
转基因大豆及其安全性评价研究进展
2020-09-09 01:18马启彬卢翔杨策王立平
安徽农业科学 2020年16期
马启彬 卢翔 杨策 王立平
摘要 转基因大豆是商业化规模最大的转基因作物,创造了巨大的经济效益。我国是转基因大豆进口量和压榨量最大的国家,民众对转基因技术的风险性及其产品的安全性存在质疑。对控制大豆重要性状的基因挖掘、功能研究及其商业化开发、转基因大豆的食用安全性和生态安全性等研究进展及存在的主要问题等进行分析,并从转基因大豆的法律法规建设、田间试验和生态安全评价、食用安全评价和科普宣传等方面提出政策建议。
关键词 转基因大豆;豆制品;食用安全性;安全评价;科普宣传
中图分类号 TS 201.6文献标识码 A
文章编号 0517-6611(2020)16-0020-05
doi:10.3969/j.issn.0517-6611.2020.16.003
Research Progress on Genetically Modified Soybean and Its Safety Evaluation
MA Qi-bin,LU Xiang,YANG Ce et al (College of Agriculture,South China Agricultural University,Guangzhou,Guangdong 510642)
Abstract Genetically modified soybean (GMS) creating huge economic benefits is the most commercialized genetically modified crop.China is the country with the largest import and crush of GMS,while Chinese people have doubts on the risk of genetically modified technology and the safety of its products.The research progress of gene mining,functional research,commercial development,edible safety and ecological safety of GMS,and existing problems were analyzed.Some policy suggestions were further put forward in the aspects of laws and regulations construction,field test and environmental safety evaluation,food safety evaluation and popular science publicity of GMS.
Key words Genetically modified soybean;Bean products;Edible safety;Safety evaluation;Popular science propaganda
基金項目
农业农村部农产品质量安全监管专项(21301091702101,4100-C17106);转基因生物新品种培育科技重大专项(2016ZX08004002-007)。
作者简介 马启彬(1968—),男,安徽六安人,副教授,博士,硕士生导师,从事大豆抗逆分子育种研究。*通信作者,研究员,博士,从事农产品质量安全与营养评价研究。
收稿日期 2020-02-24;修回日期 2020-03-19
转基因技术作为生命科学的核心技术之一,广泛用于作物的性状改良。转基因作物(genetically modified crop,GMC)是指通过基因工程手段将来自不同生物的基因整合到目标作物的基因组,使其具有更好的性状或品质的作物[1]。随着转基因技术研究和应用的深入,转基因作物的种植面积迅速扩大,转基因农产品和转基因食品在粮食市场所占的份额也越来越大。因此,转基因作物及其农产品、食品的安全性等受到全球关注[2-3]。目前,主要从短期效应和长期效应2个方面对转基因农产品的食用安全性进行评价,其评价指标主要包括营养成分和抗营养因子、毒性及致敏性、标记基因的安全性及非预期效应等[2]。据国际农业生物技术应用服务组织(ISAAA,http://www.isaaa.org/)统计,2018年全球转基因农作物种植面积为1.917亿hm2,在转基因作物商业化的23年(1996—2018年)中,累计达25亿hm2,有效地保障粮食供给,降低了资金的投入[4-5]。
自转基因安全管理条例颁布实施以来,我国正式批准商业化生产的作物主要有棉花、西红柿、烟草、牵牛花和番木瓜等,其中真正大面积生产的转基因作物只有棉花。然而,随着国民生活质量的提高,我国进口转基因大豆的总量不断攀升,现已成为世界上最大的转基因大豆进口国,近3年转基因大豆的进口量每年9 000万t左右,致使我国大豆的自给率只有15%左右(中国海关总署)。因此,转基因技术和转基因大豆食品的安全性备受各方广泛关注。笔者对控制大豆重要性状的基因功能、转基因大豆的商业化研发、转基因大豆的食用安全性和生态安全性评价等方面进行分析,阐述转基因大豆及其安全性研究的现状,在此基础上指出了我国转基因大豆产业发展存在的问题,提出了合理的发展建议,以期促进大豆产业的良性发展。
1 转基因大豆研究概况
1.1 转基因大豆商业化进展
自1996年转基因作物商业化种植以来,转基因大豆的种植面积持续增加[6]。转基因大豆种植面积从1996年50万hm2增至2018年9 590万hm2,占全球转基因作物种植面积的50%[4]。转基因大豆品种开发在抗除草剂、抗虫和品质改良方面取得了突破性研究进展。
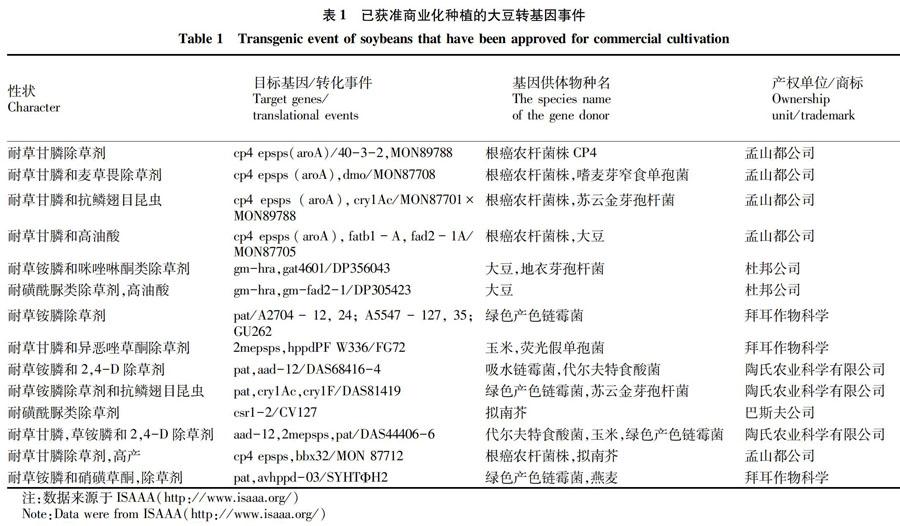
抗除草剂转基因大豆是商业化最早、类型最多的转基因大豆(表1)。据国际农业生物技术应用服务组织统计,已有32个抗除草剂转化体获得批准商业化种植,主要包括耐草甘膦、耐草铵膦、抗麦草畏和抗2,4-D等类型[4]。其中耐草甘膦大豆品种GTS 40-3-2最先商业化种植,该品种为美国孟山都公司(Monsanto Company)研发,转入的CP4-EPSPS基因来自矮牵牛。目前,耐草甘膦大豆品种GTS40-3-2共获得27个国家/地区和欧盟28国的55个批文,是目前应用最为广泛的转基因大豆。拜耳作物科学公司主要针对草铵膦研发抗除草剂大豆品种,其中A2704-12、A2704-21、A5547-35品种在1996年已获批准进行商业化种植[7]。为了加强大豆种子对环境的适应能力,孟山都公司开发了既抗草甘膦又抗麦草畏的MON8708×MON89788转基因大豆品种;美国陶氏农业有限公司则将pat、CP4-EPSPS和AAD-12 3个基因聚合在一起,研发出耐草铵膦、草甘膦基和2,4-D 3种除草剂的DAS68416×MON89788转基因品种(ISAAA)。
继抗除草剂轉基因大豆之后,抗虫转基因大豆品种的研发也取得了较快的进展(表1)。目前已经获得商业化批准的抗虫转基因大豆共有6个转化体,其中孟山都公司有4个,美国陶氏农业有2个。孟山都公司将苏云金芽孢杆菌中的抗虫基因Cyr1Ac转入大豆,培育的抗虫大豆品种MON87701和MON87751能有效预防鳞翅目昆虫[8-9];随后,孟山都公司培育的MON87701×MON89788一系列产品,同时含有抗草甘膦基因CP4-EPSPS和抗虫基因Cyr1Ac,能够有效防治杂草和鳞翅目昆虫,实现了将抗虫、耐除草剂性状聚合在一个大豆品种,加强了转基因大豆的竞争力(ISAAA)[10]。
在大豆品质改良方面,早在1997年美国杜邦公司培育了G94-1、G94-19、G168高油酸转基因大豆品种并商业化种植[11]。杜邦公司开发了无反式脂肪且油酸含量高达75%的Plenish转基因大豆品种,2012年在美国正式批准上市销售,于2014年获得中国进口许可(ISAAA)[11-12]。
1.2 控制大豆重要性状基因研究进展
随着基因组测序及转基因技术的发展,控制大豆重要性状基因功能研究越来越多,为大豆品种改良积累了丰富的基因资源。这些基因的功能涉及大豆的产量、品质、抗病虫性、抗非生物胁迫和营养等性状。研究表明,GmPPR4、GmWRKY54、GmSYP24、GmWRKY12、AtABF3、AtDREB1和FvC5SD等基因的过表达增强转基因大豆的抗旱性,其中GmSYP24和GmWRKY12基因的过表达也使得转基因大豆具有耐盐性[13-19]。GmMYB3a基因通过下调一些与植物防御信号通路相关的关键基因负向调控植物盐胁迫响应[20]。
研究表明,一些基因对产量性状具有调控作用。过量表达GmMYB68基因可增加大豆粒数和百粒重,同时也增强了转基因大豆对盐碱的抗性[21]。过量表达苜蓿MsWRKY11基因不仅增加了转基因大豆苗期的耐盐性,而且转基因MsWRKY11大豆株高、单株荚果数、单株种子数、百粒重等性状均高于野生型[22]。在抗病虫性方面,过量表达GmSnRK1.1、GmDIR22、GmWRKY31、GmHDL56、GmERF113、GmIFR、GmERF5、hrf2和PAC1等基因可增强转基因大豆对疫霉病的抗性[23-30]。GmSAMT1、SpbP0基因等过表达则可提高转基因大豆的抗虫性[31-33]。此外,GmWRI1a转录因子可正向调控大豆种子的油分积累[34]。
2 转基因大豆食用安全性评价
随着抗除草剂或/和抗虫转基因大豆种植面积的不断扩大,转基因大豆的加工产品直接或者间接被人类食用。转基因食品改变了传统食品的自然属性,食用安全具有未知性,因此,其食用风险备受关注[35]。根据国际食品法典委员会的标准,转基因大豆的食用安全性采用实质性等同的法则进行评价,主要包括营养学、毒理学和过敏性3个方面[36]。
2.1 抗除草剂转基因大豆营养学评价
转基因大豆营养学的评价多采用饲喂动物进行评估,研究表明,用抗草甘膦转基因大豆饲喂小鼠、大鼠、蛋鸡、肉鸡、鲶鱼、兔子、鲑鱼、奶牛和猪等,其生长和产品特性未产生显著影响[37-45]。与此相对应,研究表明,抗除草剂转基因大豆中具有抗癌作用的异黄酮成分会减13%左右[46],而抗草甘磷转基因大豆所含酚类物质的种类与含量也与非转基因大豆存在差异[47],转基因大豆油的营养及健康效应均不如非转基因大豆油[48]等。
2.2 抗除草剂转基因大豆毒理学评价
转基因大豆的毒理学评价主要是依靠饲喂动物待测的转基因大豆,然后观察动物是否出现中毒症状,检测动物的生理生化指标来确定待测大豆的毒性和安全摄入量,再通过所得到的数据推论到人。朱元招[49]研究表明,用RR抗草甘膦转基因豆粕高水平饲喂大鼠,其生长、生理机能及组织器官发育无明显病变,肌肉组织中也未检测出明显的外源DNA残留。Appenzeller等[50]用抗草甘膦大豆356/43饲喂大鼠,检测大鼠神经系统的功能,结果表明转基因大豆对大鼠神经行为学无显著影响。芦春斌等[51]以小鼠为实验对象进行免疫系统的检测,发现其亲代和子一代的脾淋巴细胞增殖无明显异常。但也有研究指出,抗草甘膦大豆引起成年小鼠的胚胎细胞、胰腺、睾丸的形态结构和相应酶的活性发生变化,扰乱了其正常的器官功能。Malatesta等[52]研究表明用抗草甘膦转基因大豆饲喂大鼠2年,老鼠体内衰老标记物的表达明显增多;同时兔子心脏和肾脏酶活性也发生了改变。
2.3 抗除草剂转基因大豆过敏性评价
过敏源的本质是蛋白质,而大豆中含有丰富的蛋白质,且外源基因也会表达出特异的蛋白质,这些都有可能成为过敏源。第一代抗除草剂转基因大豆RR中CP4-EPSPS氨基酸序列与1 935种已知的毒蛋白无氨基酸序列同源性,与已知的过敏蛋白特性如分子量大小、浓度、稳定性、糖基化等也无明显的同源性。因此,认为CP4-EPSPS没有明显的致敏性。食物蛋白导致过敏的另一个重要因素是这种蛋白在总食物蛋白中的比例,研究表明CP4-EPSPS蛋白只占大豆总蛋白的0.08%(大多数过敏原占总蛋白的1%~8%);通过榨油等加工手段可提取出CP4-EPSPS蛋白的绝大多数,认为精炼大豆油不会引起人类过敏反应。但也有报道指出,儿童饮用转基因大豆豆浆会产生过敏反应。因此,对于转基因大豆的致敏性尚待深入研究,暂不能定论[53]。
3 转基因大豆生态安全性评价
转基因作物因人為作用导致基因发生了改变,有可能破坏自然生存法则、改变原有的生态平衡,甚至影响生物的多样化。转化进入作物的基因有可能通过生物链等方式进行基因漂移、造成基因污染、破坏生态中生物的多样性平衡。抗除草剂转基因大豆是世界生产上种植的转基因大豆的主体,其长期种植可能导致生态问题,主要包括超级杂草的产生,对本土大豆种质资源的影响和对土壤微生物的影响等。
3.1 抗除草剂转基因大豆可能导致超级杂草的产生
长期种植抗除草剂大豆会导致选择压力加大,抗除草剂转基因大豆可能通过花粉向近缘野生植物转移,使这些植物含有抗除草剂基因或者产生突变并对除草剂产生抗性,从而成为“超级杂草”。Berberich等[54]研究认为,抗草甘膦大豆种植后已形成一种优势大豆品种,限制了普通栽培大豆的发展,且抗除草剂转基因大豆落粒成为轮作后茬作物田中杂草。随后的中长期种植转基因作物的研究表明,并没有发现转基因大豆、油菜、玉米等转变为杂草的可能性[55]。刘刚[56]以抗草甘膦转基因大豆SHZD32-01为试验材料,以受体中豆32和当地栽培种皖豆21为对照,观测栽培大豆品种与抗除草剂大豆品种在田间生长的竞争性,结果表明抗草甘膦转基因大豆SHZD32-01对草甘膦抗性较好,无栽培地生存竞争优势,不具有杂草化潜力。
3.2 抗除草剂转基因大豆可能影响大豆种质资源
大豆起源于我国,我国野生大豆有6 000多份,占全球野生大豆的50%以上。若外源目标基因漂移至野生大豆中,野生大豆原始性状可能会受到破坏,导致野生大豆等位基因丢失。转基因大豆的抗除草剂特性也可能使其变为杂草,其滋生蔓延将给大豆生产造成损失且会使大豆的遗传多样性丧失[55]。Abe等[57]研究发现,日本及俄罗斯的一些大豆栽培种来源于野生大豆和栽培大豆的自发杂交,分子证据也显示栽培大豆和野生大豆之间会发生基因渐渗的现象。陈新等[58]在田间种植抗草甘膦大豆,对其基因漂移的可能性进行了观测,并在南京试验地点检测到发生基因漂移的野生大豆1株。结果表明,抗除草剂转基因大豆向野生大豆发生基因漂移是可能的。
刘琦等[59]在田间生殖隔离状态下,将野生大豆和栽培大豆种植在抗草甘膦转基因大豆周围,按固定距离收获。每年连续筛查存活植株提取DNA,对目的基因CP4-EPSPS进行PCR检测,在监测3年内未发现抗草甘膦转基因大豆通过风媒介使花粉传播和基因漂移现象。吕晓波等[60]研究表明,抗草甘膦基因大豆的花粉在自然条件下发生漂移几乎是不可能的,但如果人为加大虫媒传粉,抗草甘膦基因的漂移概率接近0.05%,漂移距离为0.7 m。大豆的萌发活力在土壤中不能长时间保持,大豆落粒后第二年的自然萌发和生长现象较为罕见,杂草化风险较低。
3.3 抗除草剂大豆对土壤微生物的影响
土壤在农业生态系统中处于核心地位,是农业生产中物质转化与能量交换的重要场所。土壤微生物是土壤生态系统的重要组成部分,因此,评价转基因作物对土壤微生物的影响具有重要意义[61]。Means等[62]研究发现,在抗草甘膦转基因大豆田中大量喷施草甘膦会导致土壤根际微生物种群发生变化,并促进土壤中镰孢菌和腐霉菌的生长,从而延缓和减少了抗草甘膦大豆苗期根部的生长。杜伟等[63]研究发现,抗草甘膦转基因大豆AG5601影响根际土壤微生物的数量,但变化无规律可循。王争等[64]以抗草甘膦转基因大豆RRS为材料研究其根系土壤氮素转化,结果表明连续3年种植RRS对根际土壤细菌的生长和繁殖均有显著抑制作用。
朱银玲[65]以EPSPS转基因抗除草剂大豆为材料,测定不同大豆样品根际土的理化性质,大豆植株、种植及根际土的氮含量,根际土的碳、氮循环相关酶活,根际土可培养固氮菌数量,土壤固氮菌群的结瘤效应以及nifH基因丰度表征的土壤固氮菌群总体丰度变化等指标。结果表明,种植EPSPS转基因抗除草剂大豆或者喷洒一定量的草甘膦可能会对土壤微生物群落尤其是对根际土壤固氮菌群产生一定的影响。章秋艳等[66]盆栽种植抗草甘膦转基因大豆M88、GTS40-3-2、ZB及非转基因大豆中黄13,采用DGGE-cloning测序技术分析了成熟期大豆根际土壤固氮细菌的多样性。结果表明,抗草甘膦转基因大豆M88、GTS40-3-2和非转基因大豆中黄13的根际土壤固氮细菌多样性指数和均匀度指数无显著差异。与非转基因大豆相比,虽然抗草甘膦CP4-EPSPS基因导入大豆并未对根际土壤固氮细菌多样性产生显著影响,但这并不能认定转基因大豆对土壤生态环境没有影响,需要进行更深层次的长期研究[67]。
4 促进我国转基因大豆产业发展的策略及建议
4.1 健全转基因相关法律法规
自1996年起,我国开始进口转基因大豆,此后进口量迅速飙升,近3年转基因大豆进口量每年9 000万t左右,进口量占消费总量的85%左右。依据世界科技发展趋势,着眼于我国国情和农业发展的需要,农业农村部自2008年启动转基因生物新品种培育重大专项,组织国内高校和科研单位开展转基因生物新品种培育研究。为规范转基因新品种培育研发过程中的试验研究、转基因生物安全评价、转基因农产品的加工和食用安全评价等环节的规范实施,农业农村部先后出台并修订转基因相关法律法规,主要包括《农业转基因生物安全管理条例》及配套规章、《农业转基因生物安全评价管理办法》《转基因作物田间试验安全检查指南》《农业转基因生物进口安全管理办法》《农业转基因生物标识管理办法》《农业转基因生物加工审批办法》《进出境转基因产品检验检疫管理办法》(中华人民共和国农业农村部转基因权威关注网站,http://www.moa.gov.cn/ztzl/zjyqwgz/)等。
虽然这些法律法规对我国转基因生物新品种培育及其成果转化、转基因食品开发等发挥着重要作用,但在执行过程中依然存在很多问题,集中表现在安全监管执行难,民众对转基因技术的接受程度极低等。转基因安全监管采用“属地管理”,使得中央与地方权责划分出现脱节、地方监管在执行过程中出现实质性缺位现象;另一方面,公众缺乏转基因安全性基本知识,对某些媒体、公众人物及组织对转基因安全的“妖魔化”宣传缺乏监管,加剧了公众对转基因技术和食品安全的担忧[2]。因此,需要通过立法加强转基因研究安全性的执行力,对转基因安全监管加强规范管理,对转基因技术及食用安全等的妖言惑众者及组织予以批评教育或绳之以法。
4.2建立完备的大豆田间安全评价体系
2008年轉基因生物新品种培育重大专项实施以来,大豆转基因新品种培育涉及抗除草剂、抗逆、抗病虫、产量和品质等性状,其中实验室研究任务推进较快,但田间安全评价进度受阻。主要困难集中表现:①大豆对光周期敏感,遗传转化具有基因型选择性,且不同大豆产区只能选择当地推广的大豆品种为受体材料,转基因大豆品系异地异季联合鉴定无法准确反映目标性状的客观表现;②缺乏标准的且符合目标性状的田间鉴定要求的场所及场区,致使安全评价申报和执行滞后;③对转基因大豆目标性状的田间种植,目标基因在DNA、RNA、蛋白水平的检测标准时常处于变化之中,对生态环境因素和土壤微生物的检测对象和检测指标不明确。因此,需要通过立法、加大投入,在全国各省/自治区加强农业部转基因检测中心的基础设施和人才队伍建设,把转基因大豆田间安全评价委托转基因检测中心执行,做到转基因大豆种植标准化、性状观测标准化、中间试验和环境释放评价标准化。
4.3 建立完备的转基因大豆食用安全性评价体系
转基因大豆已成为我国民众直接食用(大豆油)和间接食用(含转基因大豆成分的豆制品等)的主体成分之一,大豆加工剩下的豆粕成为养殖业精饲料的重要组分,肉蛋奶等制品的食用安全性备受民众关注。目前,转基因大豆的食用安全性多采用饲喂动物的实质性等同的法则进行评价,相关试验均由项目承担单位实施,试验标准不统一,试验结果千差万别。在转基因大豆的种植标准,饲喂动物的选择、检测指标及标准、动物本身的体征状态、饲喂和检测时间的设置,检测机构、检测手段、检测机构等方面均缺乏明确的标准及相关规定。因此,农业农村部需要制定相关的法律法规,建立并规范转基因大豆食用安全性的评价体系。
4.4 加强转基因科普宣传 转基因技术的发展已经在农业生产、动物饲养、医学、环境保护等领域发挥重要的作用,这是一个不争的事实。我国要保障粮食安全、实现农业现代化,转基因技术的应用具有广阔的前景。然而,我国公众因不直接从事转基因研究,缺乏转基因安全性基本知识,加之某些媒体、公众人物及组织对转基因安全的“妖魔化”宣传,致使公众对转基因技术和食品安全广泛担忧[2]。作为一个新生事物,公众对转基因技术和转基因食品安全性的认识需要一个过程,存在疑虑和担心符合从众的心理。建议政府部门能够在主流媒体和各单位网站上开辟专栏“加强农业转基因生物技术科学普及”(2015 年中央一号文件),介绍转基因基本知识,并及时更新转基因研究进展。从科研、产业化、行业管理、学校教育、对公众的普法宣传等多个层面统筹,加强舆论和科普宣传建设,促进转基因技术服务农业现代化建设。
参考文献
[1] KAMTHAN A,CHAUDHURI A,KAMTHAN M,et al.Genetically modified(GM) crops:Milestones and new advances in crop improvement [J].Theoretical and applied genetics,2016,129(9):1639-1655.
[2] 王立平,王东,龚熠欣,等.国内外转基因农产品食用安全性研究进展与生产现状[J].中国农业科技导报,2018,20(3):94-103.
[3] 赵雨佳,范培蕾,梁亮,等.转基因作物的发展与检测分析[J].计量技术,2019(10):54-57.
[4] ISAAA.ISAAA in 2018:Accomplishment report[R].2018.
[5] SOUTH P F,CAVANAGH A P,LIU H W,et al.Synthetic glycolate metabolism pathways stimulate crop growth and productivity in the field [J].Science,2019,363(6422):1-9.
[6] DUKE S O.Perspectives on transgenic,herbicide-resistant crops in the United States almost 20 years after introduction[J].Pest manage science,2015,71(5):652-657.
[7] MIKI B,MCHUGH S.Selectable marker genes in transgenic plants:Applications,alternatives and biosafety [J].Journal of biotechnology,2004,107(3):193-232.
[8] BERMAN K H,HARRIGAN G G,RIORDAN S G,et al.Compositions of seed,forage,and processed fractions from insect-protected soybean MON 87701 are equivalent to those of conventional soybean [J].Journal of agricultural & food chemistry,2009,57(23):11360-11369.
[9] BEAZLEY K A,BURNS W C,ROBERT H C,et al.Soybean transgenic event MON 87751 and methods for detection and use thereof:US 9719145[P].2017-01-08.
[10] 谭巍巍,王永斌,赵远玲,等.全球转基因大豆发展概况[J].大豆科技,2019(4):34-38.
[11] MCNAUGHTON J,ROBERTS M,SMITH B,et al.Comparison of broiler performance when fed diets containing event DP-3O5423-1,nontransgenic near-isoline control,or commercial reference soybean meal,hulls,and oil [J].Poultry science,2008,87(12):2549-2561.
[12] 崔寧波,张正岩.转基因大豆研究及应用进展[J].西北农业学报,2016,25(8):1111-1124.
[13] GUTTIKONDA S K,VALLIYODAN B,NEELAKANDAN A K,et al.Overexpression of AtDREB1D transcription factor improves drought tolerance in soybean[J].Mol Biol Rep,2014,41:7995-8008.
[14] KIM H J,CHO H S,PAK J H,et al.Confirmation of drought tolerance of ectopically expressed AtABF3 gene in soybean[J].Mol Cells,2018,41:413-422.
[15] SHI W Y,DU Y T,MA J,et al.The WRKY transcription factor GmWRKY12 confers drought and salt tolerance in soybean[J].Int J Mol Sci,2018,19:1-20.
[16] CHEN L M,FANG Y S,ZHANG C J,et al.GmSYP24,a putative syntaxin gene,confers osmotic/drought,salt stress tolerances and ABA signal pathway[J].Sci Rep,2019,9:1-12.
[17] SU H G,LI B,SONG X Y,et al.Genome-wide analysis of the DYW subgroup PPR gene family and identification of GmPPR4 responses to drought stress[J].Int J Mol Sci,2019,20:1-12.
[18] WEI W,LIANG D W,BIAN X H,et al.GmWRKY54 improves drought tolerance through activating genes in abscisic acid and Ca2+ signaling pathways in transgenic soybean[J].Plant J,2019,100(2):384-398.
[19] ZHANG L,LI T,WANG Y,et al.FvC5SD overexpression enhances drought tolerance in soybean by reactive oxygen species scavenging and modulating stress-responsive gene expression[J].Plant Cell Rep,2019,38(9):1039-1051.
[20] HE Y X,YANG X D,XU C,et al.Overexpression of a novel transcriptional repressor GmMYB3a negatively regulates salt-alkali tolerance and stress-related genes in soybean[J].Biochem Biophys Res Commun,2018,498(3):586-591.
[21] HE Y X,DONG Y S,YANG X D,et al.Functional activation of a novel R2R3-MYB protein gene,GmMYB68,confers salt-alkali resistance in soybean (Glycine max L.)[J].Genome,2020,63(1):13-26.
[22] WANG Y J,JIANG L,CHEN J Q,et al.Overexpression of the alfalfa WRKY11 gene enhances salt tolerance in soybean[J].PLoS One,2018,13:1-16.
[23] DONG L D,CHENG Y X,WU J J,et al.Overexpression of GmERF5,a new member of the soybean EAR motif-containing ERF transcription factor,enhances resistance to Phytophthora sojae in soybean[J].J Exp Bot,2015,66(9):2635-2647.
[24] CHENG Q,LI N H,DONG L D,et al.Overexpression of soybean isoflavone reductase (GmIFR) enhances resistance to Phytophthora sojae in soybean[J].Front Plant Sci,2015,6:1-11.
[25] DU Q,YANG X D,ZHANG J H,et al.Over-expression of the Pseudomonas syringae harpin-encoding gene hrpZm confers enhanced tolerance to Phytophthora root and stem rot in transgenic soybean[J].Transgenic Res,2018,27:277-288.
[26] FAN S J,DONG L D,HAN D,et al.GmWRKY31 and GmHDL56 enhances resistance to Phytophthora sojae by regulating defense-related gene expression in soybean[J].Front Plant Sci,2017,8:1-18.
[27] LI N H,ZHAO M,LIU T F,et al.A novel soybean dirigent gene GmDIR22 contributes to promotion of lignan biosynthesis and enhances resistance to Phytophthora sojae[J].Front Plant Sci,2017,8:1-12.
[28] NIU L,YANG J,ZHANG J H,et al.Introduction of the harpinXooc-encoding gene hrf2 in soybean enhances resistance against the oomycete pathogen Phytophthora sojae[J].Transgenic Res,2019,28:257-266.
[29] WANG L,WANG H Y,HE S F,et al.GmSnRK1.1,a sucrose non-fermenting-1(SNF1)-related protein kinase,promotes soybean resistance to Phytophthora sojae[J].Front Plant Sci,2019,10:1-15.
[30] ZHAO Y L,CHANG X,QI D Y,et al.A novel soybean ERF transcription factor,GmERF113,increases resistance to Phytophthora sojae infection in soybean[J].Front Plant Sci,2017,8:1-16.
[31] MENG F L,LI Y,ZANG Z Y,et al.Expression of the double-stranded RNA of the soybean pod borer Leguminivora glycinivorella (Lepidoptera:Tortricidae) ribosomal protein P0 gene enhances the resistance of transgenic soybean plants[J].Pest Manag Sci,2017,73(12):2447-2455.
[32] LIN J Y,MAZAREI M,ZHAO N,et al.Transgenic soybean overexpressing GmSAMT1 exhibits resistance to multiple-HG types of soybean cyst nematode Heterodera glycines[J].Plant Biotechnol J,2016,14:2100-2109.
[33] WANG Z C,LI T Y,NI H J,et al.Transgenic soybean plants expressing Spb18S dsRNA exhibit enhanced resistance to the soybean pod borer Leguminivora glycinivorella (Lepidoptera:Olethreutidae)[J].Arch Insect Biochem Physiol,2018,98:1-9.
[34] CHEN L,ZHENG Y H,DONG Z M,et al.Soybean (Glycine max) WRINKLED1 transcription factor,GmWRI1a,positively regulates seed oil accumulation[J].Mol Genet Genomics,2018,293(2):401-415.
[35] 余舒斐.轉基因食品安全问题的认识与管理[J].现代食品,2018(21):73-75.
[36] 路超,袁建琴,马艳琴,等.转基因大豆的食用安全性研究进展[J].农学学报,2015,5(12):82-85.
[37] HAMMOND B G,VICINI J L,HARTNELL G F,et al.The feeding value of soybeans fed to rats,chickens,catfish and dairy cattle is not altered by genetic incorporation of glyphosate tolerance[J].J Nutr,1996,126(3):717-727.
[38] CROMWELL G L,LINDEMANN M D,RANDOLPH J H,et al.Soybean meal from roundup ready or conventional soybeans in diets for growing-finishing swine[J].J Anim Sci,2002,80(3):708-715.
[39] JENNINGS J C,KOLWYCK D C,KAYS S B,et al.Determining whether transgenic and endogenous plant DNA and transgenic protein are detectable in muscle from swine fed Roundup Ready soybean meal[J].J Anim Sci,2003,81(6):1447-1455.
[40] ZHU Y Z,LI D F,WANG F L,et al.Nutritional assessment and fate of DNA of soybean meal from roundup ready or conventional soybeans using rats[J].Arch Anim Nutr,2004,58(4):295-310.
[41] SANDEN M,BERNTSSEN M H G,KROGDAHL A,et al.An examination of the intestinal tract of Atlantic salmon,Salmo salar L.,parr fed different varieties of soy and maize[J].J Fish Dis,2005,28(6):317-330.
[42] BAKKE-MCKELLEP A M,KOPPANG E O,GUNNES G,et al.Histological,digestive,metabolic,hormonal and some immune factor responses in Atlantic salmon,Salmo salar L.,fed genetically modified soybeans[J].J Fish Dis,2007,30(2):65-79.
[43] BAKKE-MCKELLEP A M,SANDEN M,DANIELI A,et al.Atlantic salmon (Salmo salar L.) parr fed genetically modified soybeans and maize:Histological,digestive,metabolic,and immunological investigations[J].Res Vet Sci,2008,84(3):395-408.
[44] TUDISCO R,MASTELLONE V,CUTRIGNELLI M I,et al.Fate of transgenic DNA and evaluation of metabolic effects in goats fed genetically modified soybean and in their offsprings[J].Animal,2010,4(10):1662-1671.
[45] MORERA P,BASIRIC L,RONCHI B,et al.Fate of transgenic deoxyribonucleic acid fragments in digesta and tissues of rabbits fed genetically modified soybean meal[J].J Anim Sci,2016,94(3):1287-1295.
[46] 金紅,孙琪,张斌,等.利用蛋白质SDS-PAGE电泳方法检测转基因大豆的初步研究[J].食品研究与开发,2010,31(5):148-150,156.
[47] 曹柏营,欧仕益,黄才欢,等.抗草甘膦转基因大豆的酚类物质分析[J].食品科学,2006,27(8):56-59.
[48] 龙伟,沈秀,周晓靓,等.新体系评价转基因大豆油的食用安全性[J].食品安全质量检测学报,2014,5(8):2568-2574.
[49] 朱元招.抗草甘膦大豆转基因PCR检测及其饲用安全研究[D].北京:中国农业大学,2004.
[50] APPENZELLER L M, MUNLEY S M, HOBAN D, et al.Subchronic feeding study of herbicide-tolerant soybean DP-35643-5 in Sprague-Dawley rats[J].Food Chem Toxicol,2008,46(6):2201-2213.
[51] 芦春斌,张伟,刘标.抗草甘膦转基因大豆饲料对雄性小鼠脾淋巴细胞体外增殖的影响[J].大豆科学,2012,31(2):291-294.
[52] MALATESTA M,BORALDI F,ANNOVI G, et al.A long-term study on female mice fed on a genetically modified soybean:Effects on liver ageing[J].Histochem Cell Biol,2008,130(5):967-977.
[53] 郭斌,陈勇,黄斌.转基因大豆食品安全性及对生态环境影响初探[J].农业环境与发展,2002(6):29-31.
[54] BERBERICH S A,REAM J E.Safety assessment of insect-protected cotton:The composition of the cottonseed is equivalent to conventional cotton seed[J].J Agric Food Chem,1996,41:365-371.
[55] 索海翠,熊瑞权,马启斌,等.转基因大豆的潜在风险及发展趋势研究[J].广东农业科学,2010(10):244-245.
[56] 刘刚.抗草甘膦转基因大豆不具有杂草化的潜力[J].农药市场信息,2017(28):52.
[57] ABE J,KANAZAWA A,SHIMAMOTO Y.Genetic diversity and in situ conservation of wild soybeans:Report of grants-in-aid for scientific research from the J.S.P.S [M].Sapporo:Hokkaido University,2000:3-24.
[58] 陳新,王长永,朱成松,等.转基因抗草甘膦大豆安全性评价及对环境影响的检测[J].江苏农业科学,2003(6):36-40.
[59] 刘琦,李希臣,刘昭军,等.抗草甘膦转基因大豆基因漂移的研究Ⅰ大豆风媒介传粉的基因漂移研究[J].黑龙江农业科学,2008(1):14-16.
[60] 吕晓波,王宏燕,刘琦,等.抗草甘膦转基因大豆(RRS)在黑土生态系统种植的安全性研究[J].大豆科学,2009,28(2):260-265.
[61] 唐影,李世东,缪作清.转基因作物对土壤微生物的影响[J].中国生物防治,2007,23(4):383-390.
[62] MEANS N E,KREMER R J,RAMSIER C.Effects of glyphosate and foliar amendments on activity of microorganisms in the soybean rhizosphere[J].J Environ Sci Health B,2007,42(2):125-132.
[63] 杜伟,黄启星,左娇,等.南繁条件下转基因大豆对根际土壤可培养微生物的影响[J].热带作物学报,2012,33(3):417-421.
[64] 王争,王宏燕,邵婧鑫.多年种植RRS对根际土壤微生物数量及氮素转化的影响[J].东北农业大学学报,2012,43(2):92-97.
[65] 朱银玲.转EPSPS基因抗除草剂大豆对土壤原核微生物群落生态影响的研究[D].南京:南京大学,2015.
[66] 章秋艳,李刚,杨志国.转基因大豆种植对根际土壤酶活性和养分的影响[J].中国油料作物学报,2014,36(3):409-413.
[67] 左娇,郭运玲,孔华,等.转基因大豆安全性评价的研究进展[J].热带作物学报,2013,34(7):1402-1407.
猜你喜欢
中国生殖健康(2019年5期)2019-01-06
现代营销(创富信息版)(2018年7期)2018-09-05
现代营销(创富信息版)(2018年6期)2018-09-05
现代营销(创富信息版)(2018年5期)2018-07-12
农家科技下旬刊(2016年9期)2016-12-15
科学与财富(2016年15期)2016-11-24
饲料博览(2016年8期)2016-04-06
