
二化螟P-糖蛋白基因的克隆、序列分析及在阿维菌素处理下的表达模式
2020-09-07 08:24梁建文丁文兵贺华良李有志
江西农业大学学报 2020年3期
张 乾 ,潘 峰,梁建文,邱 林,丁文兵,贺华良,李有志*
(1.湖南农业大学 植物保护学院,湖南 长沙 410128;2.湖南省永顺县植保站,湖南 永顺 416700;3.湖南省赫山区植保站,湖南 赫山 413002)
【研究意义】二化螟(Chilo suppressalis(Walker))属鳞翅目(Lepidoptera)螟蛾科(Pyrlidae),广泛分布于亚洲温带和亚热带稻区,是我国最具破坏性的水稻害虫之一[1]。化学防治是当前防治二化螟的主要手段。阿维菌素是一种大环内酯类化合物,具有强烈的杀虫活性和高效低残留等特点,是防治二化螟的理想药剂[2]。阿维菌素作用于配体(谷氨酸、γ-氨基丁酸、组胺等)门控的氯离子通道,导致神经传递受阻,从而使昆虫产生麻痹、拒食直至死亡[3]。由于该药剂的不合理使用,湖南、浙江等地田间二化螟种群对阿维菌素产生了抗性,湖南祁东等地甚至达到了高水平抗性[4-5]。因此,研究二化螟对阿维菌素产生抗性的机制能够为二化螟防治和阿维菌素类药剂的合理使用提供理论指导。【前人研究进展】害虫对阿维菌素的抗性与多种解毒代谢酶有关,包括细胞色素P450 多功能氧化酶(P450s)和酯酶(CarEs)等第一阶段解毒酶[6-7],谷胱甘肽S-转移酶(GSTs)和尿苷二磷酸-糖基转移酶(UGTs)等二阶段解毒酶[8-9],以及ABC 转运蛋白(ATP-binding cassette transporters)等第三阶段药物转运蛋白[10]。ABC 转运蛋白存在于所有生物中,是由很多蛋白组成的一个蛋白超家族,它们介导无机离子、糖类、氨基酸、脂类、脂多糖、多肽、金属、异源物质和药物的转运[11]。根据ATP 保守结合域(NBDs)的序列相似性,ABC 转运蛋白可分为ABCAABCH 8 个不同的亚家族,其中ABCB、ABCC 和ABCG 亚家族成员普遍与节肢动物的异源解毒有关[12]。【本研究切入点】ABCB亚家族由半转运蛋白(HTs)和全转运蛋白(FTs)组成,其中ABCB全转运蛋白也被称为P-糖蛋白(Pgp)[13],于1976 年在耐秋水仙碱的中国仓鼠卵巢细胞中首次发现[14],P-糖蛋白基因的过表达可介导多药耐药性(MDR)。生物有机体通常具有多个P-糖蛋白基因,昆虫的P-糖蛋白基因首先在果蝇中克隆并鉴定,果蝇至少有3 个P-糖蛋白基因,根据染色体位置分别命名为mdr49,mdr50 和mdr65[15]。目前,许多昆虫的P-糖蛋白基因已经被克隆和鉴定,该基因的过量表达与多种昆虫对不同种类杀虫剂的耐受性或抗性有关[13]。在鳞翅目昆虫中,P-糖蛋白与小菜蛾对Cry1Ac的抗性有关[16],与甜菜夜蛾[10]和小菜蛾[17]对阿维菌素抗性有关。【拟解决的关键问题】本研究试图探明二化螟P-糖蛋白与阿维菌素代谢和抗性之间的关系,为此克隆了该虫两个P-糖蛋白基因并分析其序列,进而研究了P-糖蛋白基因在阿维菌素诱导下的表达模式。
1 材料与方法
1.1 供试昆虫
敏感种群由南京农业大学高聪芬教授惠赠;田间种群于2019年采自湖南农业科学院院内稻田,饲养一代后进行生物测定。养虫室温度(28±1)℃,光周期16L∶8D,相对湿度75%,饲料为人工饲料。
1.2 主要试剂
阿维菌素95%原药购自浙江钱江生物化学股份有限公司;RNAiso Plus、pMD18-T Vector Cloning Kit、DH5α感受态细胞、PrimeScript II 1st Strand cDNA Synthesis Kit、PrimeScript RT reagent Kit with gDNA Eraser、TB Green Premix ExTaqII 购自北京宝日医生物技术有限公司;Axygen®AxyPrep DNA Gel Extraction Kit购自美国康宁公司;DEPC处理水购自生工生物工程(上海)股份有限公司。
1.3 方法
1.3.1 生物测定 使用人工饲料混毒法测定阿维菌素对敏感种群的毒力回归曲线[18]。以丙酮为溶剂,将原药配制成高浓度母液,然后用清水稀释成不同浓度的药液处理(含体积分数为1%的吐温80)。将5 mL的阿维菌素药液加入到45 mL 人工饲料中(40~50 ℃),混匀、凝固后,接入4 龄幼虫。设溶剂对照和空白对照,每处理重复4次。计算处理72 h后的LC10、LC30和LC50值。
使用毛细管点滴法(水稻二化螟抗药性监测技术规程NY/T 2058-2014)测定阿维菌素对田间种群的毒力回归曲线,以丙酮为溶剂,将原药配制成一定浓度的母液,按倍半稀释法稀释成不同浓度的药液处理(含体积分数为1%的吐温80)。将药液点滴于4 龄幼虫前胸背板上,设空白对照和溶剂对照,每个处理重复4次。计算处理72 h后的LD50值,抗性倍数=田间种群的LD50值/敏感种群的LD50值,二化螟敏感种群对阿维菌素的LD50值是0.000 17µg/虫[5]。
1.3.2 二化螟总RNA 的提取和cDNA 第一链的合成 参照RNAiso Plus 说明书,单头提取敏感品系4 龄幼虫总RNA,使用NanoDrop 1 000 超微量分光光度计测定RNA 的浓度和纯度,置于-80 ℃超低温冰箱保存。使用PrimeScript II 1st Strand cDNA Synthesis Kit 试剂盒进行cDNA 第一链的合成,取1µg 提取的总RNA作为模板,反应结束后于-20 ℃冰箱保存。
1.3.3 二化螟P-糖蛋白基因的克隆 结合二化螟转录组数据信息,获得二化螟P-糖蛋白基因片段。利用Primer Premier 5.0软件分别设计了3对特异性引物(表1)。以合成的cDNA 第一链为模版进行PCR 扩增,PCR 扩增体系为20 µL,包括10 µL PremixTaq,1 µL cDNA,1 µL 上游引物,1 µL 下游引物和7 µL ddH2O。扩增条件:98°C 2 min;98 ℃10 s,53 ℃30 s,72 ℃2 min,30个循环;72 ℃延伸10 min。对PCR产物进行电泳检测,将符合预期大小的DNA片段进行纯化回收。
纯化PCR 产物连接到pMD18-T 载体上,再转化到DH5α感受态细胞,之后在LB 培养基上(含氨苄)37 ℃过夜培养,使用M13通用引物检测连接情况,连接成功的菌液委托生工生物工程(上海)股份有限公司进行双向测序。
1.3.4 二化螟P-糖蛋白序列分析 测序完成后,利用DNAMAN9.0 软件进行序列拼接,获得二化螟CsPgp1 和CsPgp2 基因cDNA 序列,使用ORF Finder 预测开放阅读框(https://www.ncbi.nlm.nih.gov/orffinder),使用ExPASY Pro-teomic Server 进行蛋白分子量和等电点预测(http://web.expasy.org/compute_pi),使用NCBI CDD(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)和SMART(http://smart.embl-heidelberg.de)进行蛋白结构域分析,使用DNAMAN9.0 软件对二化螟P-糖蛋白氨基酸序列和其他物种的同源序列进行比对,用MEGA7.0软件基于邻接法(Neighbor-joining,NJ)构建系统发育树(进行1 000次重复分析)。
1.3.5 阿维菌素对二化螟P-糖蛋白基因表达的影响 (1)不同剂量阿维菌素处理二化螟幼虫。为了检测在阿维菌素处理下,二化螟CsPgp1和CsPgp2基因在转录水平的反应模式,根据毒力测定结果,用阿维菌素对敏感种群四龄幼虫72 h的LC10、LC30和LC50值进行处理,以丙酮处理作为空白对照(CK),处理方法同本文中的生物测定方法,每个处理有4个生物学重复,分别在处理后24 h和48 h挑取活虫液氮速冻,置于-80 ℃保存。
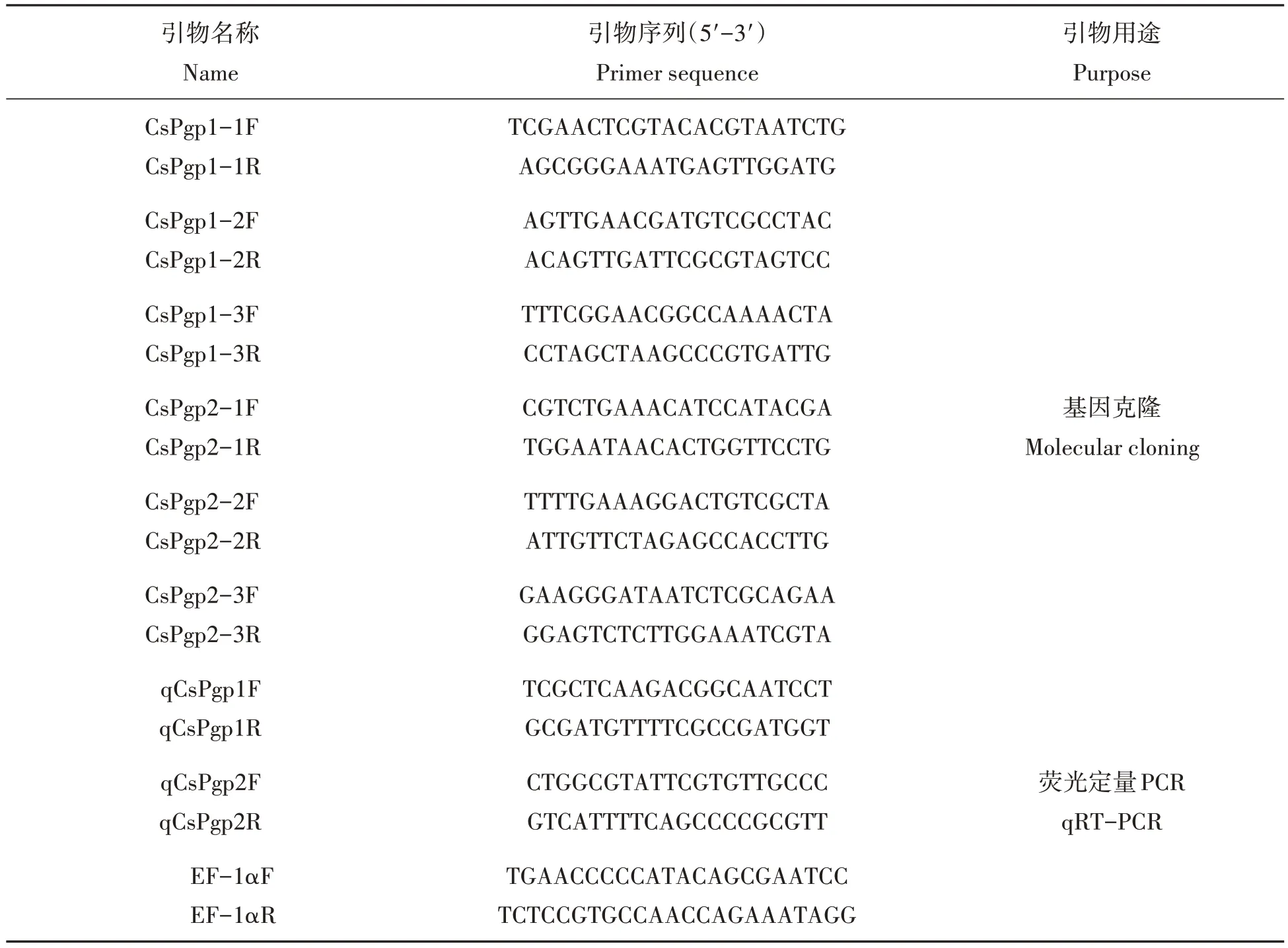
表1 研究所用引物Tab.1 Primers used in this study
(2)不同种群幼虫样品收集。分别取敏感种群和田间种群四龄二化螟幼虫样品,每个处理4 个生物学重复,置于-80 ℃保存。
(3)总RNA的提取和cDNA的合成。分别提取不同处理的幼虫总RNA,测定RNA浓度和纯度后置于-80 ℃保存。使用PrimeScript RT reagent Kit with gDNA Eraser试剂盒合成二化螟cDNA,cDNA于-20 ℃保存。
(4)实时定量分析。根据克隆获得的两个P-糖蛋白基因cDNA 序列,使用Primer-BLAST 分别设计qPCR 引物,以EF-1α作为内参基因[19],引物信息见表1。PCR 扩增体系为10µL,包括5µL 2×TB Green Premix ExTaqII,1µL cDNA,0.4µL 上游引物,0.4µL 下游引物和3.2µL ddH2O,在CFX96 Touch 荧光定量PCR仪选用两步法程序进行PCR扩增。
1.4 数据分析
使用Bio-Rad CFX Maestro 1.1 软件分析qRT-PCR 数据,采用2-△△Ct法计算P-糖蛋白基因的相对表达量,不同处理和不同种群P-糖蛋白基因的表达量使用SPSS 24.0 统计软件进行单因素方差分析(oneway ANOVA)(P<0.05),使用GraphPad Prism 8软件作图。
2 结果与分析
2.1 阿维菌素对二化螟毒力
阿维菌素对敏感种群72 h 的LC10、LC30和LC50值分别为0.004,0.010,0.017 mg/L,阿维菌素对田间种群72 h 的LD50值为0.010 54 µg/虫,抗性倍数为62 倍,为中等抗性水平,在随后的实验中,分别用LC10、LC30和LC50浓度的阿维菌素处理二化螟敏感种群4龄幼虫。
2.2 二化螟P-糖蛋白基因cDNA的克隆和序列分析
克隆获得二化螟CsPgp1 和CsPgp2 基因的cDNA 序列,开放阅读框全长分别为3 780 bp 和3 939 bp,分别编码1 259 和1 312 个氨基酸(图1),预测分子量分别为137 ku 和144 ku,等电点分别为6.58 和8.52。使用NCBI CDD 和SMART 分析发现CsPgp1 和CsPgp2 均由两个相似的部分构成,每个部分包含一个跨膜结构域(transmembrane domain,TMD)和一个核苷酸结合结构域(nucleotide-binding domain,NBD),从蛋白N 端到C 端形成了TMD-NBD-TMD-NBD 结构,这是完整的ABC 转运蛋白经典结构(图1、图2)。CsPgp1 的跨膜结构域包含12 个跨膜螺旋区域(TM),CsPgp2 的跨膜结构域包含11 个TM;每个核苷酸结合结构域都携带有ATP 结合位点,ABC 典型结构,Walker A,Walker B,D 环,Q 环和H 环等标签序列(图2、图3)。
综上所述,文化因素、社会因素、个人因素与少数民族大学生创业能力均呈现显著正相关关系,对其创业能力的提升都起着重要作用,这为少数民族大学生创业能力的有效提升提供了参考价值。

图1 二化螟CsPgp1(A)和CsPgp2(B)的核苷酸序列及预测的氨基酸序列Fig.1 Nucleotide sequence and deduced amino acid sequence of CsPgp1 and CsPgp2 from Chilo suppressalis
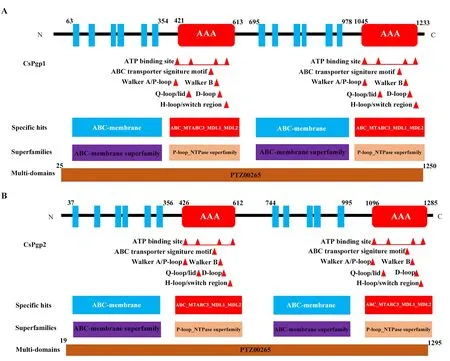
图2 CsPgp1(A)和CsPgp2(B)蛋白的一级和二级结构示意图Fig.2 Schematic drawing of the primary and secondary structures of CsPgp1(A)and CsPgp2(B)
2.3 二化螟P-糖蛋白的系统发育分析
利用CsPgp1和CsPgp2的氨基酸序列和其他物种的同源序列进行比对分析。结果表明,P-糖蛋白与其他昆虫的多药耐药性蛋白有很高的相似性,其中CsPgp1 与亚洲玉米螟Ostrinia furnacalisMDR1(XP_028158084.1)的关系最近,一致性为77%,与小菜蛾Plutella xylostellaPgp(QDY92632.1)、家蚕Bombyx moriMDR1(XP_021202653.1)和黑腹果蝇Drosophila melanogasterMDR50(NP_523740.3)氨基酸序列一致性分别为73%、72%和44%。CsPgp2 与亚洲玉米螟O.furnacalisMDR49-like(XP_028159924.1)的关系最近,一致性为81%,与粉纹夜蛾Trichoplusia niMDR3(ADV76539.1)、家蚕B.moriMDR49(XP_004929922.1)和黑腹果蝇D.melanogasterMDR49(AAA28679.1)氨基酸序列一致性分别为78%、76%和43%,CsPgp1和CsPgp2分别对应黑腹果蝇的mdr50和mdr49两个P-糖蛋白。
使用MEGA7.0 软件构建系统发育树(图4),结果显示,二化螟CsPgp1 和CsPgp2 聚为不同的两支,表明这两个基因可能具有不同的进化模式,但二者都和鳞翅目昆虫的P-糖蛋白(或MDR)聚为一支,说明二化螟CsPgp1和CsPgp2具有高度的进化保守性。此外,还可以看出二化螟与亚洲玉米螟的亲缘关系最近,其他目的昆虫各自聚成小类群也说明了P-糖蛋白在进化上具有很高的保守性。

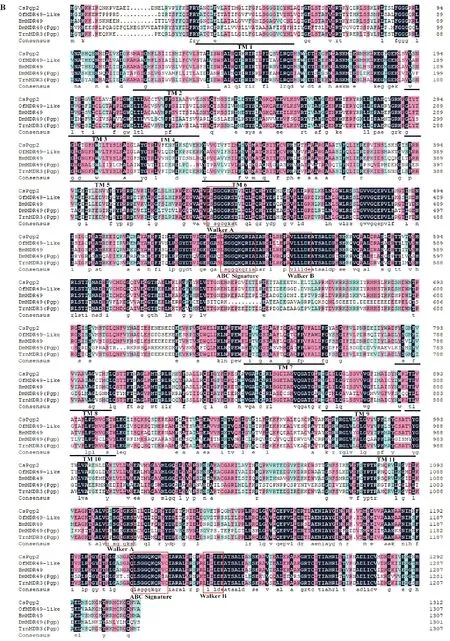
图3 CsPgp1(A)和CsPgp2(B)氨基酸序列与其他物种比对Fig.3 Comparison of the deduced amino acid sequences of CsPgp1(A)and CsPgp2(B)subunits with other species
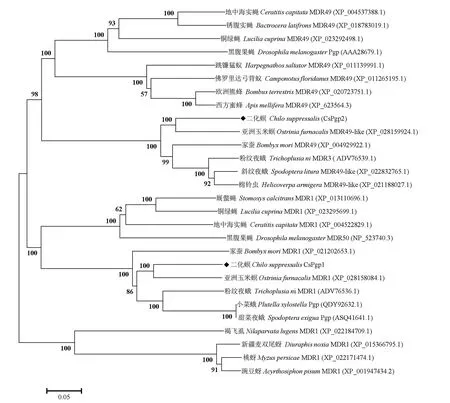
图4 二化螟CsPgp1和CsPgp2与其他物种P-糖蛋白的系统发育分析Fig.4 Molecular phylogenetic analysis of CsPgp1 and CsPgp2 along with other organism Pgps
2.4 阿维菌素处理下二化螟P-糖蛋白基因的表达模式
2.4.1 阿维菌素临时刺激对二化螟P-糖蛋白基因表达量的影响 不同剂量阿维菌素处理二化螟4龄幼虫后不同时间点CsPgp1 的转录水平如图5A 所示,与对照组相比,处理24 h,各处理组的CsPgp1 表达量显著上调(P<0.05),分别为对照组的1.97 倍、2.18 倍、1.87 倍,LC30处理组的CsPgp1 表达量显著高于LC50处理组(P<0.05);处理48 h,LC10处理组的CsPgp1 表达量显著高于对照组(P<0.05),为对照组的1.28 倍,LC30和LC50处理组的CsPgp1表达量与对照组差异不显著。
不同剂量阿维菌素处理二化螟4龄幼虫后不同时间点CsPgp2的转录水平如图5B 所示,与对照组相比,处理24 h,各处理组的CsPgp2 表达量显著上调(P<0.05),分别为对照组的3.23 倍、1.87 倍、2.12 倍,LC10处理组的CsPgp2 表达量显著高于LC30和LC50处理组(P<0.05);处理48 h,各处理组的CsPgp2 表达量与对照组差异不显著。
2.4.2 二化螟P-糖蛋白在不同种群中mRNA 相对表达量的差异 敏感和田间种群的P-糖蛋白相对表达量如图6所示,田间种群的CsPgp1和CsPgp2表达量均显著高于敏感种群(P<0.05),分别是敏感种群的3.01倍和2.39倍。
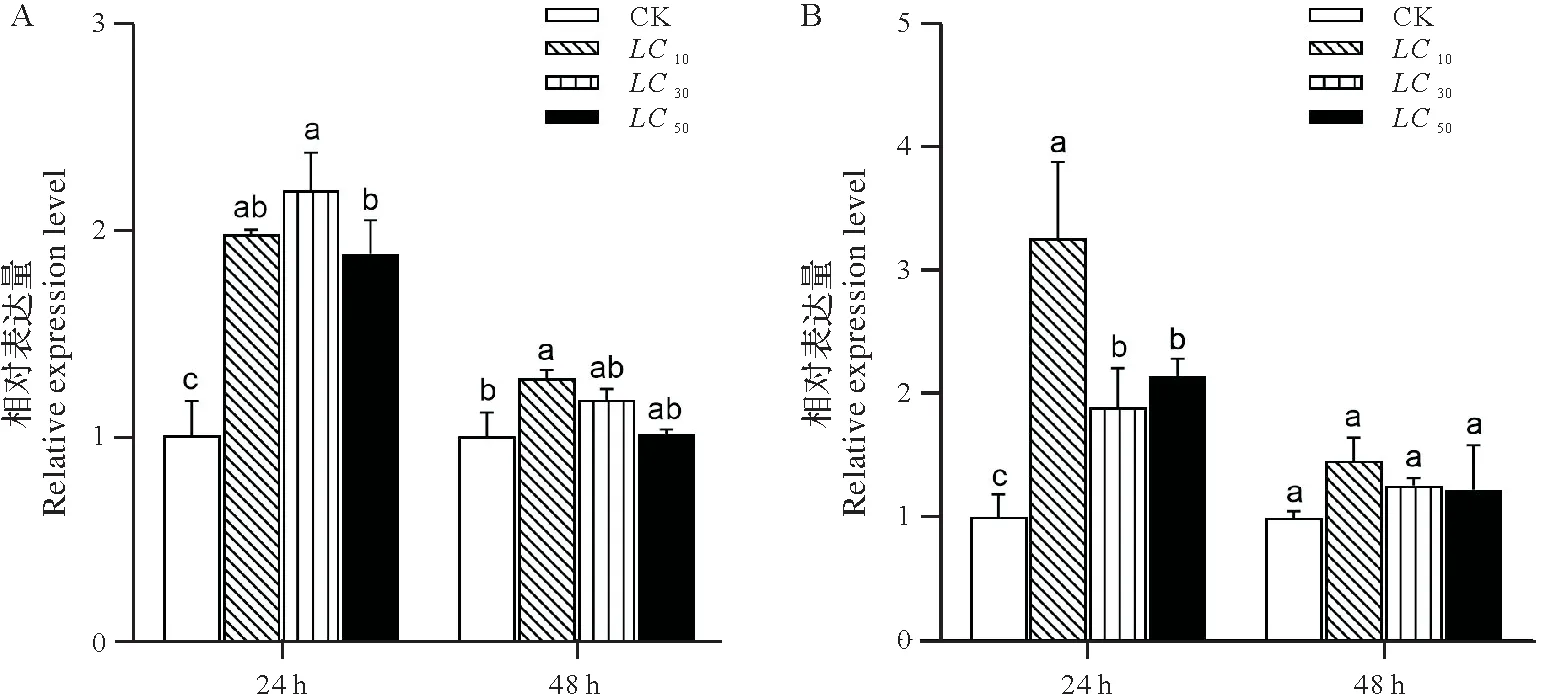
图5 阿维菌素处理下二化螟4龄幼虫CsPgp1(A)和CsPgp2(B)的相对表达量Fig.5 Relative expression level of CsPgp1(A)and CsPgp2(B)mRNA in 4th instar larval of Chilo suppressalis treated with abamectin
3 结论与讨论
害虫对杀虫剂的抗性是由不同的机制引起的,包括行为的改变,降低杀虫剂对表皮的穿透性,杀虫剂作用靶标位点的突变和解毒代谢酶介导的代谢抗性等[6-9]。ABC 转运蛋白是一类第三级药物转运蛋白,已有报道[12]显示其与27 种不同的杀虫剂(杀螨剂)的转运或抗性有关。P-糖蛋白是ABC 转运蛋白超家族的成员,具有非特异性底物结合位点,从而使昆虫对杀虫剂等产生抗性或耐受性[13]。近年来,阿维菌素广泛用于我国水稻种植区进行二化螟的防治,通过对二化螟抗药性的持续监测,2015 年湖南祁东等地田间种群已经对阿维菌素产生了高水平抗性[4]。现已在多种昆虫中证实P-糖蛋白介导了对阿维菌素和Bt蛋白的抗性[3,10,16-17],因此,研究P-糖蛋白与二化螟对阿维菌素抗性之间的关系具有重要的意义。
阿维菌素属于大环内酯的阿维菌素亚家族,已被广泛用于鳞翅目害虫的防治。目前,关于二化螟对阿维菌素抗性的研究主要在阿维菌素的受体基因,研究表明RNA 干扰二化螟GABA 受体基因的两个亚基CsRDL1和CsRDL2,幼虫对阿维菌素的敏感性显著降低[19];二化螟GluCl基因有3个可变剪接体(CsGlu-Cl A、CsGluCl B和CsGluCl C),RNA 干扰结果表明沉默该基因能够显著增加幼虫对阿维菌素的敏感性[21]。本研究中在不同浓度阿维菌素的临时刺激下,二化螟CsPgp1 和CsPgp2 的表达量在24 h 显著上调,表明这两个基因对阿维菌素均存在应激反应,可能是因为这两个基因均参与了阿维菌素在二化螟体内的代谢。两项相似的研究表明小菜蛾的1个P-糖蛋白和朱砂叶螨的两个P-糖蛋白基因同样能够在阿维菌素的诱导下表达上调[3,17]。与24 h相比,各处理组的CsPgp1和CsPgp2在48 h表达下降,表明二化螟P-糖蛋白在阿维菌素的诱导下表现出时间效应,诱导的P-糖蛋白高表达可能参与阿维菌素的代谢过程。处理24 h,LC30处理组的CsPgp1 表达量显著高于LC50处理组,LC10处理组的CsPgp2 表达量显著高于另外两种浓度处理,表明对于不同浓度的阿维菌素,P-糖蛋白的耐受水平也会不一样,在棉铃虫中也存在类似的结果[22]。
本研究检测了二化螟敏感种群和田间抗性种群(62 倍)P-糖蛋白基因的表达差异,结果显示田间抗性种群的CsPgp1 和CsPgp2 的表达量均显著高于敏感种群。许多昆虫的P-糖蛋白基因在抗性种群中高表达,如阿维菌素抗性的小菜蛾P-糖蛋白基因表达量显著高于敏感种群[17]。所以P-糖蛋白可能参与了二化螟对阿维菌素的代谢,从而产生抗性或耐受性。此外,本研究中采用的二化螟抗性种群为田间种群,田间种群用药历史复杂,抗性发展机制会有所不同。因此只能间接反映阿维菌素抗性水平高低与P-糖蛋白基因的表达量存在一定的联系,在二斑叶螨的研究中也有类似报道[2]。
综上所述,本研究发现P-糖蛋白基因在二化螟田间抗性种群和临时阿维菌素处理的敏感种群中均表达上调,推测其可能参与了二化螟对阿维菌素的代谢,可能与二化螟对阿维菌素抗性有关。这些结果为研究二化螟对阿维菌素抗性与P-糖蛋白之间的关系提供了参考。近年来,CRISPR/Cas9 基因编辑技术已经成为研究昆虫体内基因功能的有力工具[23],Guo等[24]通过敲除小菜蛾ABCC2和ABCC3基因验证了其在小菜蛾对Cry1Ac 抗性中的作用。下一步可以通过CRISPR/Cas9 技术对P-糖蛋白的功能进行验证,进一步研究P-糖蛋白与二化螟抗药性之间的关系。
猜你喜欢
特种经济动植物(2022年11期)2022-11-15
安徽农学通报(2022年6期)2022-04-07
生物灾害科学(2021年1期)2021-04-09
湖北植保(2020年5期)2020-10-20
农药科学与管理(2019年6期)2019-11-23
农家之友(2019年5期)2019-01-17
中成药(2018年7期)2018-08-04
中成药(2018年1期)2018-02-02
吉林农业(2017年15期)2017-08-29
医学研究杂志(2015年3期)2015-06-10
