪干含有机质62%,粗蛋白质30%,氮磷钾8%左右),施用量为3.3×104kg·hm-2。在新梢生长期(DOY131~162)、开花坐果期(DOY163~192)主要以施用氮肥(尿素,含氮量为42%)为主,在果实膨大期(DOY193~221)以及果实着色成熟期(DOY222~264)以施用复合肥为主,落叶期(DOY265~309)基本不施肥。N、P2O2、K2O折合为有效肥量分别为200、140、160 kg·hm-2。为防止土壤无效蒸发,进行覆膜处理。灌溉方式为自动灌溉,使葡萄全生育期土壤含水率控制在田间持水量60%~80%,当葡萄水势低于下限阈值时开始灌溉,当达到上限阈值时停止灌溉。采用Y型棚架,棵间距为0.5 m,行间距为3 m,藤长为3.5~4.5 m。
1.2 观测指标与测定方法
1.2.1 生态指标的测定 叶面积采用人工测量法,在整个试验期间随机选取10株葡萄标记新梢,测量枝条上所有叶子的长度及最大宽度。此外,随机摘取大小不一20片的叶子,记录叶长及最大叶宽,并进行拍摄,用Image J软件测量葡萄准确面积,建立叶面积与叶长、最大叶宽的回归关系。利用回归关系对枝条上葡萄叶面积进行估算,计算出相应葡萄植株的总叶面积,用叶面积投影除以冠层投影面积获得植株叶面积指数(LAI)。测量频率为7~10 d。
1.2.2 葡萄液流的测定 采用DynagageFlower32-1K系统(Dynamax Inc., Houston, TX, USA)对葡萄植株液流动态进行监测,为防止与外界热量发生交换,用锡箔纸对传感器部位进行包裹。为了防止连续加热对植株造成伤害,每10~15 d将传感器卸下并进行维护。在温室中选择8株长势一致植株进行测定,被测树体直径为19.6 cm±1.5 cm。用CR1000 型数据采集器进行数据的采集,采集频率为30 min·次-1。液流计算公式为:
(1)
式中,Fr为茎流瞬时速率(g·h-1);Pin为输入的热量(W);Qr为径向散热(W);Qv为竖向导热(W);Cp为水的比热;dT为竖向两个热电偶电压和的平均值(℃)。
1.2.3 水热平衡分量及气象数据的测定 温室中的气温(Ta,℃)、相对湿度(RH,%)采用位于温室中部的自动气象站(Campbell Scientific, Inc., Logan, UT, USA)测定,安装高度为2 m。水汽压亏缺(VPD,KPa)由空气温度与相对湿度计算可得[18]。葡萄冠层上方的净辐射(Rs,W·m-2)采用净辐射计(Model TBB-1,锦州阳光气象科技有限公司,中国)进行测定。土壤热通量(G,W·m-2)用2个UKSEFLUXHFP01热通量板测定,分别埋设距葡萄根部30 cm处,垂直深度为5 cm。使用CR1000型数据采集器进行采集,采集频率为30 min·次-1。
1.2.4 潜热通量及感热通量的计算 植物腾发过程受植被表层上的能量变化控制,忽略由平流作用产生的能量以及光合作用及各种能量转换,能量方程可表示为[10,17]:
Rn-λET-H-G=0
(2)
式中,λET为潜热通量(W·m-2);H为显热通量(W·m-2)。
由于试验为覆膜处理,可忽略土壤蒸发,即:
(3)
式中,λ为水的汽化潜热(J·kg-1);A为葡萄蒸腾面积(株·m-2);Fr为茎流瞬时速率(g·h-1)。
潜热通量(H)采用能量平衡法计算。
1.3 数据统计与分析
试验数据采用Microsoft Excel进行整理和计算,使用SPSS 19.0对数据进行统计分析,使用Origin 9.0绘图。
2 结果与分析
2.1 葡萄生态指标及日光温室萄萄园内环境变化
图1为试验期间葡萄生态指标及温室环境日变化。由于设备故障等原因导致DOY187~190、DOY199~212数据缺失。其中,图1a为全生育期葡萄叶面积指数及日均液流变化。利用叶面积指数和DOY之间进行拟合,拟合方程系数达0.98。在新梢生长期及开花坐果期LAI增长迅速,在果实膨大期及果实成熟期LAI保持相对较高,最高值达到2.64 m2·m-2。在落叶期葡萄叶脱落不严重,落叶期LAI仍相对较高,直到11月中下旬葡萄藤上仍存在许多叶子。葡萄液流日变化具有季节性,新梢生长期、开花坐果期、果实着色成熟期、落叶期日均液流量分别为37.79、61.99、46.38、27.01 g·h-1,由于果实膨大期数据缺失较为严重,并不做统计。
图1b、c为全生育期温室内主要气象因子(Rn、Ta、RH、VPD)日均变化。在试验期间Rn变化范围为33.34~129.76 W·m-2,平均值为61.65 W·m-2。VPD的变化趋势与Rn相似,变化范围为0.38~1.77 kPa,均值为0.71 kPa。从图中可以看出,在开花坐果期到果实着色成熟期,温室内Ta较高,日均值最高为33.48℃。相对其他3个气象因子来说,在整个生育期RH均保持在较高的水平,最大值达到了60.24%。
2.2 日光温室葡萄园内能量分配
选取各生育阶段典型日5~7 d,将同时段的Rn、λET、H、G进行平均,得到温室葡萄园的平均水热通量日变化,如图2所示。从图中可以看出,能量平衡各分量总体上呈现为单峰变化趋势,一般白天均为正值,夜间为负值。但不同生育期分量正负转换时间不同,这主要是由于生育阶段环境不同导致的[17]。Rn的峰值出现在12∶00—13∶00左右,峰值有不同程度波动,波动范围为110.93~618.75 W·m-2,这种变化主要是受太阳高度、天空云量以及下垫面作物生长情况等影响[19]。λET最大值波动范围是42.24~242.73 W·m-2,H最大值波动范围是56.57~327.93 W·m-2,λET与H变化规律与净辐射变化规律一致性较好,随着净辐射的增加而增加。在新梢生长期,λET的值较小且远远小于H,平均最大值为35.86 W·m-2;在其余生育期λET与H差别并不是很大,且果实着色成熟期期λET峰值大于H峰值,二者峰值分别为265.81、229.72 W·m-2。G是净辐射另外一个重要的能量支出,其变化相对平缓,在夜晚一般为负值。各生育期平均G分别为7.76、-0.3、6.48、1.94、3.69 W·m-2。可以看出,各生育阶段G大多数都低于λET和H,其滞后现象并不明显。

图1 温室内葡萄液流、生理指标(LAI)及主要环境因子(Ta, Rn, RH, VPD)日均变化Fig.1 Average daily variation in sap flow, physiological index (LAI) of grapevine and mainenvironmental factors (Ta, Rn, RH, and VPD) in greenhouse
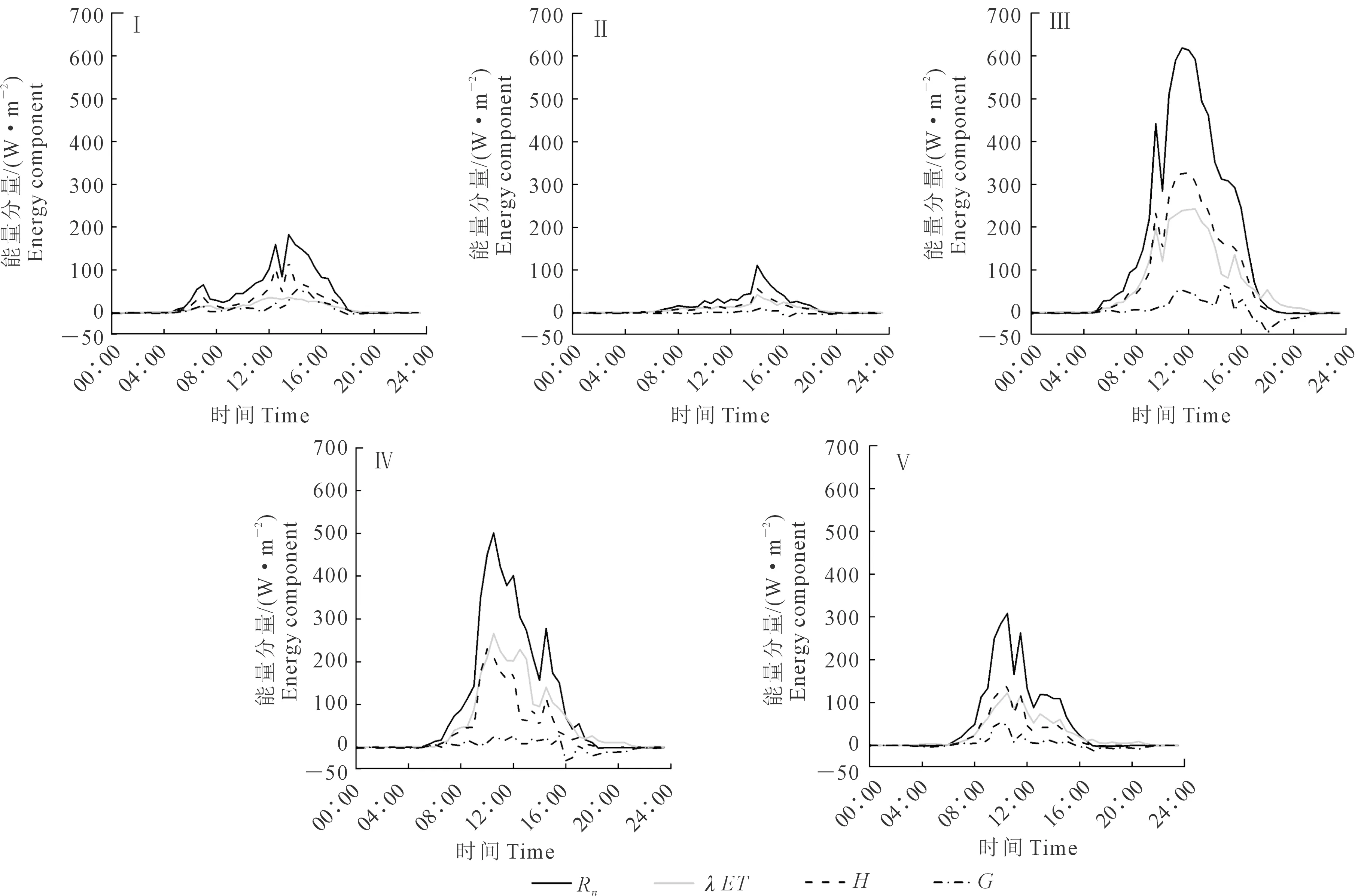
注:Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ分别代表新梢生长期、开花坐果期、果实膨大期、着色成熟期、落叶期。下同。Note: Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ represent the growth stage of new shoots, the flowering and fruiting stage, the fruit expansion stage,fruit coloring and fruit maturity stage and the leaf fall stage, respectively. The same below.图2 温室葡萄不同生育期水热通量日变化Fig.2 Daily variation of water and heat fluxes of greenhouse grapevine at different growth periods
表1与图3分别为温室内葡萄各生育期白天能量分量的平均值变化及日均能量比率的季节变化。从表1可知,白天Rn与λET的值在新梢生长期及落叶期较小,新梢生长期、落叶期Rn的值分别为120.87、110.65 W·m-2,λET为34.55 W·m-2和42.5 W·m-2。在果实着色成熟期,白天Rn与λET的值较大,分别为177.70、78.49 W·m-2。白天λET/Rn在着色成熟期最大,为44.17%,在新梢生长期最小,为28.58%。然而H/Rn在新梢生长期最大,达到为62.31%,且随着生育进程有下降趋势。在新梢生长期及开花坐果期,G/Rn同样表现为最大,分别为9.10%、12.32%。由图3仍可以看出在整个生育期比率大小依次为H/Rn>λET/Rn>G/Rn。λET/Rn季节性变化处于11.23%~68.32%之间,H/Rn处于26.34%~86.31%之间。出现以上结果的原因可能是因为在葡萄生育初期,叶子稀疏,地表裸露面积较大,冠层表面接收能量较小,而随着葡萄生长冠层面积逐渐增大。此时,冠层截获光能力变强,λET逐渐增加,H与G逐渐减小,但H仍是净辐射的最大占比通量[2]。
2.3 日光温室葡萄园潜热通量与环境因子的关系
图4为温室内葡萄各生育期瞬时潜热通量和气象因子的相关关系,图5为各生育期瞬时潜热通量与气象因子的相关系数统计。从图中可以看出,太阳辐射对λET的影响最重要,太阳辐射强度的大小决定着叶片气孔开闭状态,气孔开度越大,则蒸腾作用就越强[20],两者呈现线性关系,各生育期R2值均较高,最高达到0.88。随着Ta的升高λET也升高,而随着RH增大λET有减小的趋势。Ta、RH分别与λET之间的相关系数表现为Ta>RH(见图5)。VPD在一定程度上反映了植株叶片中的水分进入空气中的难易程度,Ta、RH对λET的影响很大程度上是通过改变水汽压梯度来实现的,VPD和λET之间的相关系数与Ta和λET之间的相关系数相比相差并不大,变化范围为0.36~0.65,果实着色成熟期相关系数远大于Ta与λET的相关系数。日尺度上VPD与λET相关系数要大于Ta与λET相关系数。总的来说,Rn是影响潜热变化的最主要气象因子,其次为VPD、Ta、RH。各生育期Rn始终是λET最主要的控制因子。

表1 温室内葡萄不同生育期白天能量分量及分配规律

图3 日光温室葡萄园能量日均分配的季节变化Fig.3 Seasonal variation of daily average energy distribution of vineyards in solar greenhouse
对全生育期温室葡萄日均潜热通量与气象因子进行线性回归分析(见表2),发现λET与气象因子的关系与各生育期相似,但相关系数有所降低,Rn与λET相关系数为0.79,与RH的相关系数仅为0.28。这表明随着研究时间尺度的增加,气象因子对λET的影响在逐渐变弱。这主要是因为在全生育期尺度上,λET不仅受气象因子的影响,还受植株生长特性的影响。但气象因子与潜热通量的相关系数仍表现为:Rn>Ta>VPD>RH。
3 讨 论
对土壤-作物水热传输系统的研究是确定作物需水规律及需水量的重要参考部分,探讨水热传输规律是提高灌溉决策的核心。本文研究温室葡萄水热通量变化时发现,在生育初期,λET/Rn的值较小,随着葡萄发育λET/Rn逐渐增大,在果实着色成熟期达到最大,落叶期有所降低,这主要是因为LAI增大,导致作物蒸腾变大,这与Qiu等[16]得到的结果是一致的。但在整个生育期,H是净辐射最主要的能量支出,H/Rn达50%以上;余昭君等[21]在对葡萄园进行研究时也发现H/Rn的值在各个生育阶段都大于0.60。干旱荒漠绿洲葡萄园在生育后期潜热通量基本高于感热通量[19],而本研究发现,只有在果实膨大期潜热通量的平均最大值会高于感热通量,这点与Oliver等[22]研究结果一致,导致出现不同结果的原因可能是因为灌溉方式不同引起的,灌水量、灌溉时间等都会影响葡萄园的生物气候情况[14-15]。葡萄属于稀疏植被,冠层结构也会对能量分配产生一定的影响,这也可导致各地区研究结果的不一致[15]。土壤热通量在整个生育期都维持在相对稳定的水平,在落叶期土壤热通量较低,这是因为此时辐射较弱,温度有所降低,导致此时土壤热通量较小。潜热通量代表腾发部分,是温室葡萄能量主要消耗形式之一,通过回归分析表明,Rn是影响潜热通量最主要的气象因子,其次是VPD,这点与其他学者研究结果一致[2,11,17]。

图4 葡萄不同生育期潜热通量(λET)与气象因子的关系Fig.4 Relationships between λET and meteorological factors in different growth periods of grape
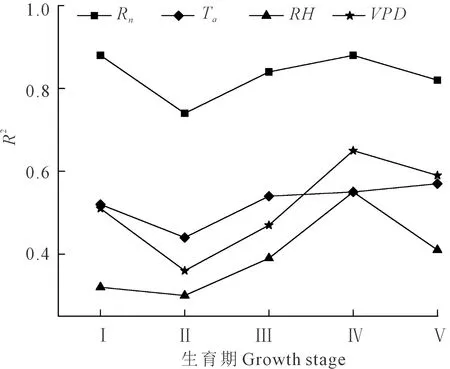
图5 葡萄不同生育期潜热通量(λET)与气象因子的相关系数变化Fig.5 Variation of correlation coefficient between λET andmeteorological factors in different growth periods of grape
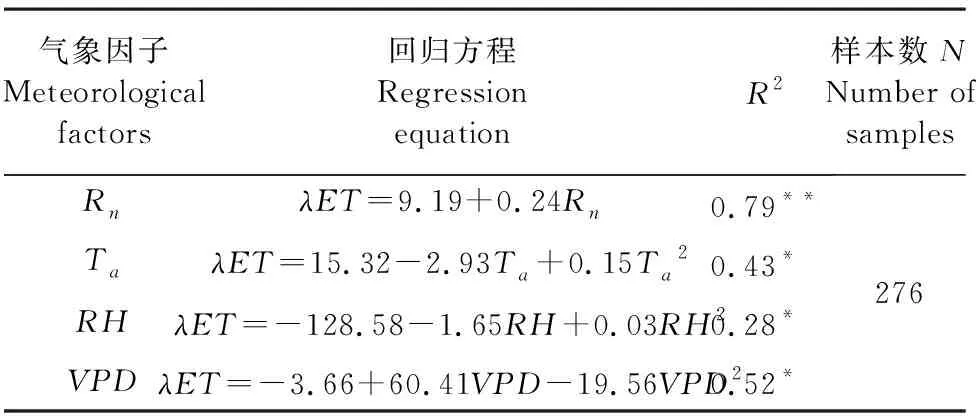
表2 温室葡萄日潜热通量与气象因子的线性回归
4 结 论
1)水热通量各分量在整个生育期日变化总体上呈现单峰趋势,Rn的峰值波动范围为110.93~618.75 W·m-2,λET峰值波动范围是42.24~242.73 W·m-2,H峰波动范围是56.57~327.93 W·m-2。在新梢生长期及落叶期,白天净辐射和潜热通量较小,随着生育期的推进潜热通量逐渐增大,在果实着色成熟期达到最大值78.49 W·m-2。显热通量在全生育期均较大。
2)白天潜热通量占净辐射的比值在新梢生长期最小,为25.28%,在果实着色成熟期最大,为44.17%。感热通量占净辐射比值整个生育期都达50%以上,土壤热通量占净辐射比值相对较小,最大仅为12.32%。在整个生育期H/Rn>λET/Rn>G/Rn。λET/Rn季节性变化处于11.23%~68.32%之间,H/Rn处于26.34%~86.31%之间。
3)通过分析气象因子(Rn、Ta、RH、VPD)对潜热通量的影响,结果表明:在不同生育阶段瞬时尺度上,Rn是影响潜热变化最主要的气象因子,R2最高达0.88。RH与潜热通量之间表现为负相关关系。与瞬时尺度相比,在日尺度上各气象因子对潜热通量的影响在逐渐变弱,RH与潜热通量相关系数仅为0.28。但无论从瞬时尺度还是日尺度来说,Rn仍然是影响潜热通量最主要的气象因子。各气象因子对潜热通量的影响为:Rn>Ta>VPD>RH。
猜你喜欢
农业工程学报(2022年6期)2022-06-27
遗传(2022年6期)2022-06-21
科学技术创新(2022年15期)2022-05-18
农业灾害研究(2022年1期)2022-05-07
环境(2021年5期)2021-06-20
暴雨灾害(2021年3期)2021-05-21
暴雨灾害(2021年2期)2021-04-02
科学与技术(2019年3期)2019-03-05
科学文化评论(2018年2期)2018-05-14
现代农业科技(2017年7期)2017-05-22
