
刺槐幼苗对NaCl胁迫的生理生化响应
2020-09-09 07:16甘红豪杨泽坤褚建民
林业科学研究 2020年4期
甘红豪,赵 帅,杨泽坤,褚建民
(国家林业和草原局滨海林业研究中心,中国林业科学研究院林业研究所,国家林业和草原局林木培育重点实验室,林木遗传育种国家重点实验室,北京 100091)
土壤中过量的盐分会抑制植物光合作用、诱导产生氧化胁迫、渗透胁迫及离子胁迫等,阻碍植物的正常生长发育进程[1-2]。植物通过合成积累游离脯氨酸、氨基酸和可溶性蛋白质等渗透调节物质,并诱导增强植物体内抗氧化酶如过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)和谷胱甘肽还原酶(GR)等的活性,从而增强了植物的耐盐能力[3-4]。
水分是植物在盐胁迫下维持正常生长的重要条件。液泡膜内在蛋白(TIPs)和质膜内在蛋白(PIPs)作为植物水通道蛋白(AQPs)家族的重要成员[5],通过调节植物水分运输,影响植物逆境应答,促进盐胁迫下植物生长发育[4]。盐胁迫下,转TsTIP1;1及TsPIP1;1基因水稻(Oryza sativaL.)的光合能力显著增强、细胞持水能力明显提高[5];过表达PIP2;1增强了拟南芥(Arabidopsisthaliana(L.) Heynh.)细胞的持水能力[6]。位于液泡膜上的Na+/H+逆向转运蛋白NHX1参与调控植物液泡渗透势,可以把细胞质内过量的Na+隔离到液泡内,减轻了拟南芥受到的盐胁迫,从而减弱了盐胁迫对其生长的抑制[7]。
刺槐(Robinia pseudoacaciaL.)生长迅速、适应性强,具有耐干旱、耐瘠薄、繁殖快等特点,是盐碱地区造林的先锋树种,具有良好的经济和环境效益[8-9]。近年从逆境生理、品种选育等方面对刺槐的耐盐机理进行了一定研究,然而,盐胁迫下各生理指标对刺槐耐盐能力的影响及刺槐体内相关基因转录表达情况的研究较少。本研究以1年生刺槐幼苗为试验材料,综合分析NaCl胁迫对刺槐幼苗生长、生理、离子含量及相关基因表达的影响,初步解析刺槐耐盐机制,从而为建立刺槐抗逆性生态生理指标体系提供理论依据。
1 材料与方法
1.1 试验材料及处理方法
以1年生刺槐播种苗为试验材料,在中国林业科学研究院科研温室中进行盆栽(花盆规格:盆高×盆口直径=22 cm×20 cm)培养,基质为V珍珠岩∶V蛭石=3∶1。培养条件为:自然光照;昼夜温度:25/20°C;相对湿度:50%~60%。试验开始前,每株幼苗每2 d浇灌1/2 Hoagland 营养液(pH值5.5)100 mL,待苗高约20 cm时,选取生长一致且健壮的幼苗进行试验。
试验分3个处理组,每个处理组10株幼苗,处理浓度分别为0、50、100 mmol·L-1NaCl溶液。NaCl处理期间,为保证NaCl浓度的一致性,每3 d用含同等浓度NaCl溶液的1/2 Hoagland营养液淋洗花盆基质,处理27 d后,每个处理组选取6株长势基本一致的幼苗进行收获。
1.2 测定方法
1.2.1 光合参数测定及材料收获 收获前,每个处理组随机选取3株幼苗,每株选取3片成熟的叶片,在晴天上午 9: 00—11: 00 用 Li 6 400光合作用测量系统 ( LI-Cor, Lincoln, Nebraska, USA) 测定刺槐幼苗叶片的光合作用参数;同时,计算刺槐叶片水分利用效率(WUE) = 净光合速率(A)/蒸腾速率(E)[10]和气孔限制值(Ls) = 1-Ci/Ca(Ci为胞间CO2浓度;Ca为空气中CO2浓度,本试验中该值为400 μmol·mol-1)[11]。
光合测定结束后,收获植株。收获时,根、茎、叶分别收获并记录各部分的生物量。所有材料保存在液氮中,并在液氮中使用球磨仪(NM400,Retsch, Haan, Germany)将其研成粉末,于-80°C保存备用。称取各处理组刺槐根、茎和叶片鲜样60 mg,于65°C烘干,计算样品干湿比及干质量。
1.2.2 叶片相对含水量和叶绿素含量测定 采用烘干称重法测定各处理组刺槐幼苗叶片相对含水量(RWC)。参照Wellburn[12]提出的方法测定刺槐幼苗叶片中叶绿素含量。
1.2.3 丙二醛(MDA)含量测定 根据 Hodges等[13]提出的方法测定刺槐样品中丙二醛(MDA)的含量。
1.2.4 游离脯氨酸、氨基酸、可溶性蛋白质含量测定 根据Tamás等[14]的方法测定样品游离脯氨酸含量。使用氨基酸(AA)含量检测试剂盒(索莱宝,北京)测定样品中氨基酸含量。样品中可溶性蛋白含量参照Luo等[15]的方法测定。
1.2.5 酶活性测定 参照Polle等[16]的方法测定CAT和APX的活性;参照 Gamble等[17]的方法测定GR的活性。
1.2.6 离子含量分析 将烘干的根和叶片样品研磨粉碎后,过0.5 mm筛,称取0.2~0.3 g样品(精确至0.001 g),置于消煮管中,加入8 mL浓硝酸,摇匀后过夜,并做空白对照。消解前加入2 mL H2O2,静置20 min,随后使用Mars高压微波消解仪(Mars-6,CEM Corp.,USA)消解,之后加热赶酸,待消解液剩余约 2 mL 时停止加热,冷却后过滤到50 mL容量瓶中并定容、摇匀。静置 20 min,吸取上清液 10 mL 至离心管中,使用等离子体发射光谱仪 iCAP 6 300(Thermo Scientific,USA)测定Na+、K+、Ca2+、Mg2+含量,并计算Na+/K+、Na+/Ca2+、Na+/Mg2+比值。
1.2.7 水通道蛋白及Na+/ H+逆向转运蛋白基因表达分析 使用植物RNA提取试剂盒(DP432,天根)提取刺槐根和叶样品中总RNA,并进行浓度及纯度检测。使用PrimeScript™ RT reagent Kit with gDNA Eraser(RR047B,TaKaRa)将纯化后的RNA进行反转录,获得cDNA,然后使用荧光定 量PCR仪(ABI7500,Applied Biosystems,USA)进行相对荧光定量(qPCR)分析。引物由北京Invitrogen公司合成,具体信息见表1,内参基因为Actin[18]。
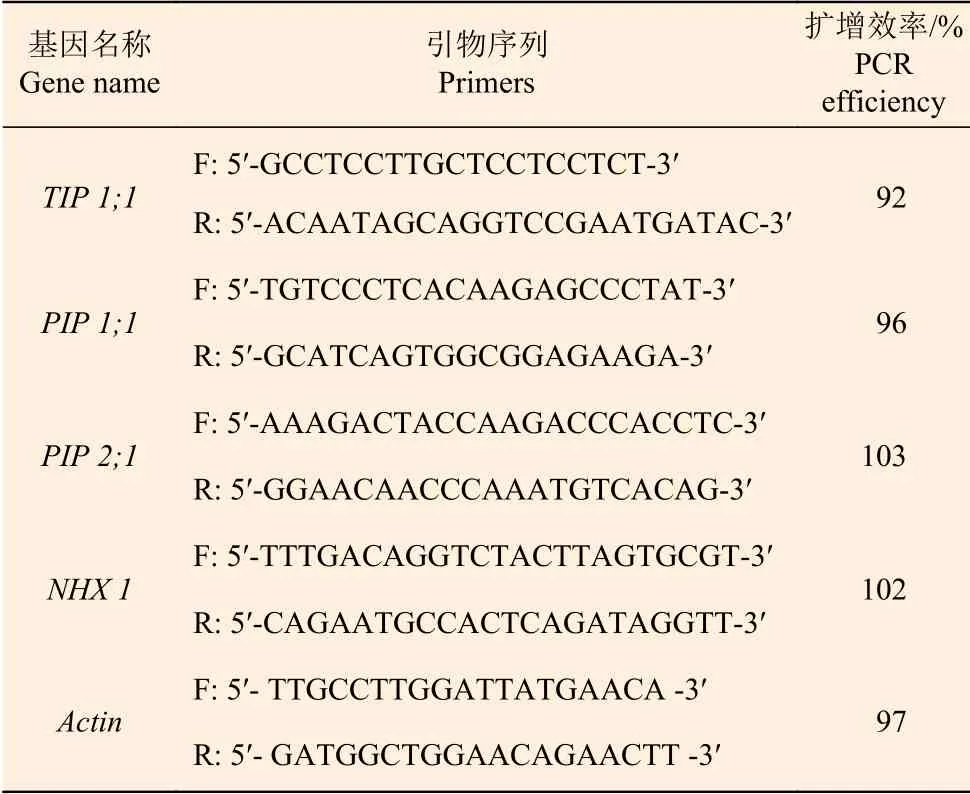
表1 qRT-PCR 引物信息Table 1 Primers used for qRT-PCR
1.3 数据统计分析
采用2-ΔΔCT法获得qPCR的差异表达倍数。对本试验获得的所有数据使用Statgraphics Centurion XVI.I (STN, St, Louis, MO, USA)软件进行分析。数据分析之前,对其进行正态分布检验。所有数据采用单因素进行分析,以NaCl作为自变量因素。采用LSD法进行数据间差异显著性分析,当F检验的P-Value < 0.05时,认为数据间的差异显著。
2 结果与分析
2.1 NaCl处理对刺槐幼苗生长和光合能力的影响
在NaCl处理下,刺槐幼苗根、茎和叶的干质量均出现不同程度的下降,根冠比增加(表2),说明NaCl处理影响了刺槐的正常生长发育。随NaCl浓度的升高,刺槐幼苗叶片相对含水量显著降低,与0 mmol·L-1NaCl处理组相比,50、100 mmol·L-1NaCl处理组刺槐幼苗叶片的相对含水量分别减少了7.0%和18.2%。
NaCl处理使刺槐幼苗叶中叶绿素a(Chl a)、叶绿素b(Chl b)和类胡萝卜素(Car)均呈下降趋势(表3)。与0 mmol·L-1NaCl处理组相比,100 mmol·L-1NaCl 处 理 使 刺 槐 幼 苗 叶 片Chl a、Chl b和Car含量分别降低了29.8%、30.5%和36.6%。刺槐净光合速率(A)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(E)均随着NaCl浓度增加显著减少,而水分利用效率(WUE)和气孔限制值(Ls)出现不同程度的增加(表4)。100 mmol·L-1NaCl 处理 组刺 槐 的A、Gs、Ci和E仅为0 mmol·L-1NaCl处理组刺槐的42.3%、40.0%、84.7%和31.1%。50 mmol·L-1NaCl 处理对刺槐幼苗叶片的WUE和Ls无显著影响;而100 mmol·L-1NaCl 处理组刺槐幼苗叶片的WUE和Ls分别比0 mmol·L-1NaCl 处理组刺槐升高了39.3%和25.0%。
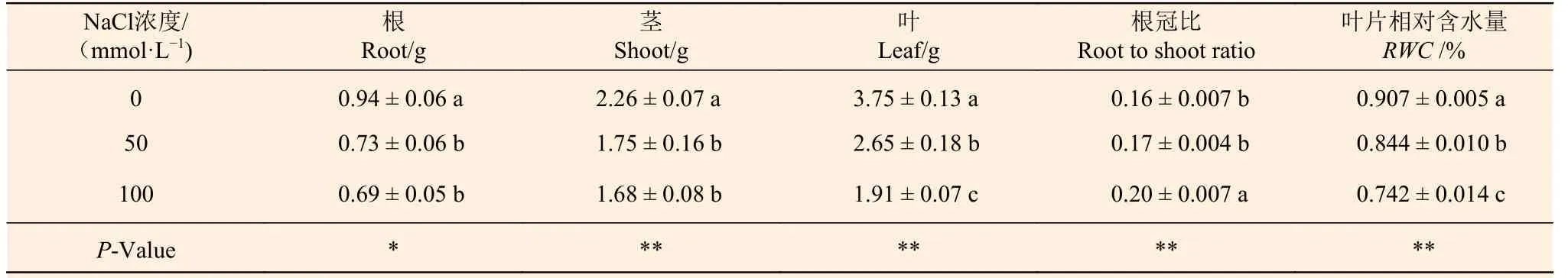
表2 NaCl对刺槐幼苗生物量、根冠比及叶片相对含水量的影响Table 2 The effects of NaCl concentrations on biomass, root to shoot ratio and leaf relative water content of R. pseudoacacia
表3 NaCl对刺槐幼苗光合色素含量及光合作用参数的影响
Table 3 The effects of NaCl concentrations on photosynthetic pigment content ofR. pseudoacacia

NaCl浓度/(mmol·L-1)叶绿素a Chl a/(mg g-1)叶绿素b Chl b/(mg·g-1)类胡萝卜素Car/(mg·g-1)叶绿素a+b Chl a+b/(mg g-1)0 9.26 ± 0.80 a 2.69 ± 0.21 a 2.02 ± 0.18 a 11.95 ± 1.02 a 50 7.92 ± 0.05 ab 2.27 ± 0.10 ab 1.48 ± 0.04 b 10.19 ± 0.15 ab 100 6.50 ± 0.05 b 1.87 ± 0.05 b 1.28 ± 0.05 b 8.38 ± 0.11 b P-Value * * ** *

表4 NaCl对刺槐幼苗光合作用参数、水分利用效率及气孔限制的影响Table 4 The effects of NaCl concentrations on photosynthetic parameters, WUE and Ls of R. pseudoacacia
2.2 NaCl处理对刺槐幼苗丙二醛(MDA)含量的影响
表5表明:在50 mmol·L-1NaCl处理下,刺槐根和叶中MDA含量均是0 mmol·L-1NaCl处理组的1.2倍;与0 mmol·L-1NaCl处理相比,100 mmol·L-1NaCl处理组刺槐幼苗根和叶中MDA含量虽然出现一定程度增加,但差异不显著。
2.3 NaCl处理对刺槐幼苗CAT、APX和GR活性的影响
表5表明:随NaCl浓度的增加,CAT、APX和GR的活性变化不同;在根中,与0 mmol·L-1NaCl 处理组刺槐相比,50 mmol·L-1NaCl处理对CAT和GR活性无显著影响,但使APX活性显著升高;100 mmol·L-1NaCl处理使CAT、APX和GR活性显著升高。在叶中,50 mmol·L-1NaCl处理使CAT活性显著升高,但对APX和GR活性无显著影响;100 mmol·L-1NaCl处理组刺槐CAT、APX和GR活性分别是0 mmol·L-1NaCl 处理组的3.0、1.8、1.7倍。
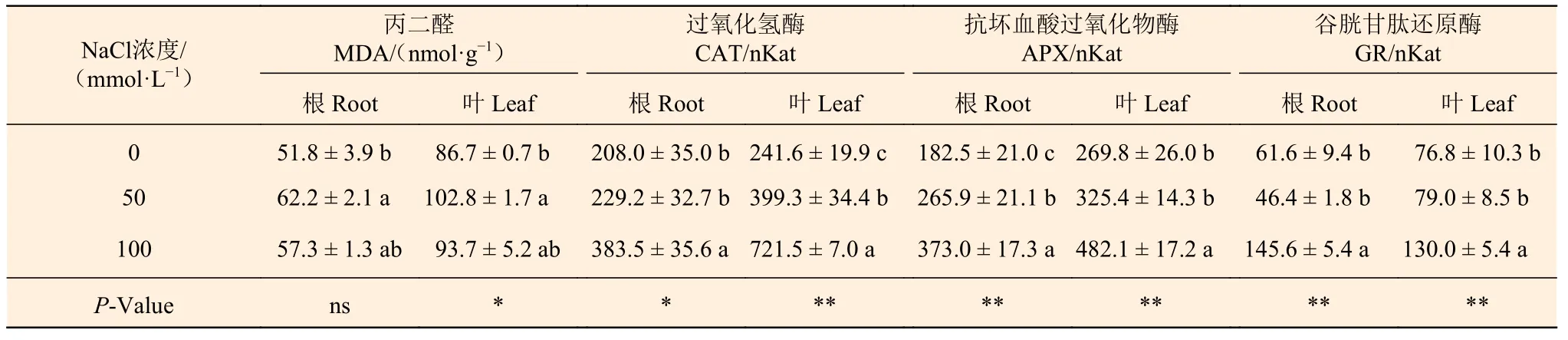
表5 NaCl对刺槐幼苗根和叶丙二醛含量,过氧化氢酶、抗坏血酸过氧化物酶和谷胱甘肽还原酶活性的影响Table 5 The effects of NaCl on MDA content, CAT, APX and GR activities in roots and leaves of R. pseudoacacia
2.4 NaCl处理对刺槐幼苗游离脯氨酸、氨基酸及可溶性蛋白质的影响
NaCl处理导致刺槐幼苗根和叶中游离脯氨酸、氨基酸和可溶性蛋白质出现不同程度升高(表6)。在根中,与0 mmol·L-1NaCl 处理组相比,50 mmol·L-1NaCl处理使刺槐幼苗游离脯氨酸、氨基酸和可溶性蛋白质分别升高了72.1%、86.4%和63.2%;100 mmol·L-1NaCl处理使刺槐幼苗游离脯氨酸升高了74.4%,可溶性蛋白质升高了156.7%,但对氨基酸含量影响不显著。在叶中,与0 mmol·L-1NaCl 处理组相比,50 mmol·L-1NaCl处理使刺槐幼苗游离脯氨酸和可溶性蛋白质分别升高了136.4%和64.3%;100 mmol·L-1NaCl处理使刺槐幼苗游离脯氨酸升高了381.8%,氨基酸升高了21.3%。
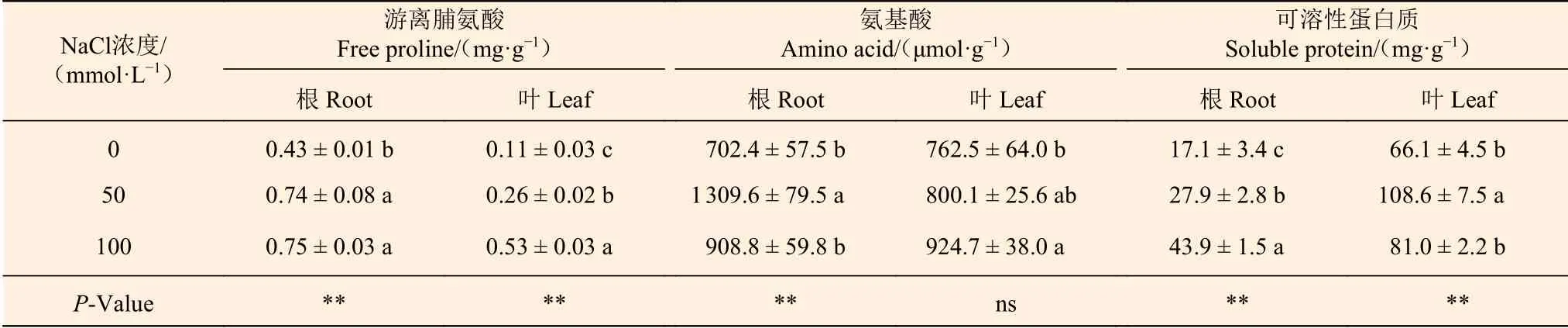
表6 NaCl对刺槐幼苗根和叶中游离脯氨酸、氨基酸及可溶性蛋白质含量的影响Table 6 The effects of NaCl on free proline, amino acid and soluble protein contents in roots and leaves of R. pseudoacacia
2.5 NaCl处理对刺槐幼苗Na+、K+、Mg2+ 和Ca2+含量及离子含量比值的影响
表7表明:随NaCl浓度的增加,刺槐幼苗根和叶中Na+含量显著升高,50 mmol·L-1NaCl处理组刺槐根和叶中Na+是0 mmol·L-1NaCl处理组的2.5和3.1倍,100 mmol·L-1NaCl处理组刺槐根和叶中Na+是0 mmol·L-1NaCl处理组的4.6和5.6倍,且根中Na+含量高于叶中含量。50、100 mmol·L-1NaCl处理使刺槐幼苗根中K+含量比0 mmol·L-1NaCl处理组分别降低了16.3% 和11.1%,但对叶中K+含量影响不显著。NaCl处理使刺槐幼苗根中Mg2+含量逐渐升高,而叶中Mg2+含量逐渐降低,与0 mmol·L-1NaCl 处理组相比,100 mmol·L-1NaCl 处理使根中Mg2+含量升高了29.7%,使叶中Mg2+含量下降了28.2%。NaCl处理对刺槐幼苗根和叶中Ca2+含量影响不显著。
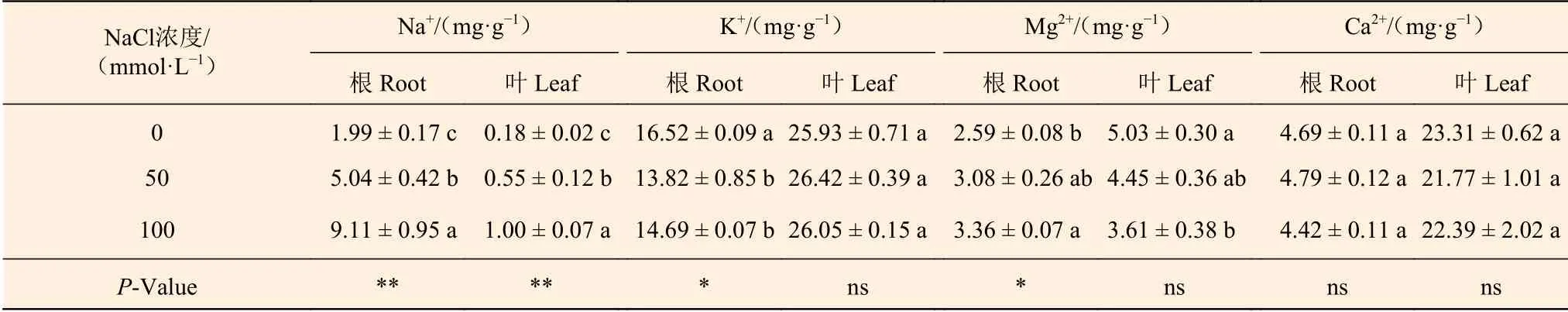
表7 NaCl对刺槐幼苗根和叶中离子含量的影响Table 7 The effects of NaCl on ion contents in roots and leaves of R. pseudoacacia
NaCl处理下,刺槐幼苗根和叶中Na+/K+、Na+/Mg2+和Na+/Ca2+显著升高,且NaCl浓度越高,比值越大(表8)。50 mmol·L-1NaCl处理使根中Na+/K+、Na+/Mg2+和Na+/Ca2+分别升高了200.0%、110.3%和150.0%,使叶中Na+/K+、Na+/Mg2+和Na+/Ca2+分别升高了100.0%、200.0%和100.0%。与0 mmol·L-1NaCl处理组刺槐相比,100 mmol·L-1NaCl处理组刺槐根中Na+/K+、Na+/Mg2+和Na+/Ca2+分别升高了4.2、2.5和3.9倍,叶中Na+/K+、Na+/Mg2+和Na+/Ca2+分别升高了3.0、6.0和3.0倍。
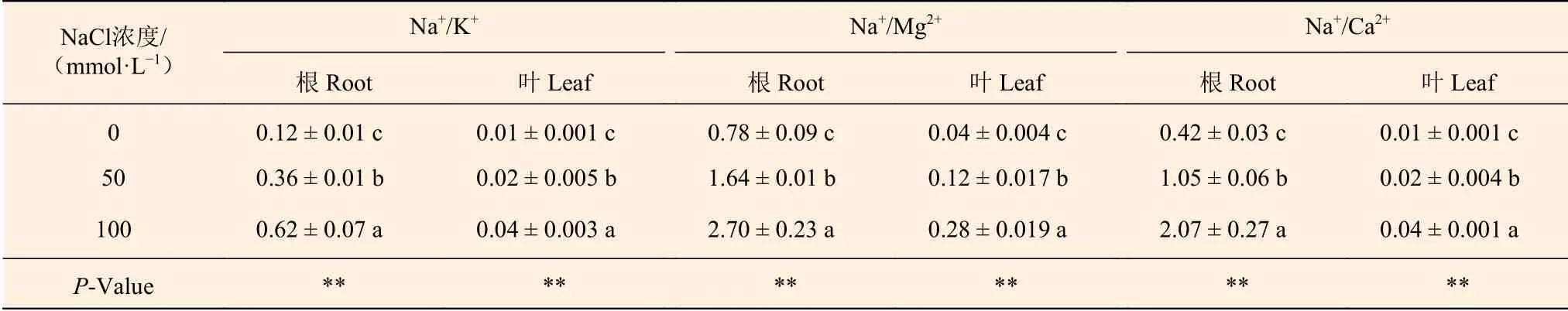
表8 NaCl对刺槐幼苗根和叶中Na+/K+、Na+/Mg2+ 和Na+/Ca2+ 的影响Table 8 The effects of NaCl on Na+/K+, Na+/Mg2+ and Na+/Ca2+ in roots and leaves of R. pseudoacacia
2.6 NaCl处理对刺槐幼苗水通道蛋白及Na+ / H +逆向转运蛋白基因表达的影响
对刺槐幼苗根和叶中编码水通道蛋白(TIP1;1,PIP1;1和PIP2;1)及Na+/H+逆向转运蛋白(NHX1)的mRNA 的表达水平进行分析,结果(表9)表明:在刺槐幼苗根中,50和100 mmol·L-1NaCl处理使TIP1;1、PIP1;1、PIP2;1和NHX1基因的mRNA 表达水平显著升高,分别是0 mmol·L-1NaCl处理组的6.4、11.1、28.6、6.4倍和5.6、6.1、14.6、3.2倍;在叶中,与0 mmol·L-1NaCl处理组刺槐幼苗相比,50 mmol·L-1NaCl处理对TIP1;1、PIP1;1和NHX1基因的mRNA 表达水平影响不显著,100 mmol·L-1NaCl处理使TIP1;1、PIP1;1和NHX1基因的mRNA表达水平增加了3.7、1.6和1.3倍,但NaCl处理使PIP2;1基因的mRNA 表达水平显著降低。

表9 NaCl处理对刺槐幼苗根和叶中相关基因表达分析Table 9 The expression of aquaporin and Na+ / H + reverse transporter genes in roots and leaves of R. pseudoacacia under NaCl treatments.
3 讨论
盐胁迫是限制植物生长发育的主要非生物逆境之一[2]。本研究中,NaCl处理显著抑制了刺槐幼苗的生长发育,致使刺槐幼苗根、茎和叶的干质量减少,根冠比增加,表明刺槐幼苗在受到NaCl胁迫后,会通过抑制地上部分生长、扩大根冠比,从而增强根系对土壤中水和营养元素的吸收,以此来适应逆境条件[19-20]。相对含水量作为衡量植株水分状况的指标之一,常用来指示植物的逆境胁迫程度。本研究发现,NaCl处理使刺槐叶片相对含水量下降,这可能与盐胁迫降低了植物根系向地上部分运输水分的能力有关[21]。
叶绿素是植物进行光合作用的主要色素,其含量对植物的耐盐能力具有重要影响[22]。本研究中,在NaCl处理下,刺槐幼苗叶片的Chl a、Chl b、Chl a+b及Car含量均出现不同程度降低。这是由于过量的NaCl破坏了植物的叶绿体结构、提高了相关叶绿素酶的活性,导致叶绿体色素合成受阻或者分解加快[22]。刺槐幼苗叶片净光合速率(A)在受到NaCl胁迫后下降,光合能力减弱。一般认为,NaCl胁迫影响植物光合作用的因素主要分为气孔因素和非气孔因素,前者表现为Gs和Ci同时下降、Ls升高,后者主要表现为A降低而Ci升高[11]。本研究中,随NaCl浓度增加,刺槐幼苗叶片A、Gs、Ci和E均显著减少,Ls升高,说明NaCl胁迫对刺槐幼苗光合能力的限制主要由气孔因素导致。水分利用效率(WUE)反映了植物对环境适应能力的强弱,NaCl处理使刺槐叶片的WUE升高,说明在一定浓度范围内,NaCl胁迫能够提高刺槐的WUE,这可能是由于NaCl胁迫诱导刺槐幼苗叶片Gs和E降低减少了蒸腾耗水引起的[23]。
研究表明,在一定盐浓度范围内,白榆(Ulmus pumilaL.)[24]、中国柽柳(Tamarix chinensisLour.)[25]等体内抗氧化酶活性均随着盐浓度的升高而增大。本研究同样发现,NaCl胁迫诱导刺槐幼苗根和叶中CAT、APX和GR活性增强,从而加快清除植株体内过量的活性氧。在NaCl胁迫下,刺槐幼苗通过合成并积累游离脯氨酸、氨基酸和可溶性蛋白质,从而调节渗透势、维持细胞含水量,提高刺槐抗氧化能力,增强了对NaCl胁迫的适应能力,这与在茶树(Camellia sinensisL.)[26]中得到的结果相似。NaCl处理导致刺槐幼苗根和叶中丙二醛(MDA)含量升高,并且100 mmol·L-1NaCl处理组刺槐MDA含量低于50 mmol·L-1NaCl处理组,这可能是在较高浓度NaCl胁迫下,刺槐体内抗氧化系统活性较高,能够较快的清除活性氧物质,减轻了其对刺槐细胞膜系统的损伤。
本研究中,NaCl 处理使刺槐幼苗根和叶中Na+积累量显著升高,且Na+在刺槐幼苗根中的积累量远大于在叶中的积累量。在多数植物中,Na+的主要毒性部位是叶片[2],因此,植物根系对Na+的留存能力可以减轻Na+对地上部分的伤害;同时,植物也可以利用根中积累的Na+进行渗透调节,降低植物水势[19]。随着NaCl浓度升高,刺槐幼苗根中K+含量显著减少,Na+/K+显著增加,这可能是由于在盐胁迫下,根系对Na+的吸收会与对K+的吸收产生竞争,导致根中K+含量显著减少所致[3,27];然而,NaCl对刺槐幼苗叶中K+含量无显著影响,但Na+/K+却显著升高,这可能是由于地上部分在NaCl胁迫下生长减弱,引起K+在刺槐根和叶中重新分配所致[27]。Mg2+作为组成植物叶绿素分子的元素之一,本研究发现,NaCl胁迫导致刺槐幼苗根中Mg2+含量上升,叶中Mg2+含量降低,同时根和叶中Na+/Mg2+显著升高,刺槐通过在根中积累Mg2+,提高了渗透调节能力,维持了刺槐幼苗根中质子跨膜运输浓度梯度[28]。研究发现,在较低浓度NaCl 胁迫下,胀果甘草(Glycyrrhiza inflataBatal.)[28]及酸枣(Ziziphus jujubavar.spinosa(Bunge) Hu ex H. F. Chow)[29]的根系通过积累Ca2+提高细胞渗透调节能力,缓解Na+毒害。本研究中,50和100 mmol·L-1NaCl处理对刺槐幼苗根和叶中Ca2+含量无显著影响,从而维持NaCl胁迫下刺槐体内细胞膜结构的稳定、调控细胞生长及胞内酶的活性。
在植物适应盐胁迫的过程中,植物水通道蛋白(AQPs)通过调控植物细胞渗透压及蒸腾作用,调控植物逆境应答,促进植物生长发育[5]。研究发现,NaCl 处理导致水稻根中OsTIP1;1、TsPIP1;1及TsTIP1;1表达水平显著上调[30-31]。本研究中,NaCl处理后,刺槐幼苗根中水通道蛋白基因TIP1;1、PIP1;1及PIP2;1转录被诱导。TIP1;1作为定位在液泡膜上的水通道蛋白,参与植物细胞渗透调节;而PIP1;1和PIP2;1均定位在质膜上,前者主要用于调节植物渗透势,后者主要负责跨膜水分运输。因此,刺槐通过诱导上述3种基因的合成表达,并与渗透调节物质共同作用,调节细胞的渗透势,增强了刺槐根系的吸水及持水能力,增强了刺槐的耐盐能力。在叶中,TIP1;1和PIP1;1转录水平在低浓度NaCl处理下变化不显著,但在较高浓度NaCl处理下显著升高,这可能是由于在较高浓度NaCl下,叶片通过诱导它们的转录表达,增强叶片细胞的渗透调节能力,并抑制PIP2;1的转录表达,减弱了叶片失水速率,减缓了盐胁迫引起的净光合速率的下降速度,从而增强了刺槐的耐盐能力[6]。然而,在玉米中,NaCl胁迫抑制了PIPs和TIPs的表达[32],这可能是与AQPs基因在逆境条件下表达存在组织、器官以及时间上的特异性有关[33]。NHX1是定位于液泡膜上的Na+/H+逆向转运蛋白,参与调控植物液泡渗透势[3]。本研究中,NaCl处理后,刺槐幼苗根和叶中NHX1的表达水平显著升高。刺槐通过增强NHX1基因的转录表达,将根和叶中过量的Na+从细胞质转运到液泡中,减缓了细胞质的受损程度,重建细胞中的离子平衡,并且将Na+转化为渗透调节物质,增强了自身的耐盐能力[34]。
4 结论
NaCl胁迫抑制了刺槐幼苗的生长以及光合作用能力,同时诱导刺槐体内产生氧化胁迫和离子失衡。为应对NaCl胁迫造成的不利影响,刺槐通过增强叶片水分利用效率、合成并积累抗氧化酶(CAT、APX和GR)和渗透调节物质(游离脯氨酸、氨基酸和可溶性蛋白质)并诱导相关基因(TIP1;1、PIP1;1、PIP2;1及NHX1)的转录表达,增强了自身的盐胁迫适应能力。然而,植物对盐胁迫的适应机制非常复杂,还需进一步深入研究。
猜你喜欢
中老年保健(2022年3期)2022-11-21
现代园艺(2020年2期)2020-12-22
科学与财富(2019年15期)2019-10-21
小溪流(故事作文)(2018年10期)2018-12-28
中成药(2018年4期)2018-04-26
现代园艺(2018年2期)2018-03-15
湖南林业科技(2017年1期)2017-02-06
故事作文·低年级(2016年10期)2016-09-10
中成药(2016年4期)2016-05-17
现代食品(2016年14期)2016-04-28
