
浙江仙居长叶榧树、刺叶栎资源现状调查及分析
2020-09-08 02:37:30周炜伦陈水飞臧明月陆丽平方炎明
中国野生植物资源 2020年8期
周炜伦,陈水飞,李 垚,王 璐,赵 荣,臧明月,陆丽平,解 雷,方炎明*
(1. 南京林业大学 生物与环境学院,南方现代林业协同创新中心,亚热带森林生物多样性保护国家林业和草原局重点实验室,江苏 南京 210037;2. 生态环境部南京环境科学研究所 自然保护与生物多样性研究中心,国家环境保护武夷山生态环境科学观测研究站,国家环境保护生物安全重点实验室,江苏 南京 210042)
全球榧属(Torreya)共6~7种,长叶榧树(TorreyajackiiChun)是其最原始的树种,这已得到分子证据的支持[1]。该属具独特的植物地理模式,长叶榧树产浙江南部;榧树(Torreyagrandis)产华东及湘黔地区,加州榧树(Torreyacalifornica)产美国加州,为东亚-北美姐妹种对;巴山榧树(Torreyafargesii)和日本榧树(Torreyanucifera)构成中国-日本姐妹种对;此外,佛罗里达榧树(Torreyataxifolia)产佛罗里达等地,云南榧树(Torreyayunnanensis)产云南,均呈零星分布状[1-2]。中国的榧树均被列为国家Ⅱ级重点保护野生植物。
刺叶栎(QuercusspinosaDavid)属于栎属(Quercus)冬青栎组(Sect.Ilex)[3],高山栎类(原高山栎组Sect.Heterobalanus)分布最低(900~1500 m)、最东的种类,从横断山(川、滇)、中东部地区(甘、陕、鄂、闽、浙、赣),一直延伸到台湾中央山脉[2,4]。刺叶栎是几种主要常绿硬叶栎树之一,主要分布冷温带[5],在东部亚热带高海拔地区呈零星分布,地理模式独特。东部群体具有鲜明的遗传特质,蕴藏特有的叶绿体单倍型[6]。
目前,国内对于两者的研究较少,多集中于繁育栽培及生长环境调查等方面。例如杨胜利等人[7]对长叶榧树开花结果习性及繁育栽培技术的研究,杨兴宏[8]通过对甘肃省成县刺叶栎的生长环境、种群数量和结构特征调查,找出刺叶栎在当地的分布特点,而对于两者植物资源现状的研究较为匮乏。
长叶榧树和刺叶栎地理分布、生态位和遗传基础的特殊性引起了学界的高度关注[9]。仙居县被誉为浙江省罕见的天然植物“基因库”和“植物博物馆”[10]。长叶榧树树和刺叶栎同时分布在仙居县,从生物多样性保护、森林生态系统保护、林木种质资源保护的角度意义重大。据此,我们对仙居县神仙居景区及上岙村长叶榧树和刺叶栎群落进行调查。
1 自然概况
仙居县植物种类丰富。据统计,县内约有种子植物142科,616属,1347种,其中裸子植物5科10属14种,区内被子植物占绝对优势[10]。
本次调查的长叶榧树种群位于浙江省仙居县神仙居景区内。该景区地处白塔镇南部,120°34′16″~120°37′30″E,28°40′26″~28°44′4″N,距仙居县城西南约25 km,区域总面积1450 hm2,其中有林地面积1348.5 hm2,森林覆盖率达93%[11]。神仙居景区属中亚热带季风气候区,光照充足,雨水充沛,每年7~9月有台风影响[10]。区内地质构造独特,是世界上最大的火山流纹岩地貌集群,地貌特征以沿溪湿地和沟谷为主。长叶榧树种群调查点海拔在406~477 m,最高点海拔477 m,属于低山地貌。
此次调查的刺叶栎种群位于朱溪镇上岙村。该村位于浙江省仙居县东南,生态林业态势良好,素有“竹乡”之称。上岙村属亚热带海洋性季风气候区,四季分明、无霜期长、年温差较小,热量条件好,气候条件十分优越,适宜多种经济林生长[12]。当地海拔垂直高度为50~1200 m,刺叶栎种群调查点海拔在1022~1026 m,最高点海拔1026 m,地貌特征以山脊为主,属于中山地貌。
2 调查内容与方法
2.1 调查方法
2019年8月,项目组通过查阅相关文献及标本,在仙居国家公园工作人员的协助下,分别对神仙居景区长叶榧树和上岙村刺叶栎的野外群落进行调查。通过调查群落海拔、经纬度、土壤类型、植物类型和地形、地貌、干扰强度等,全面了解长叶榧树及刺叶栎的野外生长状况。
在神仙居景区、上岙村常绿阔叶林中各设置10 m×10 m的群落调查样方3个,并用手持GPS进行地理坐标定位,记录群落外貌、各层的种类组成和生境特征。对样方内所有胸径≥1 cm的植物进行测量,记录其种类、数量、树高、胸径、冠幅物候期及生长状态,以便于研究这些植物群落样方中的种类组成及群落学特征。
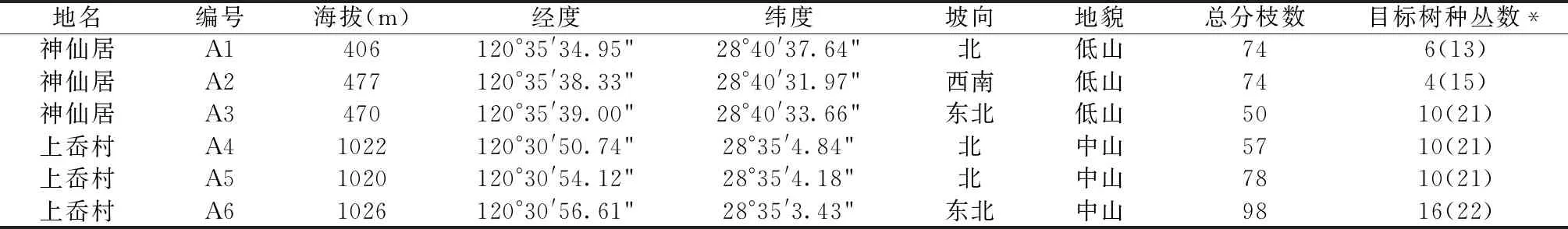
表1 长叶榧树及刺叶栎地理分布与立地现状
2.2 数据处理
参考相关文献计算出各物种的重要值(IV),IV=(相对密度+相对高度+相对显著度)/3;物种多样性的测度主要采用Shannon-Wiener指数(H)、Simpson指数(C)、和Pielou均匀度指数(J)等指标,选用目前应用较为普遍的计算公式[13]:
Shannon-Wiener指数:H=-∑PilnPi;
Simpson指数:C=∑(Pi/N)2;
Pielou均匀度指数:J=(-∑PilnPi)/lnS;
式中:Pi为第i种的相对重要值,N为取样中所有种类的重要值总值,S为样地的物种总数。
采用Excel 2010、Sigma Plot14.0软件进行数据处理与分析。
3 结果与分析
3.1 长叶榧树野生种群分布与群落特征
3块样地长叶榧树野生资源的调查结果见表1。
神仙居景区共调查三块样地。第1块样地(编号为A1)内长叶榧树个体数为6丛,共13个分枝,其胸径变化范围为1.0~3.4 cm,树高为1.5~4.0 m,群落内个体生长状态良好。群落主要分为乔木层、灌木层和草本层,乔木层为该群落的主要层,灌木层和草本层为次要层。乔木层的高度为1.5~10.0 m,盖度为70%,优势种为毛竹(Phyllostachysedulis),主要伴生树种还有浙江楠(Phoebechekiangensis)和檵木(Loropetalumchinense)等;灌木丛高度为0.5~1.0 m,以杜茎山(Maesajaponica)为优势种,盖度为20%;草本层优势种主要为野牡丹科植物,盖度约为20%,草本层植株整体较为矮小,在0.5 m以下。第2块样地(编号为A2)位于西南坡,未受人为干扰,受自然干扰较弱,长叶榧树成年植株长势旺盛,生命力强,个体数为4丛,共15个分枝。乔木层高度2~8 m,覆盖度达70%,主要伴生植物为三角枫(Acerbuergerianum)、尾叶樱桃(Cerasusdielsiana)和乌冈栎(Quercusphillyraeoides)等。第3块样地(编号为A3)处于东北坡向,低山地貌,样地内长叶榧树个体数为10丛,共21个分枝,树高为1.8~6.0 m,胸径变化范围为1.4~12.5 cm,生长状况良好,未受人为干扰,受毛竹扩张影响较弱。乔木层最高高度达12 m,覆盖度为60%。主要伴生植物为刨花润楠(Machiluspauhoi)、猴欢喜(Sloaneasinensis)和红柴枝(Meliosmaoldhamii)等。
3.2 刺叶栎野生种群分布与群落特征
3块样地刺叶栎野生资源的调查结果见表1。
上岙村共调查三个样地,均分布于1020 m左右的中山区域,就坡位而言,均处于山脊,群落郁闭度为0.5。第1块样地(编号为A4)内刺叶栎个体数为10丛,共21个分枝,其胸径变化范围为2.0~44.9 cm,树高为2.0~7.8 m,生长状态良好,乔木层为为该群落主要层,盖度为50%,主要伴生植物以黄山松(Pinustaiwanensis)、小果珍珠花(Lyoniaovalifoliavar.elliptica)和白栎(Quercusfabri)为主。第2块样地(编号为A5)生境较为恶劣,内有刺叶栎10丛,共21个分枝,乔木层高度为1.5~12.0 m,覆盖度为50%,主要伴生植物为黄山松、满山红(Rhododendronmariesii)和小果珍珠花等。第3块样地(编号为A6)乔木层最高高度为12.0 m,覆盖度达55%,刺叶栎种群数量较多,主要伴生乔木树种为黄山松、杜鹃(Rhododendronsimsii)和甜槠(Castanopsiseyrei)等。
3.3 重要值分析
根据样地调查数据,统计得出乔木层主要物种的重要值(表2)。可以看出,群落A1-A6的乔木层优势种分别为毛竹、长叶榧树、长叶榧树、刺叶栎、刺叶栎、刺叶栎,重要值在15.28~40.41之间。除群落A1外,其余群落内目标树种数量较多,分布较均匀,相对密度、相对显著度和重要值较大,在群落中起重要作用。
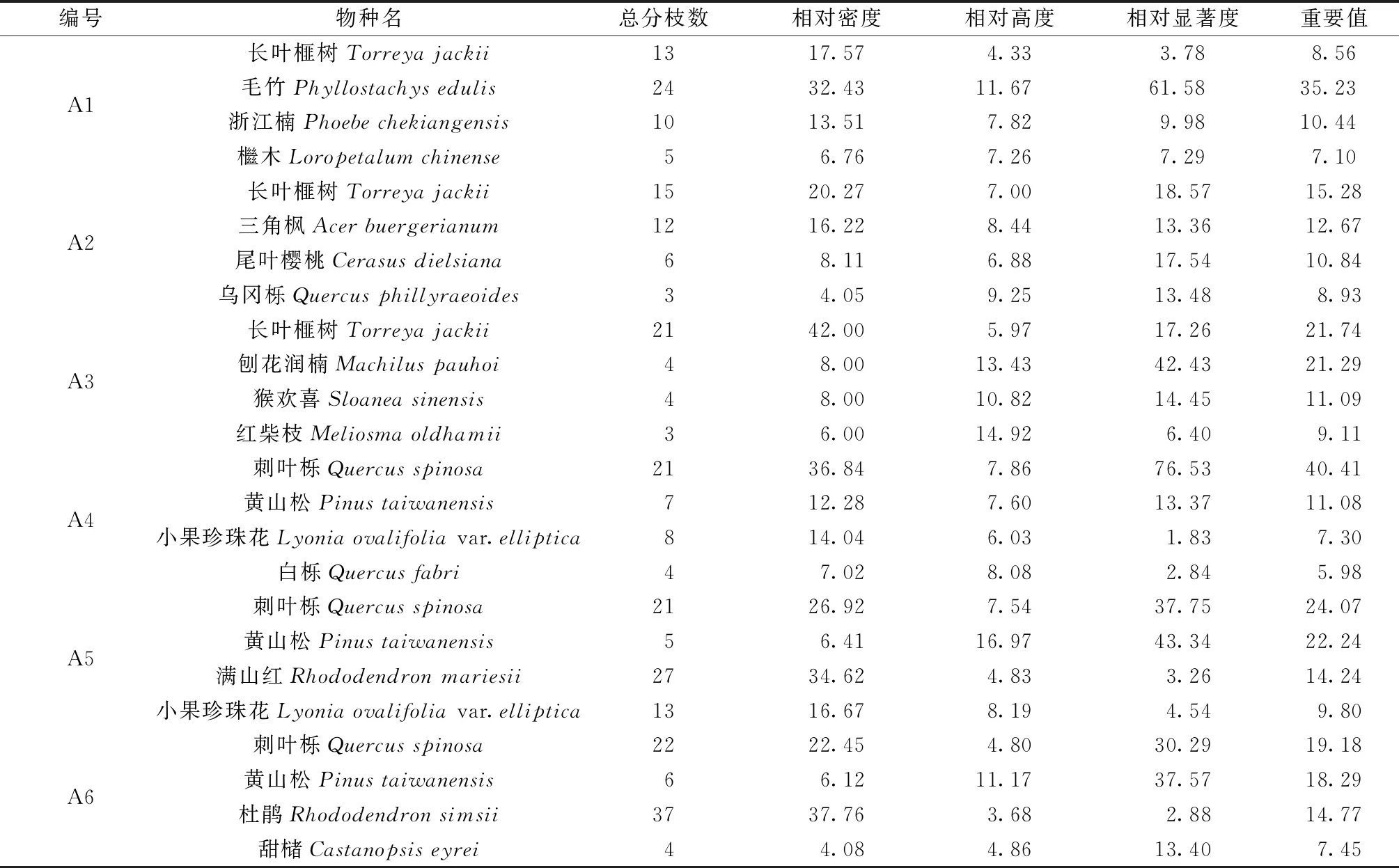
表2 乔木层植物的重要值数据统计表
3.4 径级结构
对六个群落内胸径(DBH)≥1 cm的树种全部测量,按照目标树种与群落内其他树种分别作图(a-f分别代表群落A1-A6,胸径等级1,2,3,4,5,6代表的胸径范围分别是:≤2 cm,2~4 cm,4~6 cm,6~8 cm,8~10 cm,>10 cm)。由图1可以看出,群落内目标树种个体胸径集中分布在1~6 cm间,幼年个体数较多。群落A1(图1-a)中,由于毛竹入侵,长叶榧树种群数量较少,且胸径较小,种群生境较弱。而群落A3(图1-c)中,目标树种长叶榧树幼树个体数量最多,说明该群落内长叶榧树更新能力较强。
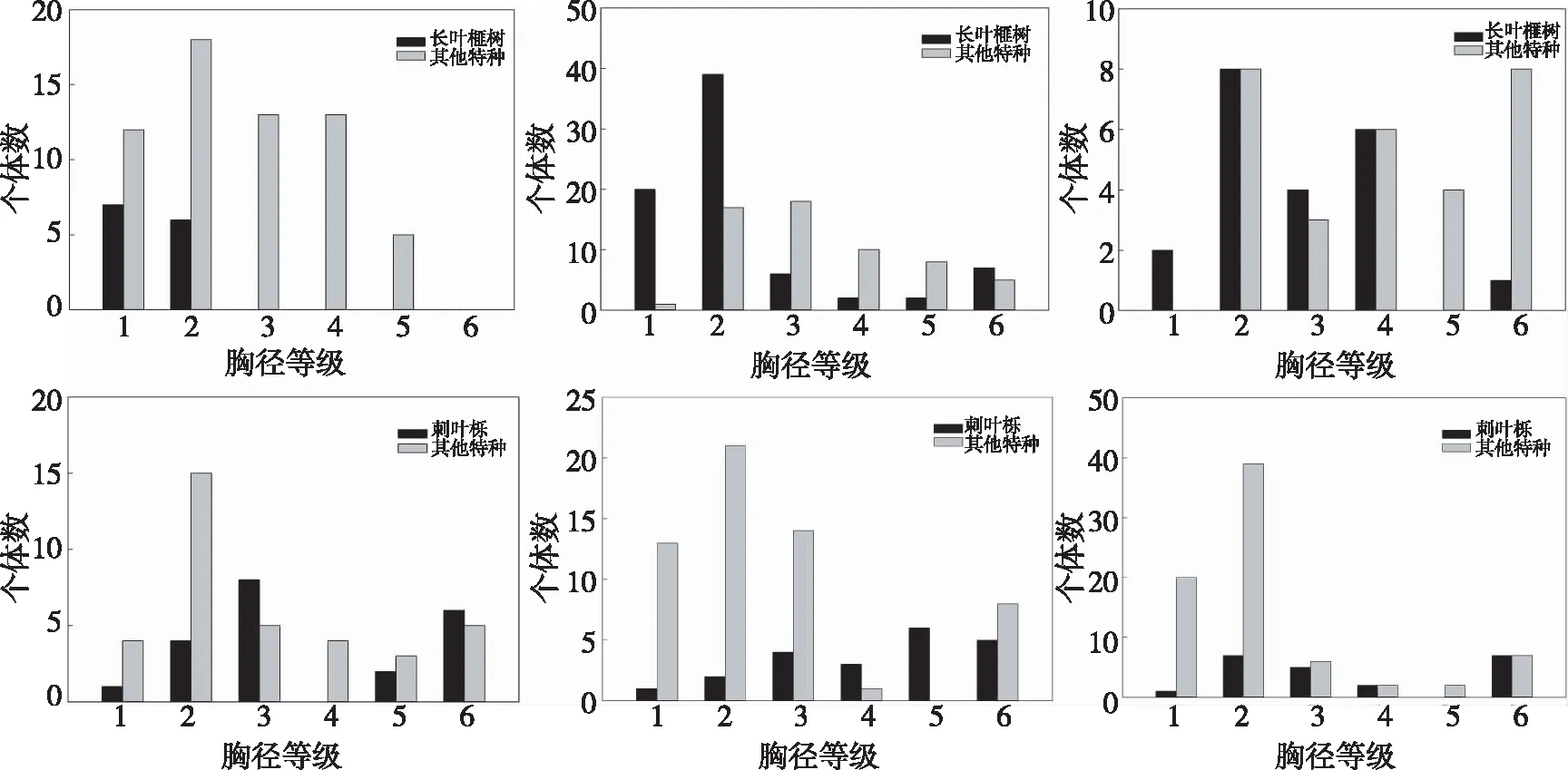
图1 长叶榧树、刺叶栎群落乔木物种的径级结构
3.5 群落物种多样性的分析
物种多样性能有效表征生物群落和生态系统的复杂性[14]。由表3可知,虽然群落A1、A2立地条件、总分枝数、总调查面积都相同,但群落A1受到毛竹扩张干扰严重,各指数都比群落A2的小。说明在相同生境下,受干扰程度越大,多样性指数就越低。从表3还可以看出,长叶榧树在群落A2多样性指数较高,刺叶栎在群落A4、A6多样性指数较高,反应出这些群落生长条件良好。
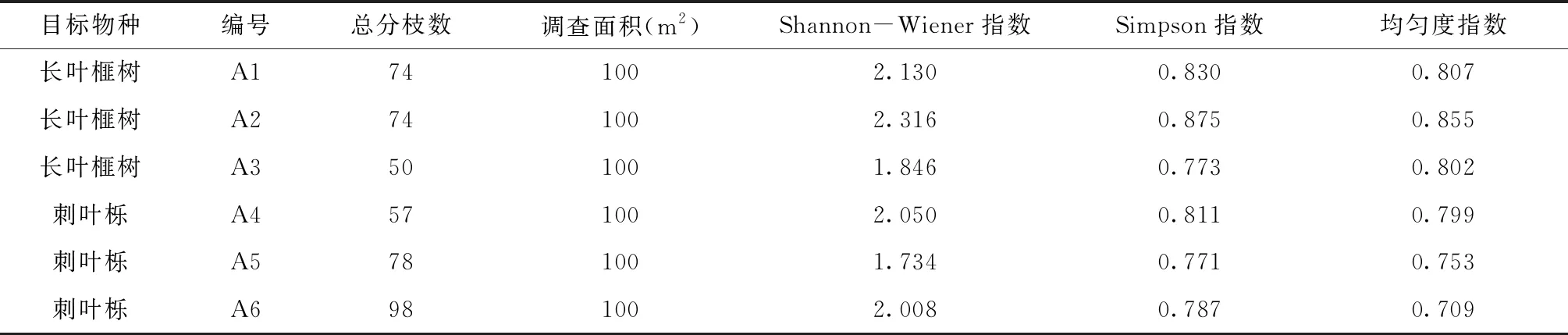
表3 长叶榧树、刺叶栎群落物种多样性
4 讨论
4.1 长叶榧树、刺叶栎的种群结构
调查研究表明,长叶榧树与刺叶栎为六个野外群落中主要的建群种与优势种,其对群落的组成、物种的结构和生态环境的调整起着决定作用。群落A1、A2、A4、A6生境状况良好,受自然及人为干扰影响较小。有关研究结果表明,群落结构越复杂,物种多样性指数就越大[15],这四个群落Shannon-Wiener指数与Simpson指数较大,均匀度指数也相对较高,说明群落结构较为复杂,有较高的稳定性,目前长叶榧树与刺叶栎野外群落生长状况良好。
本研究调查的6个群落中,群落A2、A3长叶榧树胸径结构呈倒“J”型分布,属于稳定增长型结构;群落A1长叶榧树受毛竹扩张影响,种群数量较少;刺叶栎在群落内个体数较少,在各个径级内均有分布,且分布较为均匀。立木胸径的分布可以反映群落的数量特征和年龄结构,是指示植物群落稳定性和生长发育情况的重要指标[16]。长叶榧树作为群落A2、A3的优势种,其幼龄个体数量丰富,个体胸径集中分布在1-6 cm间,这表明长叶榧树已经在群落内形成比较稳定的群落,能够在自然条件下进行稳定的自我更新。
与彭佳龙等人[11]研究刺叶栎在浙江种群的大小结构相比,上岙村刺叶栎群落的胸径分布更集中于中等径级及大径级,由于小径级刺叶栎较少,种群的更新情况较差,种群不能快速增长。当老树死亡后,由于新个体不足,整个刺叶栎种群会产生衰退甚至消失,因此,需要通过人为保护来使整个群落达到平衡状态。
4.2 长叶榧树、刺叶栎的保护价值
种群与群落调查结合文献分析,我们认为:浙江仙居的长叶榧树、刺叶栎种群及其群落具有重要的保护价值。主要体现在:其一,科学研究价值。叶绿体基因组揭示长叶榧树是榧树属最早分化的物种[1],核基因组证明长叶榧树与榧树遗传上混合而形态上分化[17]。刺叶栎是是东喜马拉雅和横断山脉常绿硬叶栎林的主要组成分子[5],在东部地区呈零星不典型状态分布,但却提示常绿硬叶栎林在东部地区的存在;事实上,乌冈栎(Quercusphillyreoides)和尖叶栎(Quercusoxyphylla)等在东部地区形成常绿硬叶栎林已有报道[18-19]。因此,保护长叶榧树、刺叶栎的科学价值不言而喻。其二,生物多样性保护价值。从属的物种分化与地理分布,榧树属好比冷杉属,在中国东南部地区分布有百山祖冷杉(Abiesbeshanzuensis)、梵净山冷杉(Abiesfanjingshanensis)、资源冷杉(Abiesziyuanensis)等多种濒危的冷杉属树种。长叶榧树的生物地理模式和遗传多样性仍需进一步探究。其三,遗传资源保护利用价值。香榧是榧中的优良变异类型经人工嫁树接培育而成的优良品种,但遗传基础窄,在长期栽培过程中导致品种退化,品种改良和创新势在必行[20]。长叶榧树作为近缘种,是重要的种质材料。研究表明长叶榧树与榧树之间存在渐渗杂交[17]。东部地区的刺叶栎与西部地区存在遗传分化,保存有特有的单倍型[6],是重要的遗传资源。其四,资源植物保护利用价值。对于榧树[21]、巴山榧树[22]、云南榧树[23]等榧树属植物的植物化学报道很多,长叶榧树也是一直潜在的植物资源。
4.3 长叶榧树、刺叶栎的受威胁状况与保护对策
过去当地环保部门缺乏对长叶榧树重要价值的认识,当地村民缺乏保护的意识,砍伐较为严重,导致长叶榧树生境丧失,一度有物种灭绝的危险。而刺叶栎自身生长缓慢,材质坚硬,是良好的用材树种,上世纪八、九十年代,当地村民常作为薪材采伐,甚至于作为烧白炭的上等之材,因而使其遭到过度采伐和严重破坏。其二,由于长叶榧树地理分布范围狭窄,其主要生长在山势陡峭峡谷、多基裸露的陡峭阴坡、溪流两旁的常绿阔叶林或次生灌丛中,生境较为恶劣,导致资源数量较少。刺叶栎在浙江省范围内,仅在仙居县淡竹乡有自然分布,分布面积187 hm2,个体数量仅存8100株左右。两者的分布区狭窄、数量少,造成种群近亲交配率提高,繁殖力下降,种群进一步衰弱[24]。过度砍伐、盲目毁林开荒、过度放牧等人类活动导致长叶榧树与刺叶栎野外生境遭到破坏,种群数量锐减,两者受到严重的威胁。
在生物多样性保护实践中经常面临以下问题:生态系统、物种和基因三层次的保护,哪一层次优先?生物和生境何者优先保护?被动就地保护与迁地保护或种群回归能否有机结合?根据我们的实地调查,笔者认为:可以采用以下三个统一:第一,生态系统、物种和基因三层次的统一。根据物种-遗传多样性关联原理,群落内的物种多样性与群体内的遗传多样性受到类似的环境因子的影响,如生境面积、隔离状况、时空异质性等,从而平行的影响迁移、遗传漂变、选择等过程[25]。因此,制定保护措施时,不能将不同层次的生物多样性对立起来,而应维护生态系统、物种和基因多样性保护的统一。第二,生物和生境保护的统一。生境影响生物-生物、生物-环境互作过程,如营养与取食互作、传粉互作、植物-微生物互作等。生境的破坏,如破碎化、山体破坏、水系破坏和重度人为干扰,均会影响生态学过程。仙居县之所以容纳高度的生物多样性,关键在于地形、土壤、气候等因子的高度异质性,形成多样的生态位。例如山区气温空间分布高度异质,不同坡度和坡向间年均气温差最高可达到10~12.5℃[26]。因此,主张生物和生境保护的统一。第三,就地保护与种群回归的统一。就地保护固然重要,必要的迁地保护、利用现代技术扩繁个体、开展种群回归试验是更加积极的保护对策。崖柏和德保苏铁是两个典型案例[27-28]。
此次调查长叶榧树野生种群位于神仙居景区内,离主栈道较远,未受人为干扰影响,自然干扰强度较弱,主要受毛竹扩张及台风暴雨影响,生长状况较为良好,长叶榧树已经在群落内形成比较稳定的群落,能够在自然条件下进行稳定的自我更新。因此,通过建立自然保护区、保护小区或保护点[29],在保护区内开辟巡护线路,加强巡护工作,设立长叶榧树、刺叶栎种群分布区专门巡护线路,定期开展巡护、观测,并记录归档,对种群数量较多的区域进行重点保护,尤其是靠近景区位置,努力恢复群落的自我修复能力。对生长状况较弱的个体采取针对性的人工干预保护措施和开展人工栽培,建立其种质资源库,实施迁地保存,以防止物种遗传多样性和优良遗传基因的丢失[30]。
此外,加强宣传教育工作,突出长叶榧树与刺叶栎在全球生态系统中的重要地位,使当地居民认识其重要性,自觉保护。加强长叶榧树与刺叶栎保护生物学的基础研究工作,通过了解群落的动态演变、种群内的遗传多样性和人工繁殖等技术,为长叶榧树与刺叶栎的保护提供更多的技术指导。
猜你喜欢
吉林林业科技(2023年6期)2023-11-20 02:13:16
滇池(2022年3期)2022-03-05 23:11:38
内蒙古林业调查设计(2021年5期)2022-01-05 02:50:40
电脑迷(2021年12期)2021-11-28 03:47:04
落叶果树(2021年6期)2021-02-12 01:29:10
现代青年·精英版(2018年5期)2018-07-02 22:37:26
南风(2018年22期)2018-01-30 15:55:00
林业勘察设计(2017年4期)2017-07-06 08:46:12
体育科技(2016年2期)2016-02-28 17:06:07
西华师范大学学报(自然科学版)(2015年3期)2015-02-27 15:31:17
