
新疆冬小麦地方品种与育成品种基于SNP芯片的遗传多样性分析
2020-09-13 15:05马艳明娄鸿耀陈朝燕倪中福
作物学报 2020年10期
马艳明 娄鸿耀 陈朝燕 肖 菁 徐 麟 倪中福 刘 杰,*
新疆冬小麦地方品种与育成品种基于SNP芯片的遗传多样性分析
马艳明1,2娄鸿耀1陈朝燕1肖 菁2徐 麟2倪中福1刘 杰1,*
1中国农业大学农学院, 北京 100193;2新疆农业科学院农作物品种资源研究所, 新疆乌鲁木齐 830091
新疆是我国西北重要的小麦优势产区和消费区。解析新疆冬小麦地方品种与育成品种之间的遗传多样性和亲缘关系, 对新疆冬小麦育种中杂交组合的合理选配以及后代选择具有重要的指导意义。本研究利用小麦55K SNP (single nucleotide polymorphism)芯片对134份新疆冬小麦地方品种及54份育成品种进行全基因组扫描, 估算其品种间的遗传距离, 揭示其遗传多样性。结果表明, 所有 SNP 位点的多态性比率达到95.62% (50,743/53,063)。每条染色体分布1068~2616个多态性位点, 多态性标记在基因组间分布呈现 A>B>D。188个品种间的两两遗传距离在0.002~0.723之间, 平均为0.378。其中134个地方品种两两之间的遗传距离在0.002~0.400之间, 平均为0.070; 54个育成品种两两之间的遗传距离在0.004~0.337之间, 平均为0.114; 134个地方品种与54个育成品种之间的遗传距离在0.605~0.723之间, 平均为0.699。聚类结果显示可将所有材料分为10个不同类群。综合SNP和系谱分析, 育成品种与地方品种之间的遗传差异最大, 其次是育成品种之间, 而地方品种之间遗传差异最小。鉴于育成与地方品种之间较大的遗传差异, 新疆冬小麦品种可以利用地方种来丰富其育种的种质基础, 拓宽遗传背景, 进而提高当地小麦育种水平。本研究为新疆冬小麦品种选育和改良提供了重要的理论指导。
新疆; 冬小麦; 地方品种; 育成品种; SNP; 遗传差异
小麦(L.)是我国三大主粮作物之一, 种植面积广、总产量高[1]。新疆是我国西北重要的小麦优势产区和消费区, 冬小麦播种面积占全区小麦总面积的70%以上[2-3]。新中国成立以来, 新疆南北疆的冬小麦种植区都经历了“地方品种—引进品种—自育品种”的更替。事实证明, 小麦品种改良在增产中取得了巨大成效。然而, 随着小麦品种产量潜力不断提高, 新育成品种也出现了品种类型相似和抗病、抗逆性较差的问题, 在产量和适应性上没有大的突破[4-5]。这可能与育种过程中长期对产量、抗性、品质等性状的连续定向选择, 造成育成品种遗传基础日益狭窄、遗传多样性下降有关。遗传变异是遗传育种的基础, 认识和了解遗传变异将对育种过程中杂交组合的合理选配以及后代选择提供指导。因此, 非常有必要结合利用最新的分子标记技术手段, 对新疆的冬小麦地方品种和育成品种(系)及其育种亲本材料的遗传基础和亲缘关系全面深入了解, 为亲本选配、分子标记辅助选择和小麦育种实践提供应用与指导。
单核苷酸多态性(single nucleotide polymorphism, SNP)是美国学者Lander于1996年提出的第3代DNA遗传标记[6], 因与芯片技术和基因克隆相结合, 具有数量多、易于自动化检测分型和通量高等优点, 在小麦遗传图谱构建、基因定位、遗传多样性分析、农艺性状关联分析、分子标记辅助育种等方面发挥着重要作用。随着高通量测序技术的快速发展, 先后开发出小麦9K、90K、660K、55K等小麦SNP芯片, 其中小麦55K SNP芯片是由中国农科院作物科学研究所小麦基因组与基因资源研究团队在小麦660K SNP芯片的基础上开发的, 经过了数千份国内小麦品种材料的检验, 相较于9K、90K和660K芯片, 其有效标记的比例更高, 具有更高的利用价值[7-8]。目前利用小麦55K SNP芯片仅见于构建高密度遗传连锁图[6-7]以及关联分析方面[9-10], 尚未见有关利用小麦55K SNP芯片开展小麦种质遗传多样性研究的报道。
由于小麦品种具有很强的区域适应性, 在不同种植区具有明显的地区特征和习性, 各地区育种目标也不完全相同, 针对特定区域的小麦品种材料及育种实践开展有关研究非常必要。本研究利用小麦55K SNP标记技术揭示134个新疆本土冬小麦地方品种和54个新疆冬小麦育成品种的遗传多样性和亲缘关系, 并探讨新疆冬小麦品种在改良过程中基因多样性的变化趋势, 全面阐述新疆冬小麦种质的遗传特性, 为小麦育种实践提供指导和参考。
1 材料与方法
1.1 试验材料及DNA提取
选用188个新疆冬小麦品种资源, 其中地方品种134个, 育成品种54个, 均为新疆农业科学院农作物品种资源研究所收集保存的材料, 试验采用2015年10月种植, 2016年7月收获的自花授粉种子。全部试验材料于2016年10月在中国农业大学小麦遗传育种温室种植, 植株长至二叶一心时, 取小麦叶片, 按Saghai-Maroof等[11]的CTAB法提取基因组DNA。
1.2 基因型分析
利用中国农业科学院作物科学研究所小麦基因组与基因资源研究团队开发的小麦55K SNP芯片对188个品种进行全基因组扫描。小麦55K芯片包含55,000个左右SNP标记, 均匀分布在小麦21条染色体上, 每条染色体平均有2600个左右SNP, 标记间的平均遗传距离约为0.1 cM, 平均物理距离小于300 kb, 并具有完整而详细的基因组信息。
1.3 数据分析
针对SNP芯片的原始扫描结果, 利用Affymetrix的Axiom Analysis Suite软件分型。选择分型成功率(Call rate)>0.8、缺失率<10%、最小等位基因频率(MAF)>0.05过滤的50,743个SNP位点, 利用PowerMarker V3.25计算多态信息含量(PIC)和品种之间的遗传距离Nei[12]。根据Nei的遗传距离, 通过MEGA6软件(http://www.megasoftware.net/)构建遗传聚类树状图。
2 结果与分析
2.1 多态性SNP位点的分布
在分布于21条染色体的53,063个SNP位点中, 共计有50,743个SNP位点检测到多态性, 多态性比率为95.62% (50,743/53,063)。每条染色体分布1068~2616个多态性位点, 6D染色体上最少, 2A染色体上最多, 平均每条染色体2416个(图1-a)。除6D外, 染色体间标记数量差异不大(图1-a)。SNP标记在A、B和D基因组间分布比较均匀, A基因组的标记数最多, 有17,702个, 占34.88%; B基因组居中,有16,943个, 占33.39%; D基因组的标记数最少, 有16,098个, 占31.73%。多态性SNP位点在7个部分同源群中的分布数量在5696~7797之间, 第 3部分同源群最多, 第6部分同源群最少, 分布从多到少依次为3>2>4>5>1>7>6 (图1-b)。
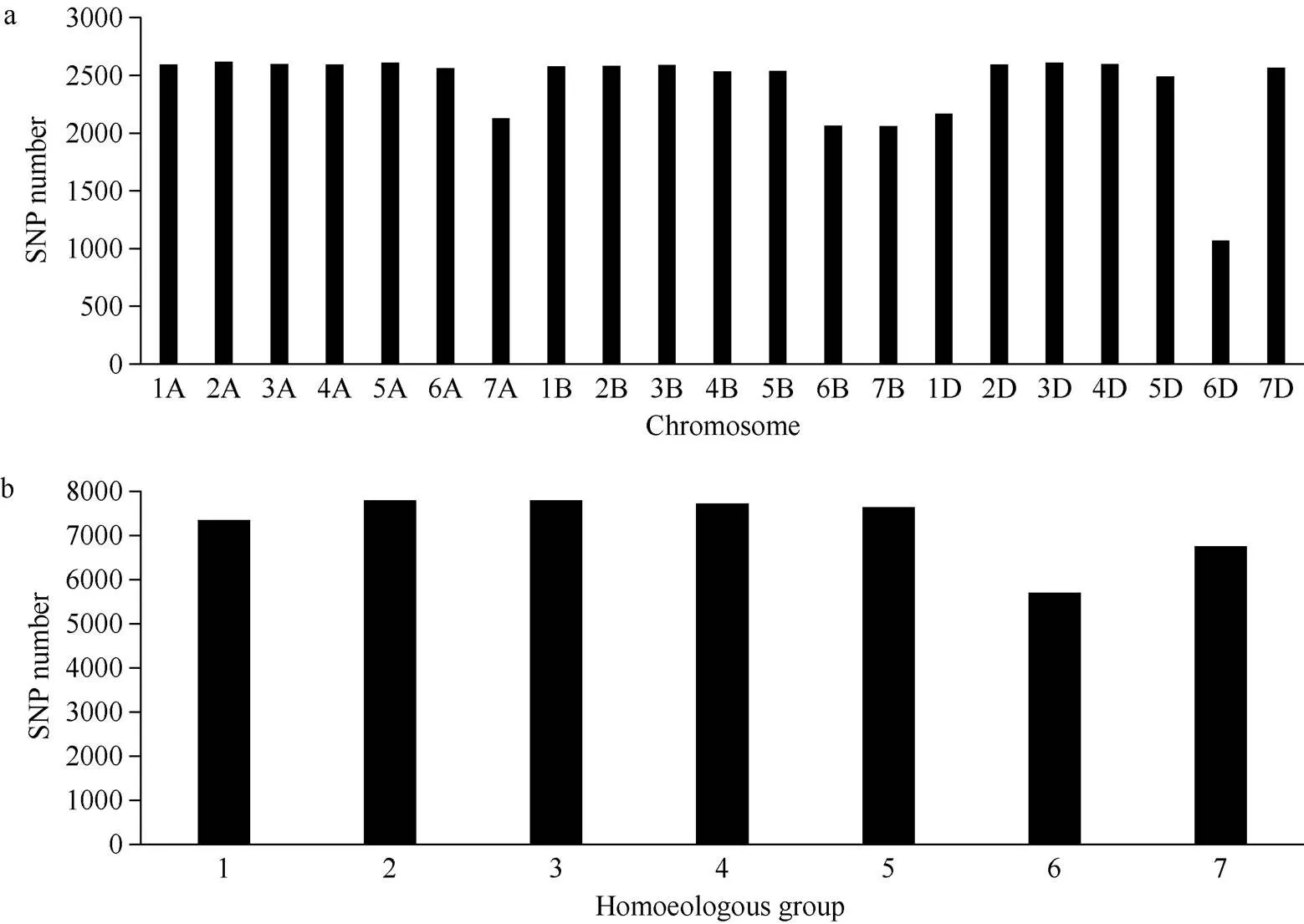
图1 多态性位点在染色体(a)和部分同源群(b)上的分布
2.2 SNP标记多态性信息含量及品种间的遗传距离
基于SNP标记数据计算品种的多态性信息含量(polymorphic information content, PIC)值, 变异范围在0.090~0.375之间, 平均为0.265。134个地方品种的PIC值平均为0.229, 最高0.374, 最低0.090; 54个育成品种的PIC值平均为0.299, 最高0.375, 最低0.099 (表1)。
利用SNP标记数据计算188个品种两两之间的遗传距离, 共获得17,580个遗传距离值, 范围在0.002~0.723之间, 平均为0.378 (表1)。巴州库尔勒市的名为阿克库斯格的地方品种与巴州若羌县的名为冬麦的地方品种之间的遗传距离最小(0.002), 喀什地区莎车县的地方品种白芒白冬麦与育成品种新冬7号之间的遗传距离最大(0.723)。
按地方品种、育成品种、地方品种与育成品种之间3组比较遗传距离(表1), 134个地方品种两两之间的遗传距离值在0.002~0.400之间, 平均为0.070, 巴州库尔勒市的名为阿克库斯格的地方品种与巴州若羌县的名为冬麦的地方品种之间的遗传距离最小(0.002), 博乐市的地方品种小红芒与和田市的地方品种卡衣木江之间的遗传距离最大(0.400)。134个地方品种中, 有26个来源地不同、品种名皆为“白冬麦”的地方品种, 其中7个来自北疆各地区, 2个来自东疆的哈密地区, 17个来自南疆各地区。26个“白冬麦”之间的遗传距离在0.003~0.263之间, 2个分别来自乌鲁木齐市与昌吉州呼图壁县的白冬麦之间的遗传距离最小, 实际这2个品种的来源地之间的地理距离也较小; 2个分别来自北疆昌吉州阜康县与南疆喀什地区叶城县的白冬麦之间的遗传距离最大, 实际这2个品种来源地之间的地理距离也较大。
54个育成品种两两之间的遗传距离值在0.004~ 0.337之间, 平均为0.114, 新冬11号与新冬12号之间的遗传距离最小, 巴冬1号与新冬20号之间的遗传距离最大。由系谱可知, 新冬11号与新冬12号的母本均为杂交组合热衣木夏/亥恩·亥德; 巴冬1号母本是新疆的冬小麦地方品种阿克库斯格, 父本是意大利引进冬小麦品种奥维斯特, 而新冬20是由河北省农林科学院育成的冀87-5018系选而来。
134个地方品种与54个育成品种之间的遗传距离在0.605~0.723之间, 平均为0.699, 说明新疆冬小麦地方品种与育成品种之间的遗传差异大于地方品种之间的遗传差异, 也大于育成品种之间的遗传差异。伊宁市的地方品种无芒麦与育成品种奎冬5号之间的遗传距离最小(0.605), 喀什地区莎车县的地方品种白芒白冬麦与育成品种新冬7号之间的遗传距离最大(0.723)。
以0.02为组距进行次数分布频数分析(图2), 188个品种间的遗传距离分布频数呈现2个波峰曲线, 一个是地方小麦品种之间以及育成品种之间的遗传距离分布频数曲线, 在0.1处密度最大, 主要分布在0.06~0.12之间, 占26.10% (4588/17,580); 另一个是地方品种与育成品种之间的遗传距离分布频数曲线, 在0.72处分布密度最大, 主要分布在0.68~ 0.72之间, 占36.39% (6397/17,580)。
2.4 聚类分析
根据所有小麦品种的SNP, 利用PowerMarker V3.25 得到188个小麦材料两两之间的遗传距离矩阵, 并构建了NJ聚类图(图3)。通过聚类, 我们将所有小麦材料分为10个不同类群, 其中第I类(47个品种)和第II类(7个品种)均为育成品种, 其余类群均为地方品种, 其中第III类有6个品种, 第IV类有5个品种, 第V类23个品种, 第VI类18个品种, 第VII类25个品种, 第VIII类29个品种, 第IX类24个品种, 第X类最少, 仅有4个品种。各类群及包含的小麦材料详见表3及表4。
第I类47个品种均为育成品种, 遗传距离在0.004~0.337之间, 分为3个亚类。第1亚类19个品种, 亲本多含有国外引进冬小麦品种, 如石冬6号与新冬26号, 均为中国科学院新疆生态与地理研究所选育的耐盐冬小麦品种, 且均以冬小麦品种红选501与墨西哥春麦品种唐努尔杂交的F1代为母本, 再与红选501和墨西哥春麦品种西埃代赛洛斯杂交的F1代为父本而育成的品种。最为突出的是新冬2号冬小麦品种, 是新疆农业科学院粮食作物研究所与新疆八一农学院(现新疆农业大学)以新疆冬小麦地方品种热衣木夏为母本、丹麦引进多花多实冬小麦品种亥恩·亥德为父本杂交选育而成, 于1966年审定命名[13]; 新疆农业科学院粮食作物研究所又以新冬2号姊妹系为母本、前苏联引进冬小麦品种奥德萨3号为父本, 进一步选育得到新冬6号、新冬7号、新冬10号3个品种。新冬号冬小麦品种曾在全疆推广种植, 对新疆小麦产量的提高起了很大作用, 其中新冬2号1980年种植23.3万公顷, 占全疆冬小麦面积的1/3。新疆农垦科学院以新疆冬小麦地方品种热衣木夏与前苏联引进品种新乌克兰83号杂交选育成新冬14号, 又利用新冬14号与河南安选5号杂交得到新冬17号小麦品种。第2亚类15个品种, 亲缘关系比较复杂, 其中奎冬5号与新冬22号均以加拿大引进小麦品种Nostar与花春84-1杂交F1为母本, 以高代品系76-4与引自罗马尼亚的洛夫林13杂交F1为父本选育而来; 喀冬1号、奎花1号、新冬15号、新冬16号、新冬18号均为利用我国新疆冬小麦地方品种(巴克甫克、热衣木夏、阿克苏红冬麦)与丹麦引进品种亥恩·亥德的杂交后代再与其他国内外冬小麦品种杂交选育而来, 如引自前苏联的无芒4号、小鹅186、阿芙乐尔、50周年等, 以及国内其他省份的西北134、华北187、京花1号、山东蚰包、冀麦2号等。第3亚类有13个品种, 均含有引自国内其他省份的冬小麦优良品种血缘, 如引自河北的冀6159、冀87-5018, 引自山东的PH82-2- 2、鲁植79-1, 引自陕西的西北612、山西的太谷核不育系以及庄巧生院士选育的优良品种华北187等。其中喀冬4号、新冬20号、新冬46号分别是华北187、冀87-5018、农大211引入新疆试种推广后审定的品种, 新冬25号的父母本均为河北的品种。
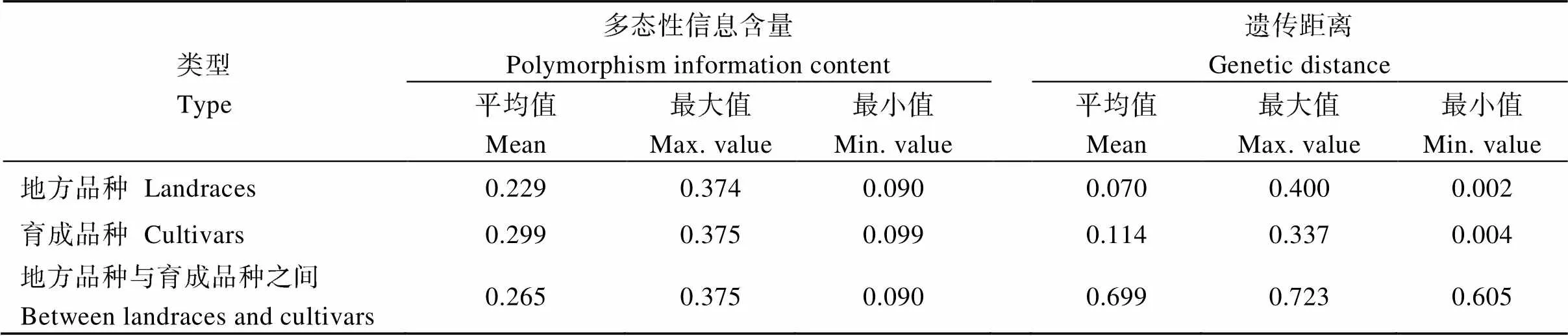
表1 地方品种和育成品种遗传差异比较
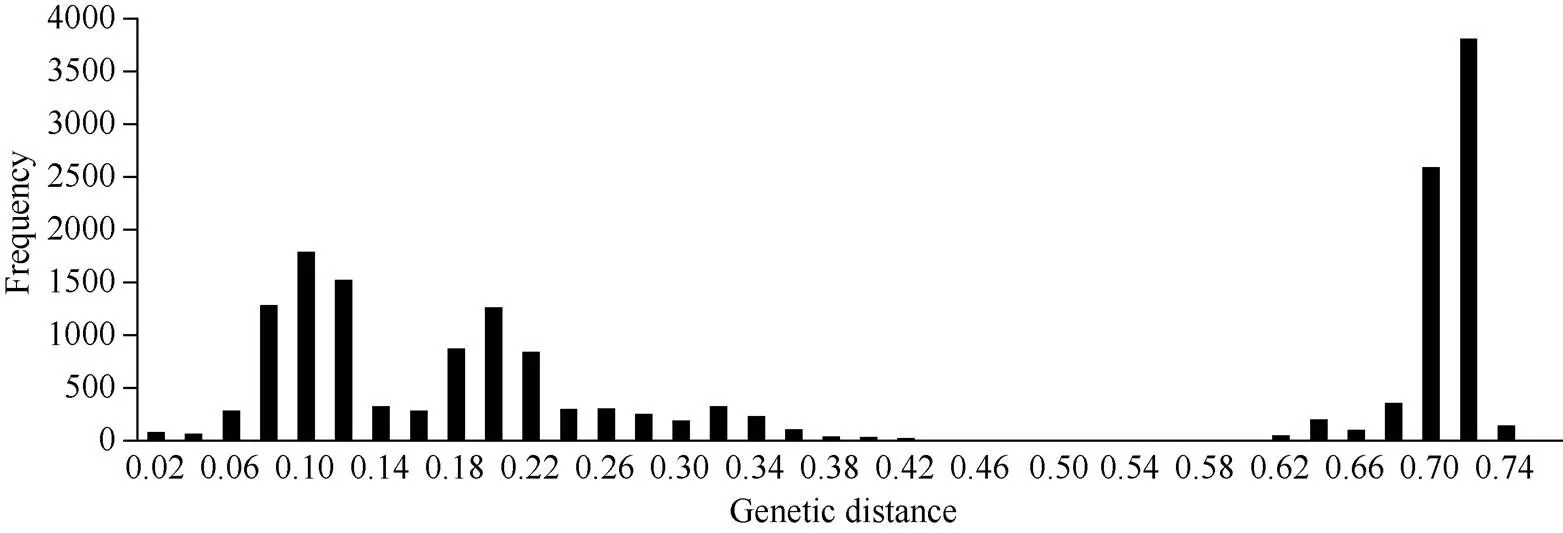
图2 品种间遗传距离的次数分布
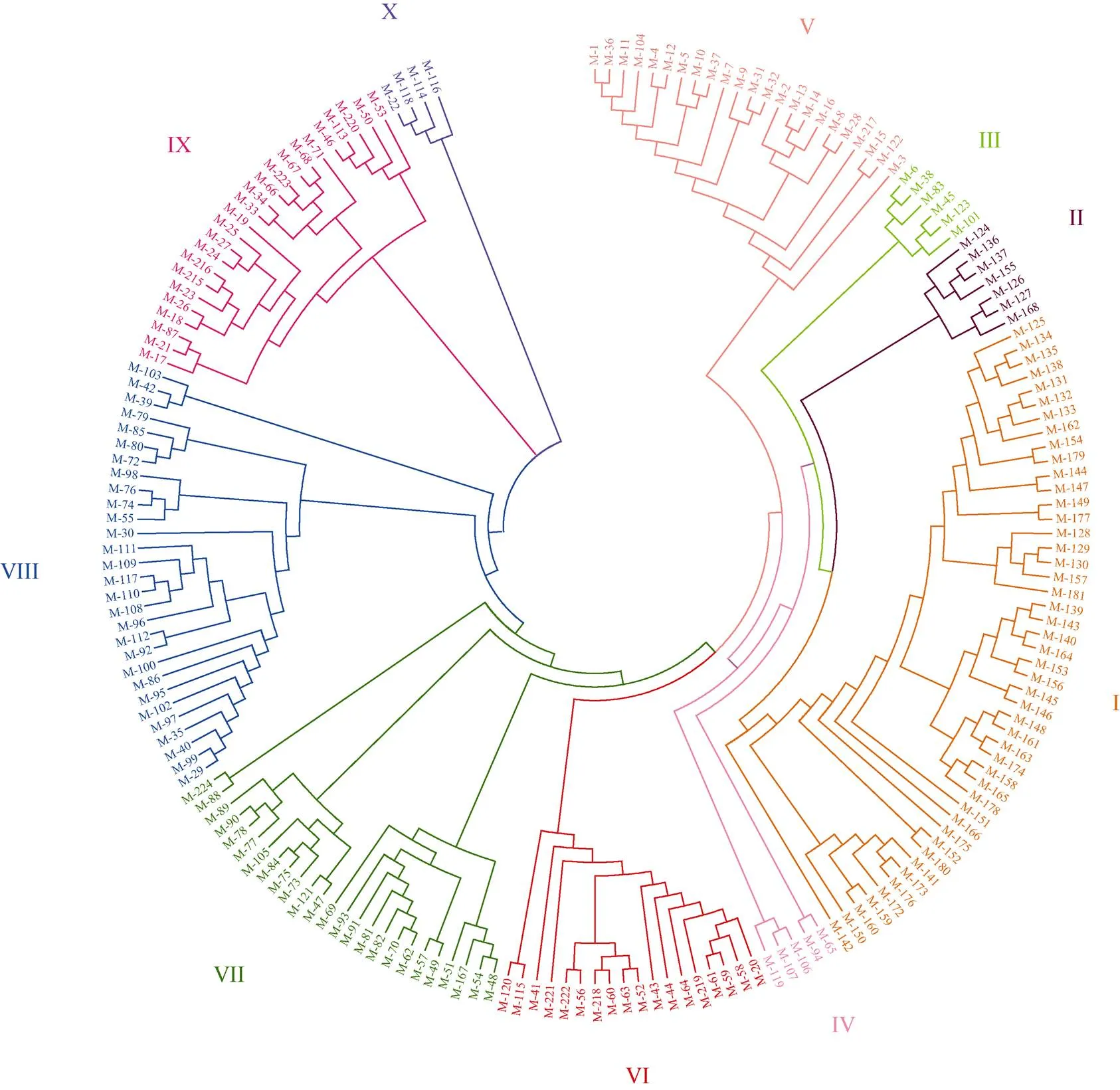
图3 188个新疆小麦地方品种和育成品种的SNP数据聚类图
代码的含义同表4。Codes correspond with those given in Table 4.
第II类7个品种均为育成品种, 遗传距离在0.006~0.269之间, 新冬2号与新冬3号之间的遗传距离最小, 喀冬2号与新冬13号之间的遗传距离最大。其中新冬2号与新冬3号的亲本之一都是上世纪50至60年代冬小麦育种多选的、引自丹麦的多粒源亲本亥恩·亥德, 新疆农业科学院粮食作物研究所又以品质表现较好的新冬3号为母本、较抗病抗寒的前苏联冬小麦品种乌克兰0246为父本进行杂交, 培育出了中产、品质好、容重高且抗病、抗寒性等都比新冬2号有所提高的新冬13号小麦品种。此亚类中巴州1号和巴州2号的共同亲本之一是引自意大利的冬小麦品种奥维斯特; 喀冬2号(乌克兰0246/叶城春麦)和石冬6号(春小麦京5029/辐射育种冬小麦品系77-21)的亲本之一都是春麦, 均具有早熟特性。

表3 各类群小麦品种列表
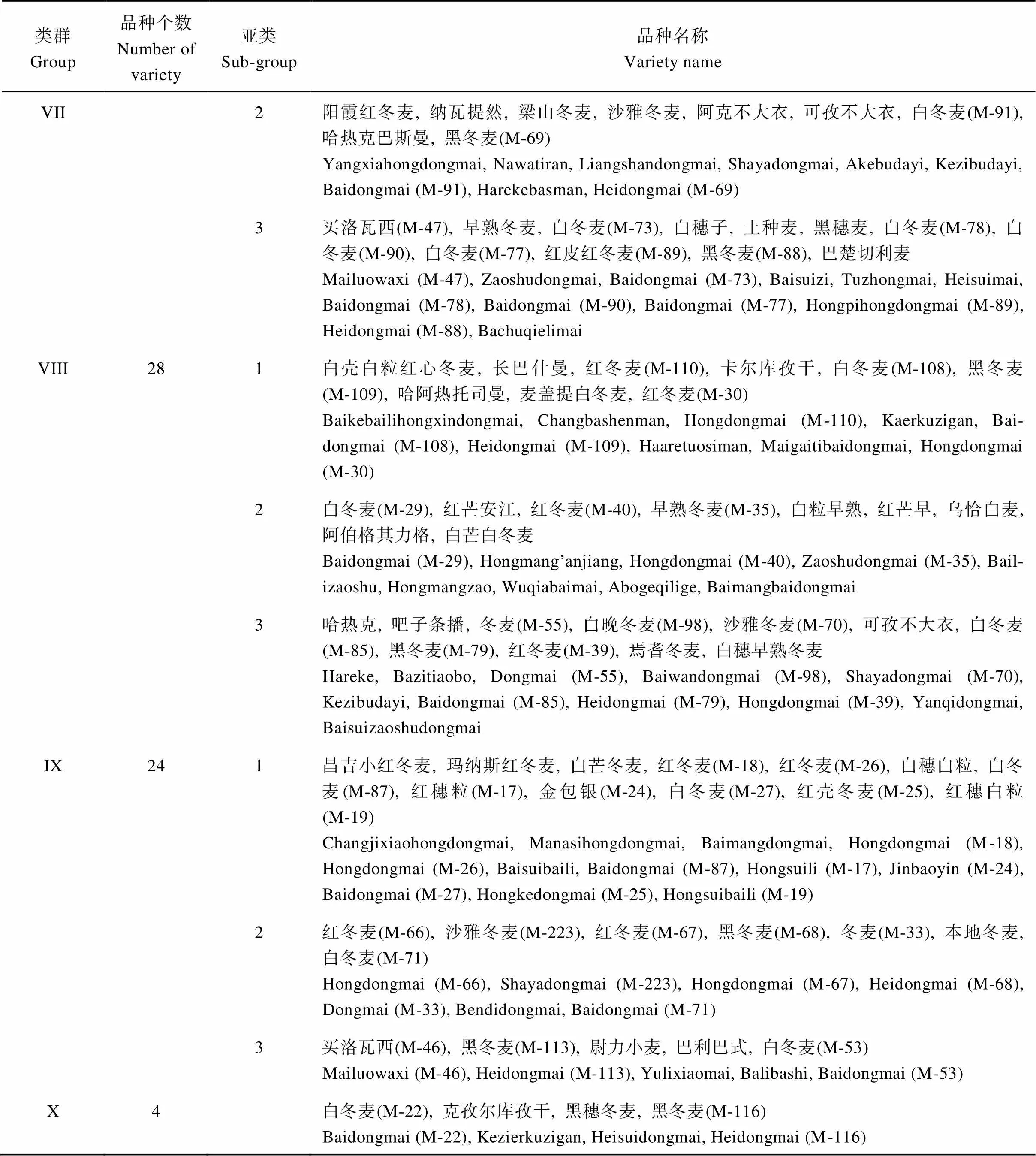
(续表3)
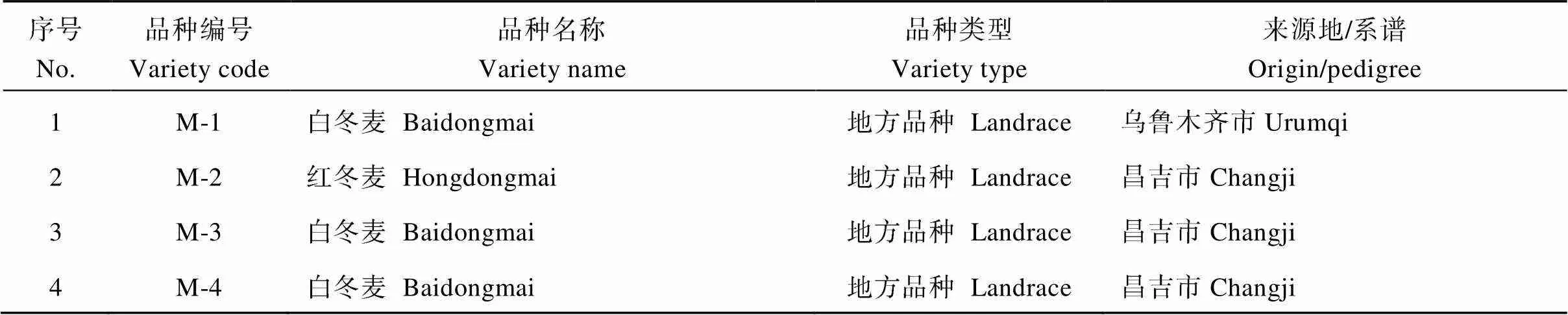
表4 SNP分析的新疆冬小麦品种名称及来源
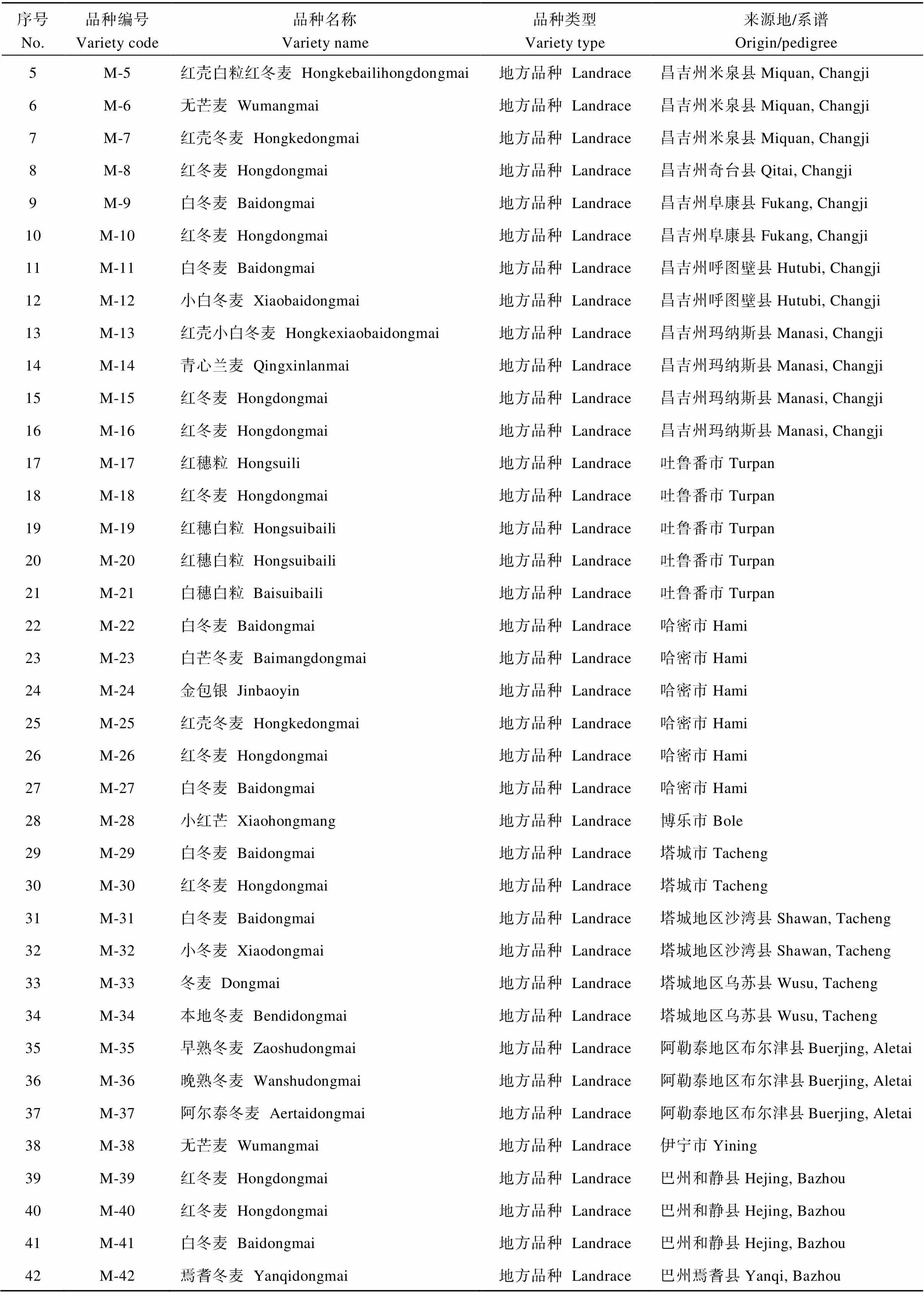
(续表4)

(续表4)
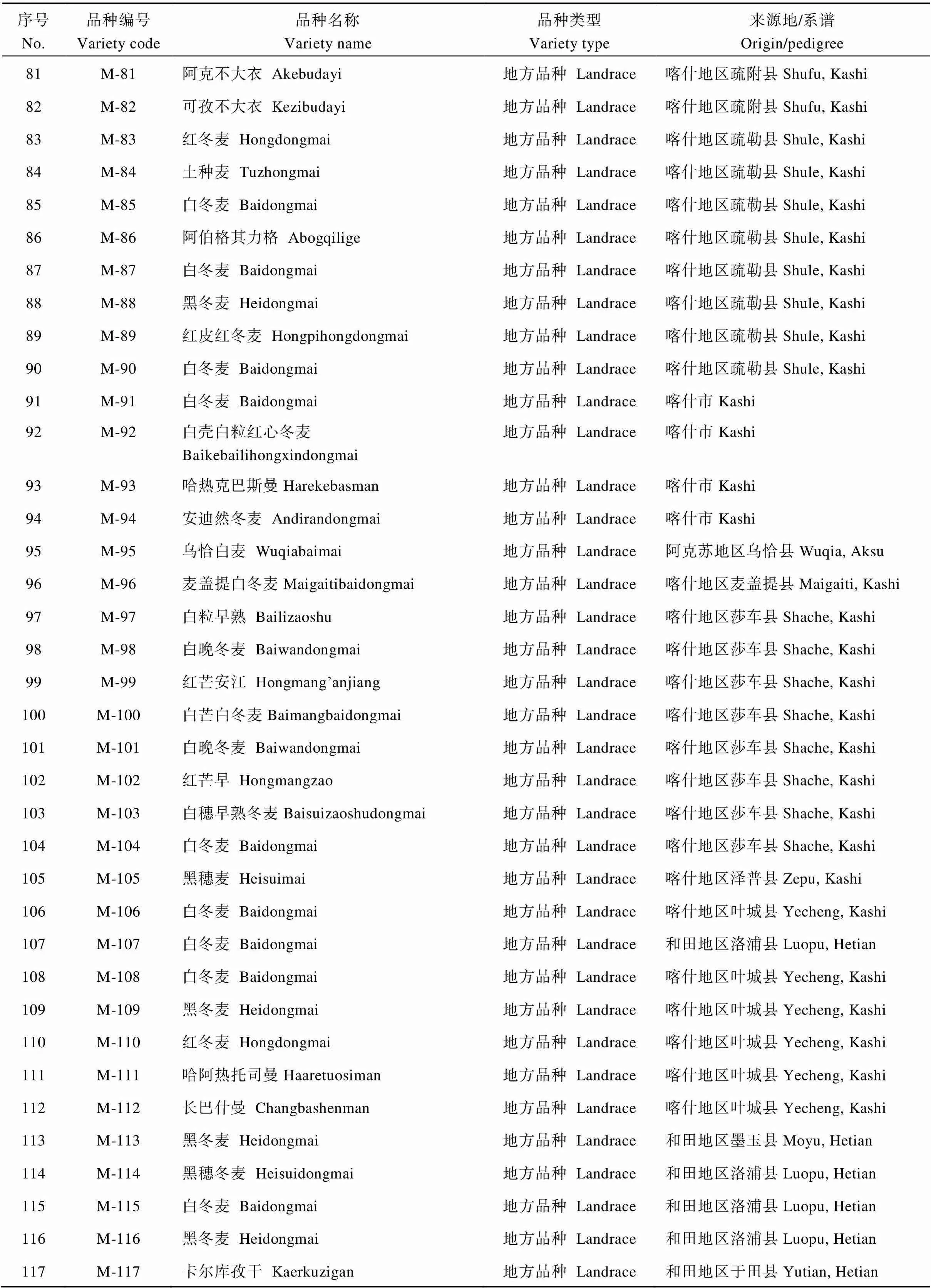
(续表4)
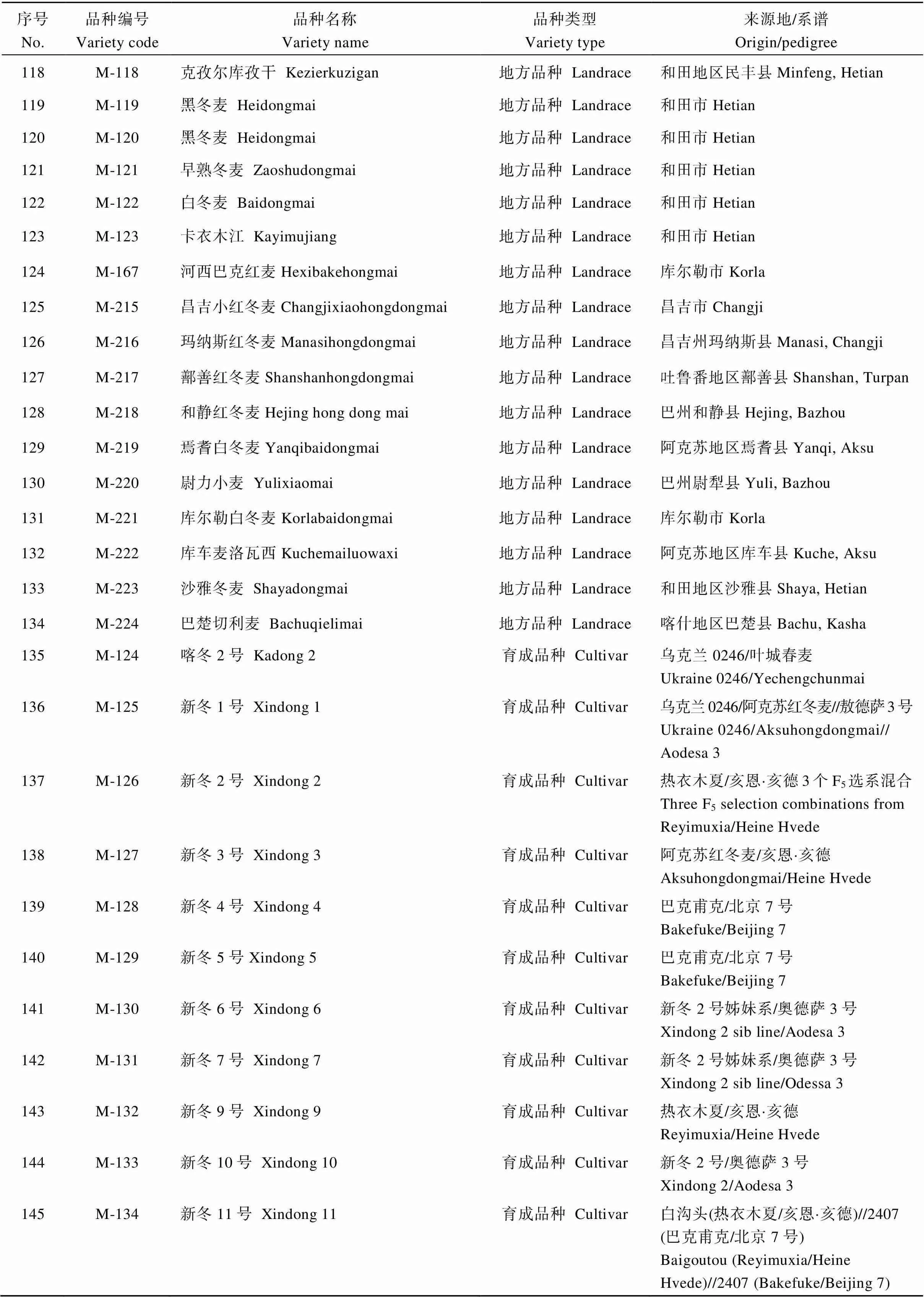
(续表4)
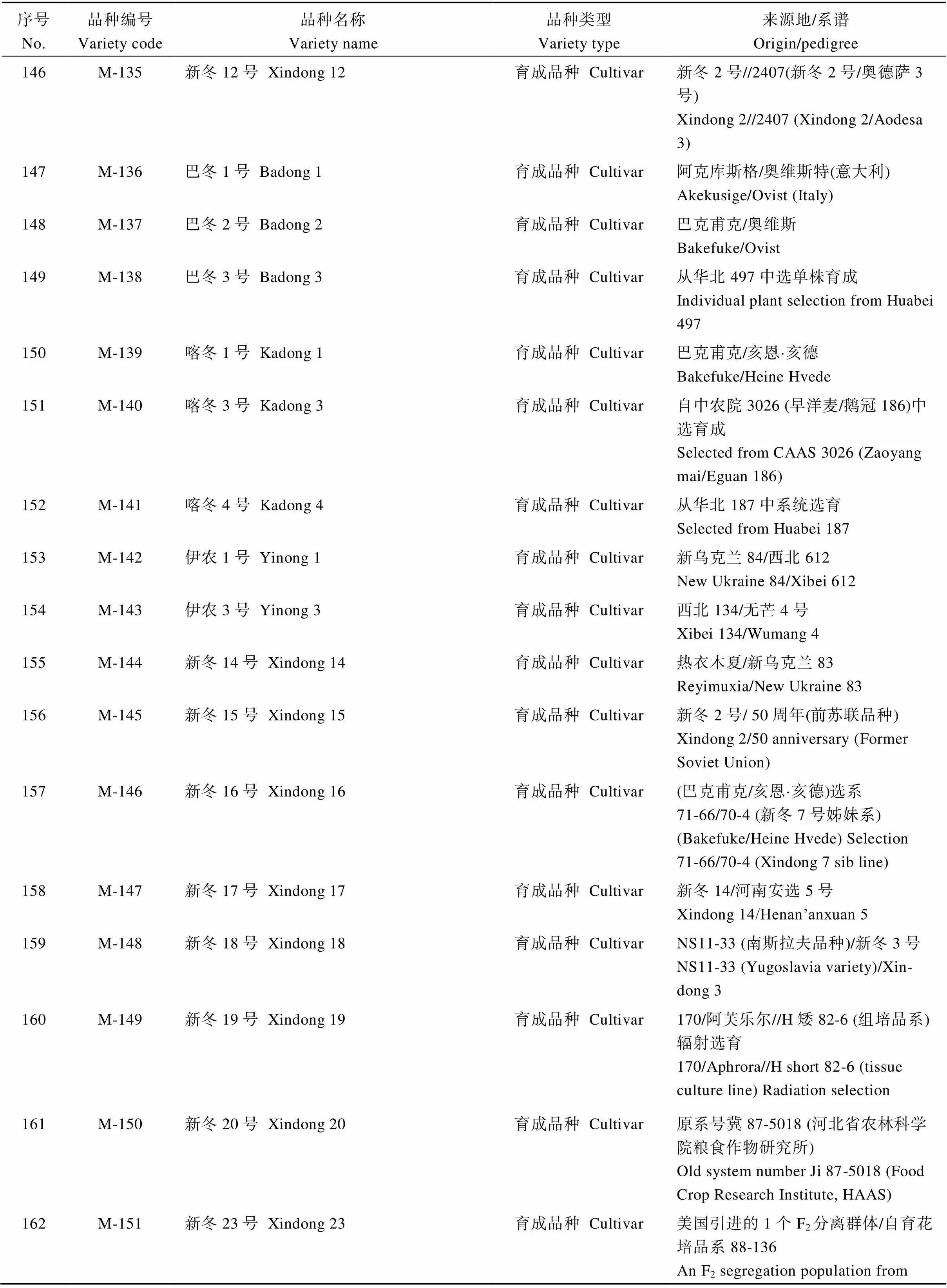
(续表4)
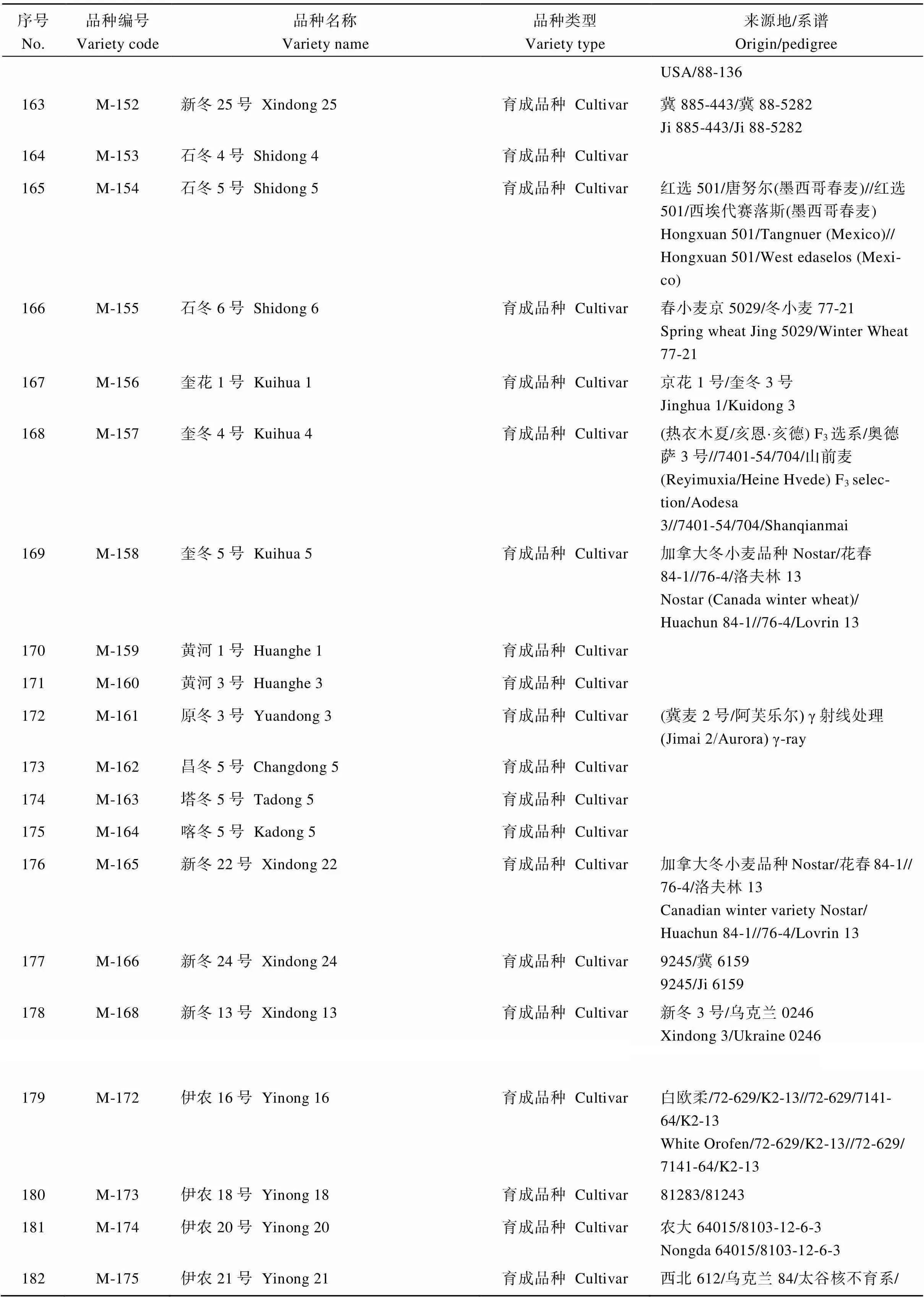
(续表4)
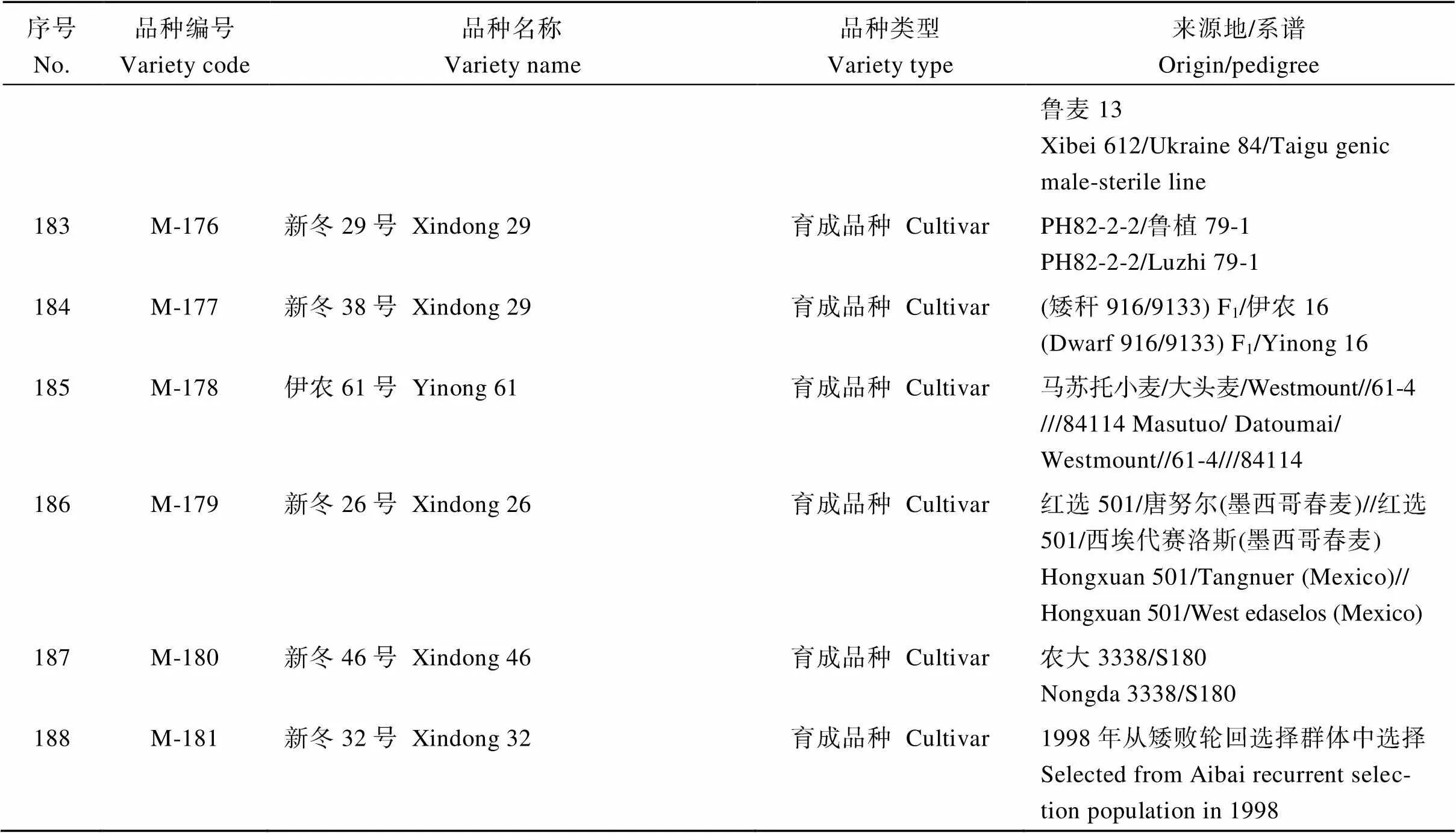
(续表4)
表中昌吉回族自治州简称昌吉州(Changji), 巴音郭楞蒙古自治州简称巴州(Bazhou), 伊犁哈萨克自治州简称伊犁州(Yili), 博尔塔拉蒙古自治州简称博州(Bozhou)。
The Changji Hui Autonomous Prefecture was abbreviated as Changji, the Bayingolin Mongolian Autonomous Prefecture was abbreviated as Bazhou, and the Bortala Mongol Autonomous Prefecture was abbreviated as Bozhou in the above table.
自第III类到第X类均为地方品种, 详述如下。
第III类有6个品种, 遗传距离在0.149~0.211之间。其中2份分别来自北疆昌吉州米泉县和伊犁州伊宁县的名称均无芒麦的品种聚在一起, 二者之间的遗传距离为0.166。其余4份材料均来自南疆地区, 2份来自喀什, 1份来自和田, 1份来自巴州, 地域相邻。
第IV类有5个品种, 遗传距离在0.093~0.274之间。2个分别来自喀什地区叶城县和和田地区洛浦县的均名为“白冬麦”的品种遗传距离最小, 为0.093; 来自阿克苏地区温宿县的“红冬麦”与来自喀什地区叶城县的白冬麦遗传距离最大, 为0.274。
第V类23个品种, 遗传距离在0.003~0.137之间, 包括红壳冬麦和白壳冬麦两种类型, 仅有2份白冬麦来自南疆喀什地区, 其余21份材料均来自北疆地区, 且有14份材料来自昌吉州, 表现为地域相近的材料聚在一起。其中来自乌鲁木齐市的“白冬麦”与阿勒泰地区布尔津县的晚熟冬麦之间的遗传距离最小, 为0.003; 来自昌吉州玛纳斯县的“红冬麦”与一份来自昌吉市的“白冬麦”之间的遗传距离最大, 为0.137。
第VI类18个品种, 遗传距离在0.003~0.111之间, 以白壳类型为主, 除了1个品种来自吐鲁番、2个品种来自和田地区外, 其余15个品种均来自南疆的阿克苏和巴州两个地区, 表现出明显的地域性。
第VII类25个品种, 遗传距离在0.003~0.125之间, 来自库尔勒市的买洛瓦西与来自和田市的早熟冬麦之间的遗传距离最小, 来自阿克苏地区新和县的梁山冬麦与喀什地区巴楚县的巴楚切利麦之间的遗传距离最大。此类包括白壳和红壳两种壳色类型, 分3个亚类, 第1亚类4个品种均来自巴州, 第2亚类9个品种中4个来自喀什地区、4个来自阿克苏地区、1个来自巴州; 第3亚类12个品种中有10份来自喀什地区, 也表现为地域相近的品种聚在一起。
第VIII类28个品种, 遗传距离在0.003~0.122之间, 包括红壳和白壳两种类型, 也分为3个亚类, 第1亚类9个品种中7个来自喀什地区; 第2亚类10个品种中有6个来自喀什地区, 其余4个品种分别来自塔城、阿勒泰、巴州和阿克苏地区; 第3亚类11个品种中6份来自喀什地区, 3份来自巴州, 2份来自阿克苏地区。
第IX类24个品种, 遗传距离在0.116~0.287之间, 来自昌吉州玛纳斯县的玛纳斯红冬麦与哈密地区哈密市的白芒冬麦之间的遗传距离最小, 来自喀什地区疏勒县的“白冬麦”与来自昌吉市的昌吉小红冬麦之间的遗传距离最大。分为3个亚类, 第1亚类12个品种中8个为红壳类型, 4个白壳类型, 其中8个品种来自吐鲁番, 3个品种来自与吐鲁番相邻的昌吉和哈密地区, 1个品种来自喀什地区。第2亚类7个品种中有2个来自北疆的塔城地区乌苏县, 5个均来自南疆的阿克苏、和田和巴州3个地区。第3亚类5个品种均来自南疆, 有4个品种来自巴州地区, 1个品种来自和田地区。
第X类最少, 仅有4个品种, 遗传距离在0.038~ 0.095之间, 3个来自和田地区, 1个来自哈密地区, 2个白壳类型聚在一起, 2个红壳类型聚在一起。
3 讨论
小麦地方品种是小麦育种的重要种质资源, 是改良品种的基础。地方品种是在自然经济农业时期, 在某地长期种植的“土著”品种, 由于各地生态条件千差万别, 造成地方品种具有不同的生态适应性[13-14]。新疆冬小麦地方品种多为强冬性, 具有耐寒、耐旱、耐盐碱、适应性强的特点, 但存在不耐肥、易倒伏、严重感锈病等缺陷, 逐渐被引进品种和自育品种所代替。育成品种(品系)往往都是丰产性、适应性、抗病性的结合体, 综合性状好, 是最便于利用的种质。在小麦育种中, 由于人们总是按照一定目标, 沿着一定方向选择, 使杂交育种亲本越来越集中到对当地条件最能适应, 综合性状最好, 配合力最佳的少数几个品种上, 导致育成品种遗传基础越来越狭窄[15]。如19世纪60年代, 新疆的小麦育种者利用具有抗寒、抗旱、耐盐碱特点的地方品种热衣木夏, 与引自丹麦的具有多花多实特性的冬小麦品种亥恩·亥德杂交育成新冬2号等品种及其衍生品种在南北疆大面积种植。本研究选用的54个新疆冬小麦育成品种中, 直接或间接以新冬2号为亲本选育成的就有新冬6号、新冬7号、新冬10号、新冬11号、新冬15号、奎冬4号6个品种, 这6个品种与其他42个育成品种聚在一类, 但新冬2号并没有与这6个品种聚在一类, 而是与新冬3号、新冬13号、巴冬1号等6个主要以新疆冬小麦地方品种与国外引进小麦品种为亲本育成的品种聚为一类(图3)。王立新等[16]对2005—2009年间547个国家冬小麦区域试验品系及134个国审品种分子标记遗传相似系数(GS)进行比较, 发现不同年度国家冬小麦区域试验GS>0.90的品种(系) 大多来自同一地区的育种单位。由此可见, 小麦育种中经常使用少数骨干亲本, 会导致小麦品种遗传多样性降低。李小军等[17]利用363对SSR标记对早洋麦、阿勃、欧柔和洛夫林10号等11份国外引种的骨干亲本材料及其衍生的33份代表性品种进行多样性分析, 发现衍生品种中约76.3%的等位变异类型均来自于国外亲本, 说明引种国外亲本对提高我国小麦品种的遗传多样性具有重要作用。
因小麦地方品种多为人们根据品种表型特征如颖壳色、种皮色、芒色+熟性/地名等方式命名, 且随人员流动而传播, 因此多存在同物异名或同名异物现象, 需要通过田间种植表型观察结合分子标记方法来判断区分。王琨等[18]对长期种植于陕西关中不同生态种植区的6个同名地方品种蚂蚱麦进行农艺性状遗传变异分析, 认为不同来源的地方品种蚂蚱麦在形态和农艺性状表现上的差异不大, 但差异程度与地理位置有关。李正玲等[19]利用SSR标记对河南省15组名称相同的小麦地方品种共计155份材料进行组间和组内遗传多样性分析发现, 同名品种组内15组材料的平均遗传相似系数变化范围为0.66~0.93; 同名品种组间遗传相似系数分布范围为0.75~1.00, 认为小麦地方品种具有丰富的遗传多样性, 同名小麦地方品种间存在同名同质和同名异质的现象。本研究所选134个地方品种中, 名为“白冬麦”的有26个, 名为“红冬麦”的有17个, 名为“黑冬麦”的有9个, 根据品种间遗传距离和聚类结果, 证明同名品种间有差异, 品种间遗传差异与地理位置有关, 这与王琨等[14]的研究结果一致。
目前国内多以SSR标记开展小麦地方品种遗传多样性研究[20-24], 针对小麦地方品种材料仅见利用660K SNP标记进行研究的报道[25-26]。王宏飞等[22]利用42对SSR引物对75份新疆小麦地方品种进行遗传多样性分析, 共检测到317个等位变异, 等位变异变化范围为2~15个, 平均7.55个, PIC值变化范围为0.169~0.905, 平均0.696; 基因组的平均等位变异是D>B>A, 春小麦地方品种的遗传多样性高于冬小麦地方品种, 聚类结果反映了一定的地域特性。Zhou等[25]为了揭示中国小麦的传播历史、适应性进化和品种选择, 利用27,933个DArTseq标记和312,831个660K SNP标记分析来自中国10个农业生态区的717个中国小麦地方品种的遗传结构, 证明小麦首先是从中国西北部引进种植的, 环境压力和独立选择导致了中国小麦地方品种在种群水平上的差异, 群体遗传结构在很大程度上与中国小麦地方品种的地理分布相一致, 大多数具有亲缘关系的小麦地方品种来源地相近, 与各小麦种植区的小麦栽培历史一致; 在A、B、D三个基因组中, D基因组的基因多样性少于A和B基因组; 在单条染色体上, 中国小麦地方品种端粒的遗传变异远高于近着丝粒。陕西关中蚂蚱麦和山西平遥小白麦是我国北方小麦品种的原始骨干亲本, 白彦明等[26]利用小麦660K SNP芯片对陕西关中蚂蚱麦和山西平遥小白麦及其衍生品种(系)共计149份材料进行全基因组扫描, 发现来自黄淮冬麦区的4份蚂蚱麦与来自北部冬麦区的小白麦及其系选品种燕大1817的平均遗传相似系数GS为0.750, 高于蚂蚱麦和小白麦衍生系的GS (平均为0.619), 与本研究中育成品种的遗传多样性高于地方品种的结论一致。针对小麦地方品种与育成品种之间的遗传差异, 魏育明等[24]利用24个SSR标记对8份四川小麦地方品种和8份主栽品种的遗传多样性比较研究, 表明四川小麦地方品种群体内的遗传多样性(平均GS为0.812)低于主栽小麦品种群体内的遗传多样性(平均GS为0.670), 也与本研究的结论相似。推测原因, 可能是在早期较低的生产水平和环境条件下, 通过长期的人工和自然选择造成的地方品种等位基因趋同化。因此, 在育种过程中应充分利用外来种质资源, 实现外来种质中优异基因资源的导入和聚合, 拓宽当地小麦品种的遗传基础。
SNP标记作为第三代分子标记, 因其具有自动化程度高、通量大、速度快、易于建立标准化操作等优势, 已成为研究种质资源遗传多样性、遗传结构评估以及推断个体间祖先关系的有力工具[27-28]。国内目前利用SNP标记基因分型开展遗传多样性分析多见于以小麦育成品种为材料的报道[29-31]。刘易科等[29]利用高密度SNP芯片, 分别对国内不同麦区的229份冬小麦品种(系)进行遗传多样性研究, 发现长江中下游麦区小麦品种(系)的相似性系数最高(0.712), 黄淮南片麦区、黄淮北片麦区和北部麦区的小麦品种(系)的相似性系数相对较低(0.660~0.686), 可能与不同区域小麦育种单位实力以及引进小麦种质资源的遗传多样性有关。李珊珊等[30]利用小麦90K基因芯片对143份河北省小麦育成品种的基因分型数据分析其亲缘关系与遗传多样性表明, 河北省143份小麦育成品种的遗传相似系数主要分布于0.52~0.73之间(占89.23%), 低于曹廷杰等[31-32]对河南省2000—2013年审定的96个小麦品种遗传多样性研究的结果(主要分布在0.63~0.83之间, 占95.94%)。从以上结果可以看出, 遗传相似性高的小麦品种具有相似的地域性或来源于同一育种单位, 表明近年来各育种单位育种材料趋同, 且相同或相似的骨干亲本高频被应用, 导致小麦品种遗传多样性降低。
国外也有利用SNP标记对小麦品种资源开展遗传多样性研究的报道。Zahra等[33]利用152K SNP芯片评估了188个来自西亚和北非的面包小麦品种的遗传多样性, UPGMA聚类分析将基因型明确分为两大类, 一类包含177个品种, 另一类仅11个品种, 种质之间的相似系数在0.30~0.99之间, 平均值为0.64。Bhatta等[34]利用47,526个SNP标记对包括人工合成小麦和面包小麦种质的143份材料进行遗传多样性分析表明, SNP标记均匀分布在A、B、D三个基因组中; 群体结构分析根据小麦种质的类型和地理起源分为3个不同的小麦基因型群(日本人工合成小麦组, CIMMYT人工合成小麦组和面包小麦组);遗传多样性比较表明, 人工合成小麦比面包小麦品系高33%, 认为人工合成小麦可用于拓宽现代小麦品种的遗传多样性。
依据本研究中55K芯片结果, 在分布于21条染色体上的53,063个SNP位点中有50,743个被检测到多态性, 多态性比率高达95.62%, 远高于刘易科等[29]、李珊珊等[30]、曹廷杰等[31-32]利用90K SNP获得的多态性; 并且本研究中SNP标记在A、B、D基因组间分布比较均匀, 分别为17,702个, 16,943个和16,098个, 而前人的90K芯片的SNP标记在B和D基因组间存在较明显的不均匀分布现象[29-32]。其原因可能由于小麦55K SNP芯片是以国内小麦材料为基础开发的, 并且所用的材料有普通小麦、四倍体小麦以及小麦野生近缘种材料, 因此SNP位点在各染色体上的分布更加均匀。由此可见, 小麦55K SNP芯片可能更加适宜于对国内小麦种质材料开展遗传多样性研究。
4 结论
利用我国自主研发, 覆盖小麦全基因组的55K SNP芯片, 对134个新疆冬小麦地方品种和54个新疆冬小麦育成品种进行全基因组扫描, 根据品种间的遗传距离将188个品种分为 10个类群, 育成品种集中聚在2个类群, 地方品种聚在另外8个类群。新疆冬小麦育成品种之间的遗传多样性略大于地方品种, 但育成品种与地方品种之间的遗传差异较大, 说明在现代小麦育种中, 地方品种可作为进一步拓宽育成品种遗传基础的育种材料。本研究证明小麦55K SNP标记能够揭示小麦品种资源的遗传差异以及品种间的亲缘关系, 可为小麦育种亲本选配提供重要理论依据。
[1] 赵吉平, 任杰成, 郭鹏燕, 许瑛, 任超. 小麦常规育种技术与改进策略研究. 中国种业, 2019, (7): 13–15.Zhao J P, Ren J C, Guo P Y, Xu Y, Ren C. Study on wheat general breeding techniques and improvement strategies., 2019, (7): 13–15 (in Chinese).
[2] 张冬梅, 辛涛, 米克拉依·吾甫尔. 新疆小麦生产现状及增产潜力分析. 中国农技推广, 2014, 30(11): 9–10. Zhang D M, Xin T, Mikrayi Wufuer. Analysis of Xinjiang wheat production status and yield increase potential., 2014, 30(11): 9–10 (in Chinese).
[3] 郎新婷, 马惠兰. 新疆小麦生产效率及地区差异研究. 中国农业资源与区划, 2016, 37(10): 127–133. Lang X T, Ma H L. Wheat production efficiency and differences in Xinjiang., 2016, 37(10): 127–133 (in Chinese).
[4] 林作楫, 揭声慧. 近50年河南省小麦育种工作的回顾. 河南农业科学, 2005, (9): 5–8. Lin Z J, Jie S H. Review of wheat breeding in Henan province in the past 50 years., 2005, (9): 5–8 (in Chinese).
[5] 李振声. 我国小麦育种的回顾与展望. 中国农业科技导报, 2010, 12(2): 1–4. Li Z S. Retrospect and prospect of wheat breeding in China., 2010, 12(2): 98–104 (in Chinese with English abstract).
[6] Lander E S. The new genomics: global views of biology., 1996, 274: 536–539.
[7] Liu J J, Luo W, Qin N N, Ding P Y, Zhang H, Yang C C, Mu Y, Tang H P, Liu Y X, Li W, Jiang Q T, Chen G Y, Wei Y M, Zheng Y L, Liu C J, Lan X J, Ma J. A 55K SNP array-based genetic map and its utilization in QTL mapping for productive tiller number in common wheat., 2018, 131: 2439–2450.
[8] Ren T, Hu Y, Tang Y, Li C, Yan B, Ren Z, Tan F, Tang Z, Fu S, Li Z. Utilization of a wheat 55K SNP array for mapping of major QTL for temporal expression of the tiller number., 2018, 9: 333.
[9] Ye X L, Li J, Cheng Y K, Yao F J, Long L, Yu C, Wang Y Q, Wu Y, Li J, Wang J R, Jiang Q T, Kang H Y, Li W, Qi P F, Lan X J, Ma J, Liu Y X, Jiang Y F, Wei Y M, Chen X M, Liu C J, Zheng Y L, Chen G Y. Genome-wide association study reveals new loci for yield-related traits in Sichuan wheat germplasm under stripe rust stress., 2019, 20: 640.
[10] Ye X L, Li J, Cheng Y K, Yao F J, Long L, Yu C, Wang Y Q, Wu Y, Li J, Wang J R, Jiang Q T, Li W, Ma J, Wei Y M, Zheng Y L, Chen G Y. Genome-wide association study of resistance to stripe rust (f. sp.) in Sichuan wheat., 2019, 19: 147.
[11] Saghai-Maroof M A, Soliman K M, Jorgensen R A, Allard R W. Ribosomal DNA spacer-length polymorphisms in barley: mendelian inheritance, chromosomal location, and population dynamics., 1984, 81: 8014–8018.
[12] Liu K J, Muse S V. Power Marker: an integrated analysis environment for genetic marker analysis., 2005, 21: 2128–2129.
[13] 庄巧生. 中国小麦品种改良及系谱分析. 中国农业出版社, 2003. pp 393–419. Zhuang Q S. Chinese Wheat Improvement and Pedigree Analysis. Beijing: China Agriculture Press, 2003. pp 393–419 (in Chinese).
[14] Dwivedi S L, Ceccarelli S, Blair M W, Upadhyaya H D, Are A K, Ortiz R. Landrace germplasm for improving yield and abiotic stress adaptation., 2016, 21: 31–42.
[15] 中国农学会遗传资源学会编. 中国作物遗传资源. 中国农业出版社, 1994. pp 3–7. Society of Crop Genetic Resources Chinese Association of Agricultural Science Societies. Crop Genetic Resources in China. Beijing: China Agriculture Press, 1994. pp 3–7 (in Chinese).
[16] 王立新, 李宏博, 廖琴, 邱军, 常利芳, 刘丽华, 任立平, 高新欢, 赵昌平. 利用分子标记筛查小麦相似品种(系). 作物学报, 2010, 36: 1490−1497. Wang L X, Li H B, Liao Q, Qiu J, Chang L F, Liu L H, Ren L P, Gao X H, Zhao C P. Screening wheat varieties with genetic similarity using molecular markers., 2010, 36: 1490−1497.
[17] 李小军, 徐鑫, 刘伟华, 李秀全, 杨欣明, 李立会. 应用SSR分子标记分析国外种质对我国小麦品种的遗传贡献. 作物学报, 2009, 35: 778−785. Li X J, Xu X, Liu W H, Li X Q, Yang X M, Li L H. Genetic contribution of introduced varieties to wheat breeding in China evaluated using SSR markers., 2009, 35: 778−785 (in Chinese with English abstract).
[18] 王琨, 崔志钢, 顾昌华, 赵会芳, 张玲丽. 同名小麦地方品种形态农艺性状遗传多样性分析. 江苏农业科学, 2014, 42(8): 77–80. Wang K, Cui Z G, Gu C H, Zhao H F, Zhang L L. Genetic diversity analysis of morphological and agronomic traits of wheat with the same name., 2014, 42(8): 77–80 (in Chinese with English abstract).
[19] 李正玲, 胡琳, 王会伟, 董海滨, 李艳, 李春鑫, 许为钢. 河南省同名小麦地方品种SSR遗传多样性分析. 麦类作物学报, 2016, 36: 564–570.Li Z L, Hu L, Wang H W, Dong H B, Li Y, Li C X, Xu W G. Genetic diversity of Henan wheat landraces with the same name detected by SSR markers., 2016, 36: 564–570 (in Chinese with English abstract).
[20] 吴涛. 长江中上游流域小麦地方品种基于SSR的遗传多样性分析. 华中农业大学硕士学位论文, 湖北武汉, 2013.Wu T. Analysis of Genetic Diversity of Wheat Landraces from Yangtze River Valley by SSR Markers. MS Thesis of Huazhong Agricultural University, Wuhan, Hubei, China, 2013 (in Chinese with English abstract).
[21] 张玲丽, 孙道杰, 冯毅, 王辉, 宋喜悦. 中国小麦地方品种的SSR遗传多样性分析. 西北农林科技大学学报(自然科学版), 2010, 38(7): 85–90. Zhang L L, Sun D J, Feng Y, Wang H, Song X Y. Analysis on genetic diversity of Chinese wheat landraces using SSR markers.(Nat Sci Edn), 2010, 38(7): 85–90 (in Chinese with English abstract).
[22] 王宏飞, 李宏琪, 丛花, 章艳凤, 肖菁, 宋羽, 严勇亮. 新疆小麦地方品种遗传多样性的SSR分析. 中国农业科技导报, 2010, 12(6): 98–104. Wang H F, Li H Q, Cong H, Zhang Y F, Xiao J, Song Y, Yan Y L. Genetic diversity of wheat landraces from Xinjiang using SSR markers., 2010, 12(6): 98–104 (in Chinese with English abstract).
[23] 卞春梅. 四川小麦地方品种重要SSR位点评价及其与主要性状相关分析. 四川农业大学硕士学位论文, 四川成都, 2009. Bian C M. Evaluation of Important SSR Locus of Sichuan Wheat Landraces and Relationship with Main Characters. MS Thesis of Sichuan Agricultural University, Chengdu, Sichuan, China, 2009 (in Chinese with English abstract).
[24] 魏育明, 郑有良, 周永红, 刘登才, 兰秀锦, 颜泽洪, 张志清. 四川小麦地方品种和主栽品种SSR多态性比较研究. 四川农业大学学报, 2001, 19(2): 117–121. Wei Y M, Zheng Y L, Zhou Y H, Liu D C, Lan X J, Yan Z H, Zhang Z Q. Comparison of genetic variations among Sichuan wheat landraces and cultivars based on SSR analysis., 2001, 19(2): 117–121 (in Chinese with English abstract).
[25] Zhou Y, Chen Z X, Cheng M Q, Chen J, Zhu T T, Wang R, Liu Y X, Qi P F, Chen G Y, Jiang Q T, Wei Y M, Luo M C, Nevo E, Allaby R G, Liu D C, Wang J R, Dvorák J, Zheng Y L. Uncovering the dispersion history, adaptive evolution and selection of wheat in China., 2018, 16: 280–291.
[26] 白彦明, 李龙, 王绘艳, 柳玉平, 王景一, 毛新国, 昌小平, 孙黛珍, 景蕊莲. 蚂蚱麦和小白麦衍生系的遗传多样性分析. 作物学报, 2019, 45: 1468–1477. Bai Y M, Li L, Wang H Y, Liu Y P, Wang J Y, Mao X G, Chang X P, Sun D Z, Jing R L. Genetic diversity assessment in derivative offspring of Mazhamai and Xiao-baimai wheat., 2019, 45: 1468–1477 (in Chinese with English abstract).
[27] Wang S, Wong D, Forrest K, Allen A, Chao S M, Huang B E, Salvi S, Milner S G, Cattivelli L, Mastrangelo A M, Whan A, Stephen S, Barker G, Wieseke R, Plieske J. Characterization of polyploid wheat genomic diversity using a high density 90,000 single nucleotide polymorphism array., 2014, 12: 787–796.
[28] Akhunov E D, Akhunova A R, Anderson O D, Anderson J A, Blake N, Clegg M T, Coleman-Derr D, Conley E J, Crossman C C, Deal K R. Nucleotide diversity maps reveal variation in diversity among wheat genomes and chromosomes., 2010, 11: 702.
[29] 刘易科, 朱展望, 陈泠, 邹娟, 佟汉文, 朱光, 何伟杰, 张宇庆, 高春保. 基于SNP标记揭示我国小麦品种(系)的遗传多样性. 作物学报, 2020, 46: 307–314 . Liu Y K, Zhu Z W, Chen L, Zou J, Tong H W, Zhu G, He W J, Zhang Y Q, Gao C B.Revealing the genetic diversity of wheat varieties (lines) in China based on SNP markers., 2020, 46: 307–314 (in Chinese with English abstract).
[30] 李珊珊, 易腾飞, 徐渴, 张树华, 赵勇, 杨学举. 河北省小麦品种基于SNP标记的遗传多样性分析. 分子植物育种, 2019, 17: 6850–6859. Li S S, Yi T F, Xu K, Zhang S H, Zhao Y, Yang X J.Genetic diversity of wheat varieties in Hebei province based on single nucleotide polymorphism., 2019, 17: 6850–6859 (in Chinese with English abstract).
[31] 曹廷杰, 谢菁忠, 吴秋红, 陈永兴, 王振忠, 赵虹, 王西成, 詹克慧, 徐如强, 王际睿, 罗明成, 刘志勇. 河南省近年审定小麦品种基于系谱和SNP标记的遗传多样性分析. 作物学报, 2015, 41: 197–206.Cao T J , Xie J Z, Wu Q H, Chen Y X, Wang Z Z, Zhao H, Wang X C, Zhan K H, Xu R Q, Wang J R, Luo M C, Liu Z Y. Genetic diversity of registered wheat varieties in Henan province based on pedigree and single-nucleotide polymorphism., 2015, 41: 197–206 (in Chinese with English abstract).
[32] 曹廷杰. 河南省小麦新品种(系)遗传解析. 北京: 中国农业大学, 2015.Cao T J. Genetic Analysis of New Wheat Varieties (lines) in Henan Province. Beijing: China Agricultural University, 2015 (in Chinese with English abstract).
[33] Zahra M T, Zahra T M, Kumarse N, Khalil Z N, Farshad R, Abdelfattah A D. Combined study on genetic diversity of wheat genotypes using SNP marker and phenotypic reaction to., 2019, 66: 1791–1811.
[34] Bhatta M, Shamanin V, Shepelev S, Baenziger P S, Pozherukova V, Pototskaya I, Morgounov A. Genetic diversity and population structure analysis of synthetic and bread wheat accessions in western Siberia., 2019, 60: 283–289.
Genetic diversity assessment of winter wheat landraces and cultivars in Xinjiang via SNP array analysis
MA Yan-Ming1,2, LOU Hong-Yao1, CHEN Zhao-Yan1, XIAO Jing2, XU Lin2, NI Zhong-Fu1, and LIU Jie1,*
1College of Agronomy and Biotechnology, China Agricultural University, Beijing 100193, China;2Institute of Crop Germplasm Resource, Xinjiang Academy of Agricultural Sciences, Urumqi, Xinjiang 830091, China
Xinjiang is a major wheat production and consumption area in northwest China. Assessment of genetic diversities of wheat landraces and cultivars in Xinjiang has great benefit for wheat cultivar breeding and improvement in this area. In this study, a wheat 55K SNP (single nucleotide polymorphism) chip was employed to perform genome-wide scanning for the genetic diversities of 188 winter wheat varieties from Xinjiang, including 134 landraces and 54 modern cultivars. Our results showed that the polymorphism ratio of the SNP loci was about 95.62% (50,743/53,063). Each chromosome contained 1068–2616 polymorphic loci, and the polymorphic SNP marker number in three genomes was A > B > D. The genetic distance among the 188 wheat varieties ranged from 0.002 to 0.723, with an average of 0.378. The genetic distance of 134 landraces was between 0.002 and 0.400, with an average of 0.070, whereas that of 54 modern cultivars was between 0.004 and 0.337, with an average of 0.114. The genetic distance between landraces and modern cultivars was between 0.605 and 0.723, with an average of 0.699. The clustering results showed that all tested wheat materials could be divided into ten different groups. According to the comprehensive analyses of SNP and pedigree, the genetic diversity was the largest between the cultivars and the landraces, medium between different landraces, and small between modern cultivars. Because the genetic diversity of Xinjiang winter wheat cultivars is not rich enough, the wheat landraces should be used as the breeding materials in the future. Our data provide novel scientific basis and guidance for breeding and improvement of wheat cultivars in Xinjiang.
Xinjiang; winter wheat; landraces; modern cultivars; SNP; genetic diversity
10.3724/SP.J.1006.2020.91077
本研究由国家农作物种质资源共享服务平台(新疆)项目(NICGR2015-029)和科技部科技基础性工作专项“西北干旱区抗逆农作物种质资源调查”项目(2011FY110200)资助。
This study was supported by the National Ministry of Science and Technology and the Ministry of Finance's National Crop Germplasm Resources Sharing Service Platform (Xinjiang) Project (NICGR2015-029) and the Special Items for Basic Science and Technology Work of Science and Technology Ministry, “Investigation on Germplasm Resources of Stress-resistant Crops in Arid Area of Northwest China” (2011FY110200).
刘杰, E-mail: liujie13251@163.com, Tel: 010-62734072
E-mail: ymma213@sina.com
2019-12-27;
2020-03-24;
2020-04-28.
URL: http://kns.cnki.net/kcms/detail/11.1809.S.20200428.1250.002.html
猜你喜欢
区域治理(2022年40期)2022-11-27
今日农业(2022年4期)2022-06-01
农业灾害研究(2022年1期)2022-05-07
少儿科学周刊·儿童版(2021年21期)2021-12-11
今日农业(2021年4期)2021-06-09
动漫界·幼教365(小班)(2019年10期)2019-10-28
动漫界·幼教365(大班)(2019年10期)2019-10-28
动漫界·幼教365(中班)(2019年10期)2019-10-28
小学生导刊(2018年34期)2018-12-18
山东青年(2016年3期)2016-02-28
