
家蝇对氯菊酯抗性的分子机制研究*
2020-09-02 12:41:56邱星辉
寄生虫与医学昆虫学报 2020年2期
刘 艳 李 梅 邱星辉**
(1.中国科学院动物研究所农业虫害鼠害综合治理研究国家重点实验室, 北京 100101;2.中国科学院大学,北京100049)
家蝇是一种重要的病媒生物,可以传播上百种人畜疾病,如霍乱、痢疾、肠出血性大肠杆菌、禽流感病毒以及导致死亡的抗生素抗性细菌等(Scottetal., 2014)。家蝇繁殖能力非常强,单雌产卵量可达千粒,在适宜的温湿度条件下每10~14 d就可以完成一个世代,如不加控制,将带来非常严重的公共卫生灾害。家蝇的控制长期以来主要依靠化学杀虫剂,其中拟除虫菊酯类杀虫剂由于高效、对哺乳动物毒性低以及在环境中易分解等特点被广泛应用。由于拟除虫菊酯类杀虫剂的大量使用,家蝇种群普遍对这类药剂产生了抗药性(Gaoetal., 2012; Wangetal., 2012;Scottetal., 2013)。抗药性的产生将导致家蝇化学防治的困难甚至失败,因此开展抗药性的监测和抗性机制的研究对于杀虫剂的选用具有重要的意义。
家蝇对拟除虫菊酯的抗性主要有两大类机制,即细胞色素P450介导的代谢解毒作用增强以及靶标(钠离子通道)不敏感性(Scottetal., 2017)。拟除虫菊酯抗性家蝇的钠离子通道通常存在1014位点的氨基酸替换,即由野生型(敏感型)的亮氨酸突变为苯丙氨酸(L1014F)或组氨酸(L1014H)。随着杀虫剂选择压的增加,家蝇可能进化产生多位点突变的抗性等位基因,如1014和918双位点突变(M918T+L1014F)。近年来,在美国采集的家蝇中发现了2个抗性相关的新突变即D600N 和T929I,并鉴定了两个新的抗性等位基因,即D600N+M918T+L1014F三位点突变型和T929I+L1014F双位点突变型,该两等位基因相对于以往发现的M918T+L1014F和L1014F抗性等位基因可以导致水平更高、更广谱的抗药性(Kasaietal.,2017;Sunetal., 2017)。
家蝇基因组中有146个P450基因(Scottetal., 2014),文献报道的在某些拟除虫菊酯类抗性品系中超量表达的细胞色素P450基因包括CYP6D1v1、CYP6D3、CYP6D8、CYP6A5v1、CYP6A5v2、CYP6A36、CYP6A40、CYP6G4(Zhuetal., 2008a; Zhuetal., 2008b; Gaoetal., 2012;Scottetal.,2017),其中CYP6D1v1被证明可以代谢解毒菊酯类杀虫剂(Wheelocketal., 1992)。由于P450介导抗性存在进化可塑性,不同家蝇种群导致抗性的P450种类可能不同(Scottetal., 2004),加上P450种类多样,且是膜结合蛋白,其功能鉴定在技术上困难很大,至今人们对P450介导的代谢抗性机制的认识还非常有限(Scottetal.,2017)。
我们在实验室经溴氰菊酯汰选出的高抗性家蝇品系中发现钠离子通道M918T和L1014F突变的存在(Qiuetal., 2007),而在我国家蝇的野生种群中未检测到钠离子通道M918T突变,却发现L1014H是在我国家蝇中广泛分布的抗性突变类型(Wangetal., 2012)。细胞色素P450抗性等位基因CYP6D1v1 在我国野生家蝇种群中也存在(Wangetal., 2012), 但其单独存在时对拟除虫菊酯抗性的贡献不大(Panetal., 2018)。为更全面深入地揭示家蝇对拟除虫菊酯抗性的分子基础,本文分析了从山东济南采集的家蝇对常用杀虫剂氯菊酯的抗性状况与抗性机制。
1 材料与方法
1.1 家蝇品系
本研究涉及到4个家蝇品系,WHO敏感品系从丹麦Michael Kristensen博士实验室引进。TJS品系是实验室长期保存的杀虫剂敏感品系,自1982年采集在实验室正常饲养,未接触过杀虫剂。济南品系(JN)是2009年夏天从山东济南的一个垃圾场采集的野外种群,济南-Sel(JN-Sel)是从JN品系经用氯菊酯连续汰选(死亡率平均在60%左右)7代后抗性水平更高的品系。
1.2 主要试剂和仪器
总RNA采用Trizol试剂提取。用于定量PCR的cDNA合成采用PrimeScriptTM RT reagent Kit with gDNA Eraser试剂盒(TaKaRa),荧光定量PCR采用SuperReal PreMix Plus(SYBR Green)试剂盒(天根公司)。其他主要试剂包括Hpy188Ⅲ、MlucI、EcoRI限制性内切酶(NEB公司),rTaq聚合酶、Primer STAR高保真聚合酶(TaKaRa),RNaseA(天根公司)。氯菊酯(95%)由中国疾病预防控制中心孟凤霞研究员惠赠。荧光定量PCR采用Mx3000P qPCR和3005P qPCR系统(Stratagene), 使用的PCR扩增仪为T 100TM Thermal Cycler PCR扩增仪(Bio-Rad)。
1.3 家蝇品系汰选方法
对羽化后2 d的成虫,根据预实验的毒力测定结果,配制杀死种群约60%个体的剂量,采用点滴法进行处理,24 h后,将存活的家蝇转移到干净的养虫笼中继续传代饲养,保证每次用于传代的家蝇总数在100头以上。
1.4 家蝇对氯菊酯敏感性测定
药剂浓度现用现配,由母液用丙酮稀释,设立5个浓度。选取羽化后第3~5 d大小一致的雌虫,体重在18 mg左右。药液滴在雌蝇中胸背板中央,每头滴1 μL,低浓度向高浓度点滴,每一个浓度点滴30只,每个浓度重复3次,以不含药的丙酮做对照。药剂处理过的家蝇置塑料杯中放在人工气候箱中正常饲养,设置温度为25 ℃,相对湿度为65%。24 h后检查死亡率,用软件Poloplus计算死亡率机率-剂量回归方程的斜率值(b)、致死中剂量(LD50)及其95%的置信区间。
1.5 核酸提取与cDNA合成
单头家蝇基因组DNA的提取采用文献(Rinkevichetal.,2006)的方法,总RNA提取以及cDNA合成按试剂盒说明书进行。
1.6 钠离子通道1014位点基因分型
采用Qiu等(2012)建立的PCR-RFLP方法,通过综合分析两个不同的PCR反应和对应的两个不同限制性内切酶的酶切反应(反应A,反应B)产物的凝胶电泳图谱来判断家蝇样品的基因型。反应A产物的电泳图如果显示156 bp一条带,基因型为LL、HH或HL;如显示3条带(59、96和156 bp),基因型为FL或HF;如显示两条带(59和96 bp)则为FF。反应B电泳图如显示为220 bp的一条带,该样品基因型为LL、FF或FL;如显示三条带(50、170和220 bp)为HL或HF型;如显示两条带(50和170 bp)则为 HH型。
1.7 P450基因表达量的实时荧光定量检测
采用反转录实时荧光定量法(RT-qPCR)检测4个品系8个P450基因(目标基因)的表达量, 以 Actin和RPS3为内参基因。每个品系做3个生物学重复,每个生物学重复采用10头家蝇腹部提取RNA。目标基因与内参基因的PCR引物的核苷酸序列见表1。反应体系为:2×SuperReal PreMix Plus 10 μL, 正反向引物各0.6 μL(终浓度0.3 μmol/L), cDNA 4 μL(80 ng), 50×ROX Reference Dye 0.4 μL,用无RNase酶的水补齐至20 μL。PCR条件为95 ℃ 15 min预变性,40循环95 ℃ 10 s,60 ℃ 32 s,然后95 ℃ 1 min,55 ℃,30 s,接着逐步升温到95 ℃,在升温过程中记录荧光信号值。仪器记录各个样品得到的Ct值、产物熔解曲线。每个样品设3个技术重复,用ddH2O代替cDNA模板作为空白对照。目标基因相对内参基因的表达量(即相对表达量)采用2-ΔCt方法计算,品系之间的目标基因相对表达量值采用单因素方差分析(ANOVA)和 LSD方法进行多重比较, 用不同字母来表示品系间的差异有统计意义(P<0.05)。

表1 荧光定量 PCR引物
2 结果
2.1 不同品系家蝇对氯菊酯的敏感性
4个检测的家蝇品系对氯菊酯的敏感性见表2。从表2可以看到WHO 与TJS品系没有显著差别,而济南品系相对WHO 品系表现出近50倍的抗性。经用氯菊酯汰选7代后的JN-Sel品系获得了比JN更高水平的抗性,相对WHO品系抗性倍数超过100倍。

表2 四个家蝇品系对氯菊酯敏感性
2.2 钠离子通道1014位点基因型检测
为查明抗性品系是否存在1014位点突变介导的击倒抗性机制,采用RFLP的方法检测钠离子通道1014位点基因型。在检测的济南(48头)和济南_Sel品系(63头)家蝇的RFLP图谱可以看到,酶切反应A(图1-A)的条带为156 bp, 酶切反应B(图1-B)的条带220 bp,表明这些家蝇钠离子通道的基因型为1014LL 纯合型。
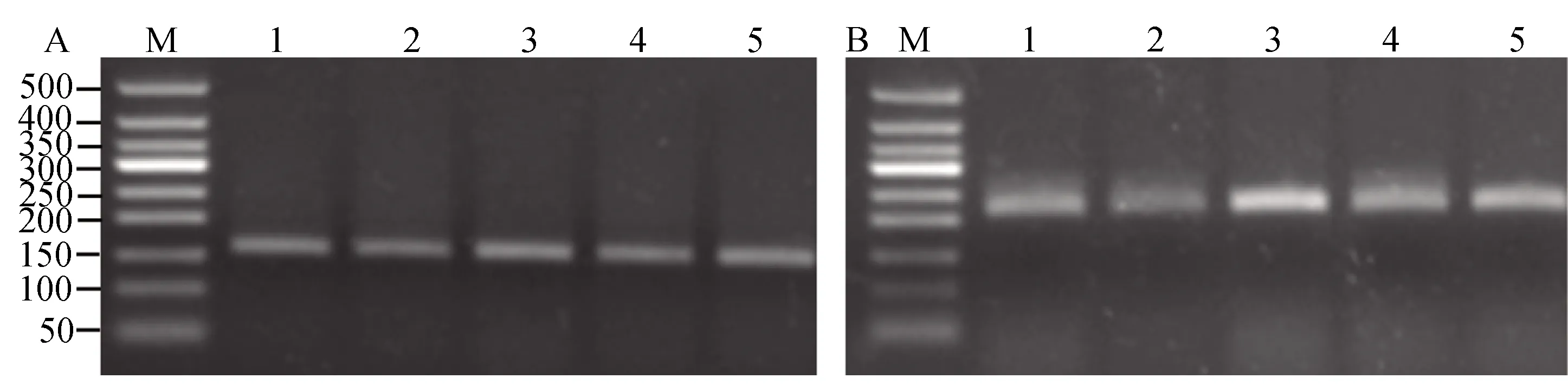
图1 家蝇钠离子通道基因1014位点基因型检测代表性结果
2.3 不同品系家蝇8个细胞色素P450基因的表达分析
用Real-time PCR方法,对4个品系中的可能与拟除虫菊酯类杀虫剂抗性相关的8个P450基因的表达量进行检测(图2)。图2显示,不同基因在不同品系家蝇中的表达是有差异的,整体来看,CYP6G4 和CYP6D1的表达丰度较高,而CYP6A36、CYP6A40和 CYP6D3的表达丰度低;CYP6A40在WHO品系中有更高的表达,CYP6A5v1、CYP6A5v2和CYP6D1在JN抗性品系、CYP6D8和CYP6G4在高抗性JN-Sel品系中表达量最高。相对于两个敏感品系,在两抗性品系中都过量表达的P450基因有CYP6A5v2、CYP6A36和CYP6G4。两个遗传背景相同的抗性品系比较,CYP6D8和CYP6G4在JN-Sel品系中表达量更高,但JN品系CYP6D8的表达量与TJS敏感品系没有统计差异。CYP6G4在两个敏感品系中的表达量没有统计意义的差异,但在两个抗性品系中CYP6G4的表达量比敏感品系高很多,其中JN品系CYP6G4的表达量是敏感品系的35倍左右,而在高抗性的JN-Sel品系中的表达量是敏感品系的70倍。

图2 八个P450基因在不同家蝇品系中的相对表达量
3 讨论
拟除虫菊酯杀虫剂是家蝇化学控制的主要药剂,在杀虫剂的选择压下家蝇在理论上会产生抗药性。敏感性测定结果显示山东济南的野生种群已表现出较高水平的氯菊酯抗性,在进一步接触杀虫剂的情况下,抗性继续上升,达到高抗性水平(表2)。
昆虫抗药性的机制可能因药剂类型、昆虫种类或同种昆虫不同种群的不同而不同。家蝇对拟除虫菊酯的抗性的通常机制是细胞色素P450的过量表达和/或钠离子通道基因突变。尽管钠离子通道1014位点的突变在我国抗性家蝇种群中普遍存在,但也有例外(Wangetal., 2012)。通过对济南(48头)和济南_Sel 品系(63头)家蝇的钠离子通道的基因型检测,未检测到经典的L1014F或L1014H 抗性等位基因的存在(图1),表明钠离子通道1014位点突变不是JN和JN-Sel品系对氯菊酯抗性的原因。
通过对4个抗性水平不同的家蝇品系(WHO, TJS,JN和JN-Sel)中8个P450基因的mRNA水平的定量分析发现,不同P450基因在不同品系中的表达量存在差异(图2),其中CYP6G4的mRNA水平在抗性品系中显著高于敏感品系的表达量,且抗性水平越高的品系,CYP6G4的表达量也更高。在检测的8个基因中,只有CYP6G4的表达量与氯菊酯抗性正相关,加上该基因在家蝇中的表达丰度相对较高,我们推测CYP6G4过量表达是JN和JN-Sel品系家蝇抗药性的主要机制。
CYP6G4在我国的抗性家蝇品系或种群中普遍超量表达,提示CYP6G4很可能在我国家蝇抗性进化中发挥着重要作用(Gaoetal., 2012)。家蝇CYP6G4与果蝇的Cyp6g1遗传进化距离最近,其氨基酸相似性达到了59.70%,由此推测 CYP6G4是果蝇Cyp6g1的直系同源基因。果蝇Cyp6g1过量表达已被证明可以导致果蝇对DDT和烟碱类杀虫剂的抗性(Dabornetal., 2001;2002;Joußenetal., 2008)。CYP6G4在新烟碱类杀虫剂抗性品系中比敏感品系高(Højlandetal., 2014;Højlandetal., 2017),被认为是家蝇对新烟碱类杀虫剂主要抗性基因(Højlandetal., 2014)。前人的相关研究发现以及本研究的研究结果提示CYP6G4过量表达可能导致对多个类型杀虫剂(如拟除虫菊酯类、新烟碱类)的抗药性(即交互抗药性),值得研究人员和家蝇防治工作者的重视。
猜你喜欢
江西农业学报(2022年8期)2022-11-04 07:37:02
乡村科技(2022年2期)2022-03-25 14:56:16
仪器仪表用户(2021年10期)2021-11-27 08:26:14
奇闻怪事(2020年7期)2020-09-10 01:41:40
山东第一医科大学(山东省医学科学院)学报(2019年11期)2019-11-07 02:01:12
科学种养(2016年4期)2016-04-19 03:47:43
中华老年多器官疾病杂志(2016年2期)2016-01-16 03:15:46
电源技术(2015年2期)2015-08-22 11:28:30
物理化学学报(2015年5期)2015-02-28 17:34:57
湖北科技学院学报(医学版)(2014年3期)2014-02-28 19:43:05
